
Mechanistic Understanding of Idiosyncratic Drug-Induced Hepatotoxicity Using Co-Cultures of Hepatocytes and Macrophages


Abstract
:1. Introduction
2. Materials and Methods
2.1. Material
2.2. Cell Culture
2.2.1. Maintenance, Monoculture, and Differentiation of THP-1
2.2.2. Maintenance and Monocultures of HepG2 Cells
2.2.3. Co-Culture of HepG2 and THP-1 Macrophages
2.3. Characterization of Cell Models
2.3.1. Immunofluorescence
2.3.2. Transcriptomic Analysis
2.3.3. Functionality of Co-Cultures
2.4. Toxicity Studies
2.5. Statistical Analysis
3. Results
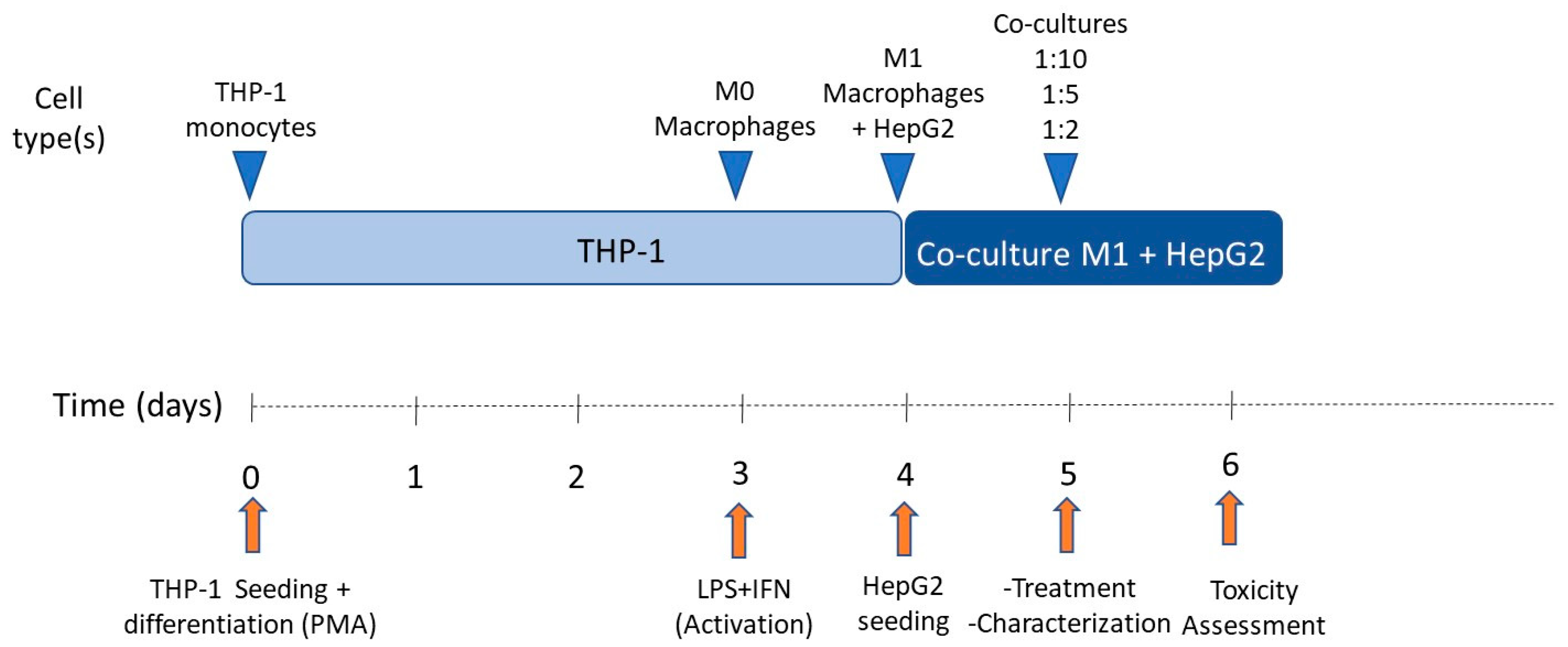
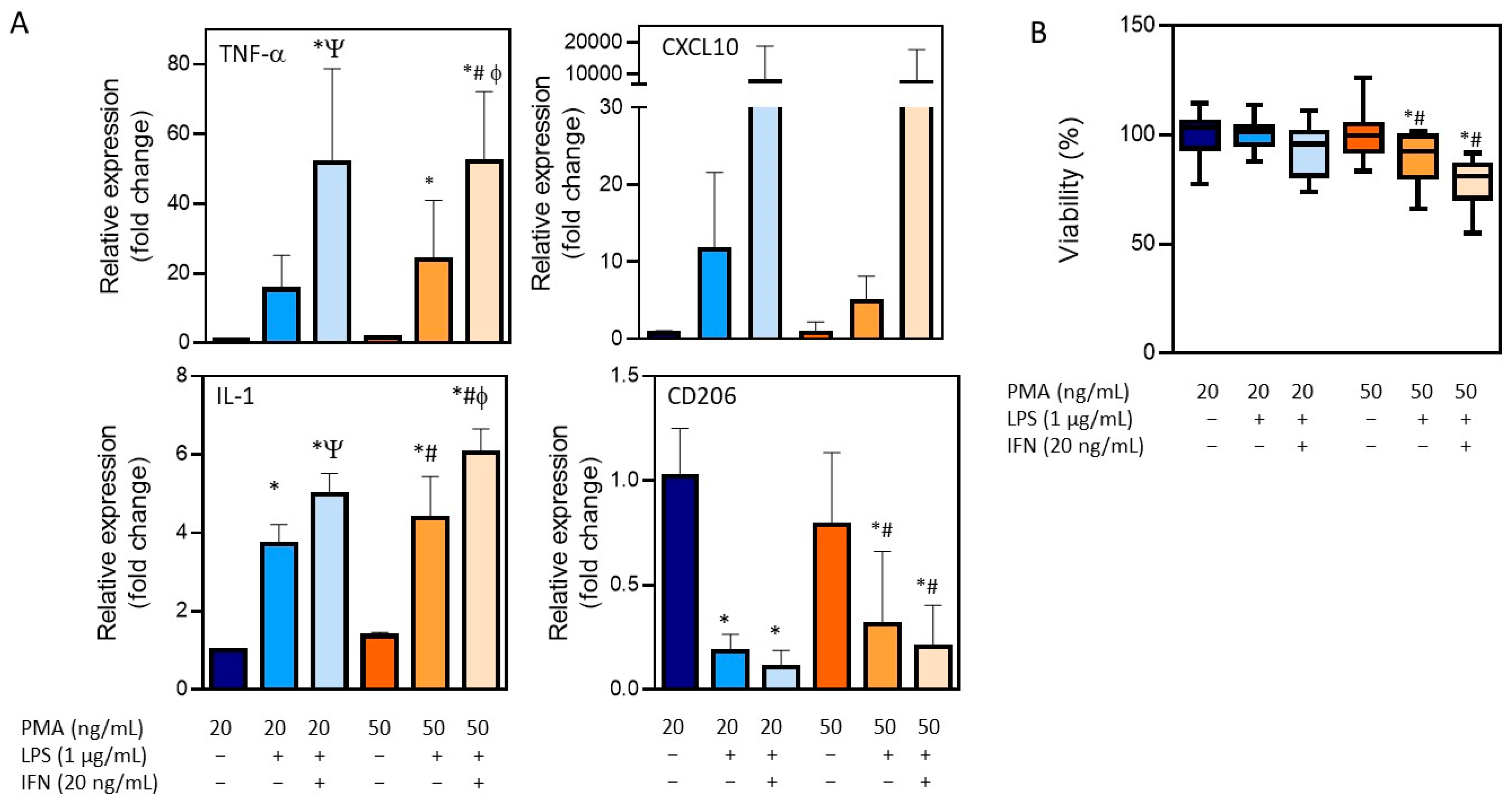
3.1. Differentiation of THP-1 Monocytes into Macrophages
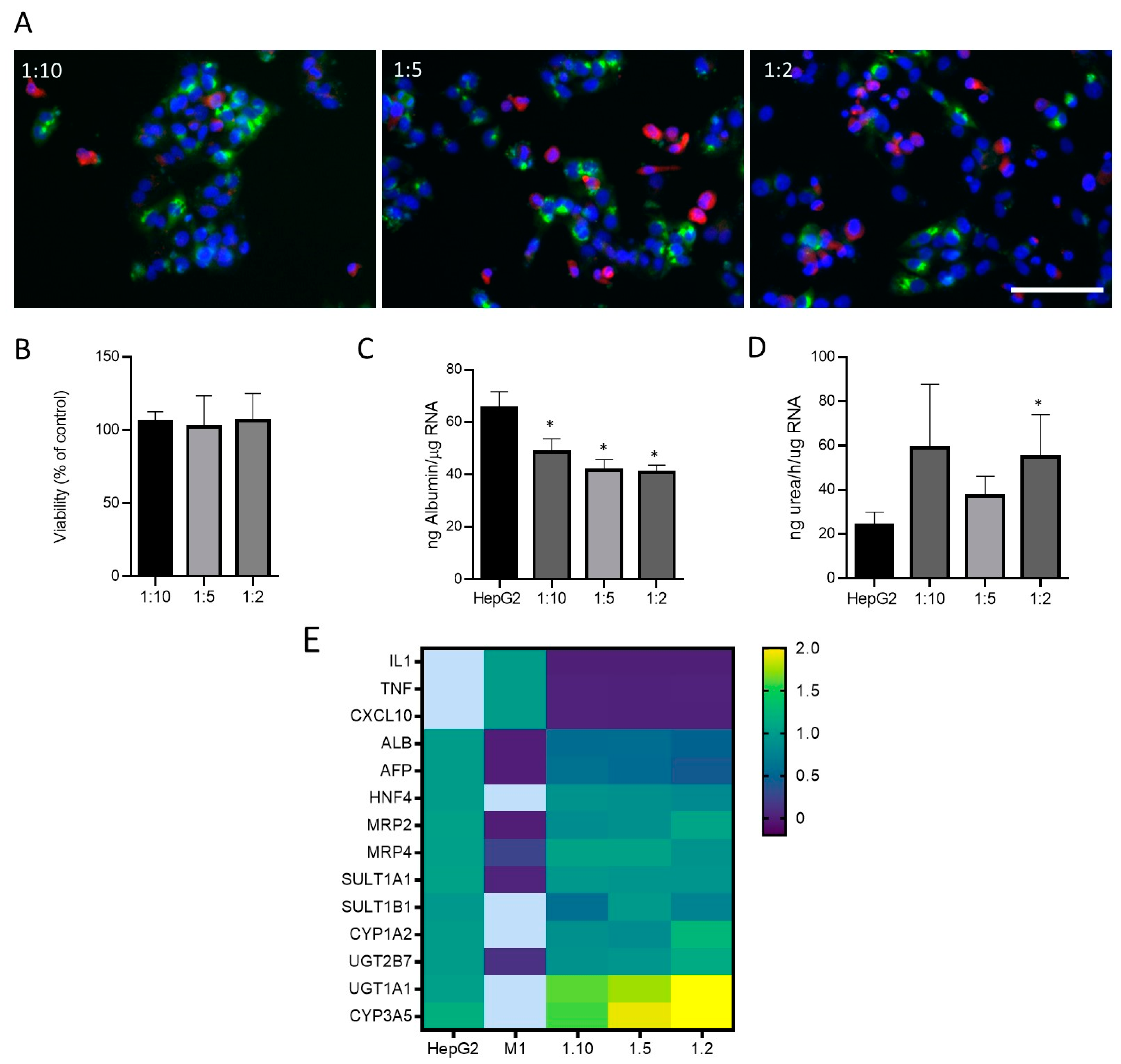
3.2. Optimization of Cell Co-Culture Conditions
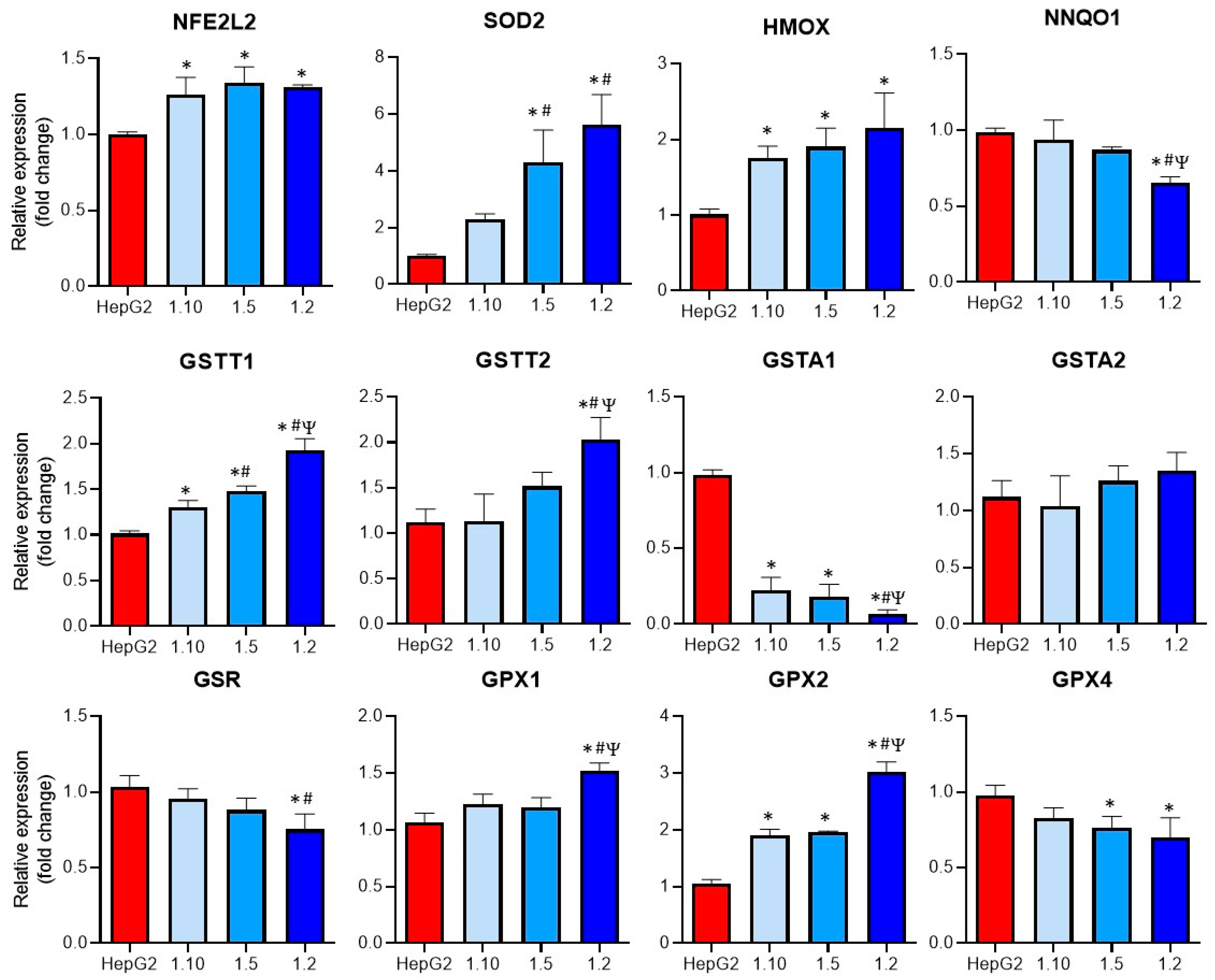
3.3. Modulation of Oxidative Stress Enzymes Expression in Co-Cultures of HepG2 and M1-THP-1 Cells
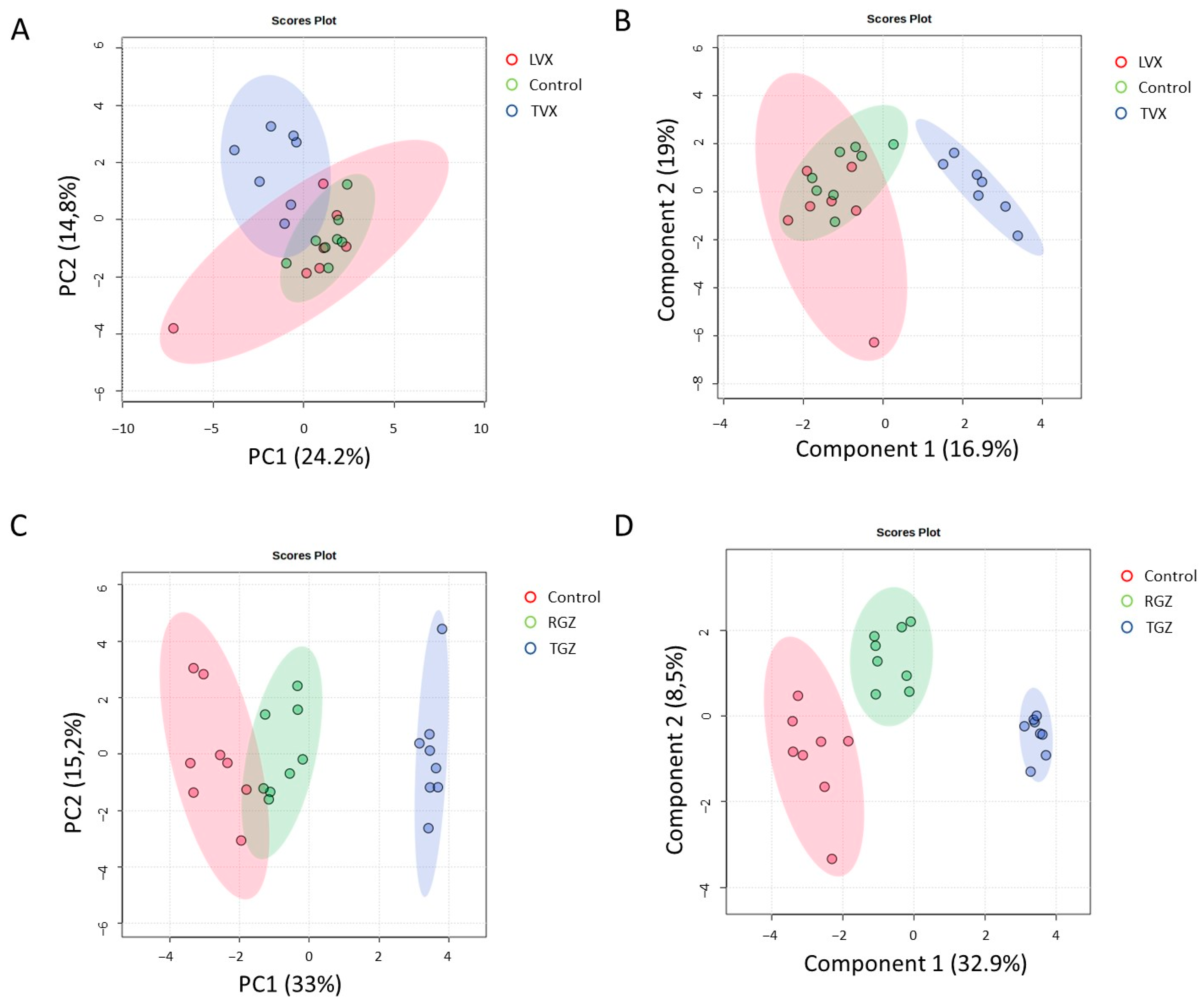
3.4. Co-Cultures Exhibit Different Sensitivity to Hepatotoxicants (Pro-Inflammatory Macrophages Increase Trovafloxacin-Induced Toxicity)
3.5. Mechanistic Understanding of Idiosyncratic Drug-Induced Toxicity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Kaplowitz, N. Idiosyncratic drug hepatotoxicity. Nat. Rev. Drug Discov. 2005, 4, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.I.; Imai, K.; Mima, K.; Nakagawa, S.; Hashimoto, D.; Chikamoto, A.; Baba, H. Idiosyncratic drug-induced liver injury: A short review. Hepatol. Commun. 2017, 1, 494–500. [Google Scholar] [CrossRef]
- Bjornsson, E.S.; Bergmann, O.M.; Bjornsson, H.K.; Kvaran, R.B.; Olafsson, S. Incidence, presentation, and outcomes in patients with drug-induced liver injury in the general population of Iceland. Gastroenterology 2013, 144, 1419–1425.e3. [Google Scholar] [CrossRef]
- Ware, B.R.; Khetani, S.R. Engineered Liver Platforms for Different Phases of Drug Development. Trends Biotechnol. 2017, 35, 172–183. [Google Scholar] [CrossRef] [Green Version]
- Dixon, L.J.; Barnes, M.; Tang, H.; Pritchard, M.T.; Nagy, L.E. Kupffer cells in the liver. Compr. Physiol. 2013, 3, 785–797. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Chang, N.; Li, L. Heterogeneity and Function of Kupffer Cells in Liver Injury. Front. Immunol. 2022, 13, 940867. [Google Scholar] [CrossRef]
- Roth, A.D.; Lee, M.Y. Idiosyncratic Drug-Induced Liver Injury (IDILI): Potential Mechanisms and Predictive Assays. BioMed Res. Int. 2017, 2017, 9176937. [Google Scholar] [CrossRef] [Green Version]
- Iorga, A.; Dara, L.; Kaplowitz, N. Drug-Induced Liver Injury: Cascade of Events Leading to Cell Death, Apoptosis or Necrosis. Int. J. Mol. Sci. 2017, 18, 1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosgrove, B.D.; Alexopoulos, L.G.; Hang, T.C.; Hendriks, B.S.; Sorger, P.K.; Griffith, L.G.; Lauffenburger, D.A. Cytokine-associated drug toxicity in human hepatocytes is associated with signaling network dysregulation. Mol. Biosyst. 2010, 6, 1195–1206. [Google Scholar] [CrossRef] [Green Version]
- Cosgrove, B.D.; Alexopoulos, L.G.; Saez-Rodriguez, J.; Griffith, L.G.; Lauffenburger, D.A. A multipathway phosphoproteomic signaling network model of idiosyncratic drug- and inflammatory cytokine-induced toxicity in human hepatocytes. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2009, 2009, 5452–5455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosgrove, B.D.; King, B.M.; Hasan, M.A.; Alexopoulos, L.G.; Farazi, P.A.; Hendriks, B.S.; Griffith, L.G.; Sorger, P.K.; Tidor, B.; Xu, J.J.; et al. Synergistic drug-cytokine induction of hepatocellular death as an in vitro approach for the study of inflammation-associated idiosyncratic drug hepatotoxicity. Toxicol. Appl. Pharmacol. 2009, 237, 317–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granitzny, A.; Knebel, J.; Muller, M.; Braun, A.; Steinberg, P.; Dasenbrock, C.; Hansen, T. Evaluation of a human in vitro hepatocyte-NPC co-culture model for the prediction of idiosyncratic drug-induced liver injury: A pilot study. Toxicol. Rep. 2017, 4, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Kostadinova, R.; Boess, F.; Applegate, D.; Suter, L.; Weiser, T.; Singer, T.; Naughton, B.; Roth, A. A long-term three dimensional liver co-culture system for improved prediction of clinically relevant drug-induced hepatotoxicity. Toxicol. Appl. Pharmacol. 2013, 268, 1–16. [Google Scholar] [CrossRef]
- Perez, R.A.; Jung, C.R.; Kim, H.W. Biomaterials and Culture Technologies for Regenerative Therapy of Liver Tissue. Adv. Healthc. Mater. 2017, 6, 1600791. [Google Scholar] [CrossRef]
- Nightingale, S.L. From the Food and Drug Administration. JAMA 1999, 282, 19. [Google Scholar] [CrossRef]
- Okuda, T.; Norioka, M.; Shitara, Y.; Horie, T. Multiple mechanisms underlying troglitazone-induced mitochondrial permeability transition. Toxicol. Appl. Pharmacol. 2010, 248, 242–248. [Google Scholar] [CrossRef]
- Moya, M.; Benet, M.; Guzman, C.; Tolosa, L.; Garcia-Monzon, C.; Pareja, E.; Castell, J.V.; Jover, R. Foxa1 reduces lipid accumulation in human hepatocytes and is down-regulated in nonalcoholic fatty liver. PLoS ONE 2012, 7, e30014. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.J.; Henstock, P.V.; Dunn, M.C.; Smith, A.R.; Chabot, J.R.; de Graaf, D. Cellular imaging predictions of clinical drug-induced liver injury. Toxicol. Sci. 2008, 105, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Tolosa, L.; Donato, M.T.; Gomez-Lechon, M.J. General Cytotoxicity Assessment by Means of the MTT Assay. Methods Mol. Biol. 2015, 1250, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for Comprehensive Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2016, 55, 14.10.11–14.10.91. [Google Scholar] [CrossRef] [PubMed]
- Baxter, E.W.; Graham, A.E.; Re, N.A.; Carr, I.M.; Robinson, J.I.; Mackie, S.L.; Morgan, A.W. Standardized protocols for differentiation of THP-1 cells to macrophages with distinct M(IFNgamma+LPS), M(IL-4) and M(IL-10) phenotypes. J. Immunol. Methods 2020, 478, 112721. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, H.; Rippe, R.; Niemela, O.; Lin, M. Roles of oxidative stress in activation of Kupffer and Ito cells in liver fibrogenesis. J. Gastroenterol. Hepatol. 1995, 10 (Suppl. 1), S50–S53. [Google Scholar] [CrossRef]
- Bleriot, C.; Barreby, E.; Dunsmore, G.; Ballaire, R.; Chakarov, S.; Ficht, X.; De Simone, G.; Andreata, F.; Fumagalli, V.; Guo, W.; et al. A subset of Kupffer cells regulates metabolism through the expression of CD36. Immunity 2021, 54, 2101–2116.e6. [Google Scholar] [CrossRef] [PubMed]
- Brenner, C.; Galluzzi, L.; Kepp, O.; Kroemer, G. Decoding cell death signals in liver inflammation. J. Hepatol. 2013, 59, 583–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segovia-Zafra, A.; Di Zeo-Sanchez, D.E.; Lopez-Gomez, C.; Perez-Valdes, Z.; Garcia-Fuentes, E.; Andrade, R.J.; Lucena, M.I.; Villanueva-Paz, M. Preclinical models of idiosyncratic drug-induced liver injury (iDILI): Moving towards prediction. Acta Pharm. Sin. B 2021, 11, 3685–3726. [Google Scholar] [CrossRef]
- Messner, S.; Agarkova, I.; Moritz, W.; Kelm, J.M. Multi-cell type human liver microtissues for hepatotoxicity testing. Arch. Toxicol. 2013, 87, 209–213. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Park, T.; Park, D.; Kang, M.G. Trovafloxacin drives inflammation-associated drug-induced adverse hepatic reaction by changing macrophage polarization. Toxicol. Vitr. 2022, 82, 105374. [Google Scholar] [CrossRef]
- Oda, S.; Matsuo, K.; Nakajima, A.; Yokoi, T. A novel cell-based assay for the evaluation of immune- and inflammatory-related gene expression as biomarkers for the risk assessment of drug-induced liver injury. Toxicol. Lett. 2016, 241, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Takebe, T.; Sekine, K.; Enomura, M.; Koike, H.; Kimura, M.; Ogaeri, T.; Zhang, R.R.; Ueno, Y.; Zheng, Y.W.; Koike, N.; et al. Vascularized and functional human liver from an iPSC-derived organ bud transplant. Nature 2013, 499, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, P.; Matchett, K.P.; Dobie, R.; Wilson-Kanamori, J.R.; Henderson, N.C. Single-cell technologies in hepatology: New insights into liver biology and disease pathogenesis. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Mathijs, K.; Timmermans, L.; Claessen, S.M.; Hecka, A.; Weusten, J.; Peters, R.; van Delft, J.H.; Kleinjans, J.C.S.; Jennen, D.G.J.; et al. Omics-based identification of the combined effects of idiosyncratic drugs and inflammatory cytokines on the development of drug-induced liver injury. Toxicol. Appl. Pharmacol. 2017, 332, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Maiuri, A.R.; Breier, A.B.; Gora, L.F.; Parkins, R.V.; Ganey, P.E.; Roth, R.A. Cytotoxic Synergy between Cytokines and NSAIDs Associated with Idiosyncratic Hepatotoxicity Is Driven by Mitogen-Activated Protein Kinases. Toxicol. Sci. 2015, 146, 265–280. [Google Scholar] [CrossRef] [Green Version]
- Pognan, F.; Beilmann, M.; Boonen, H.C.M.; Czich, A.; Dear, G.; Hewitt, P.; Mow, T.; Oinonen, T.; Roth, A.; Steger-Hartmann, T.; et al. The evolving role of investigative toxicology in the pharmaceutical industry. Nat. Rev. Drug Discov. 2023, 22, 317–335. [Google Scholar] [CrossRef]
- Pereira, C.V.; Nadanaciva, S.; Oliveira, P.J.; Will, Y. The contribution of oxidative stress to drug-induced organ toxicity and its detection in vitro and in vivo. Expert Opin. Drug Metab. Toxicol. 2012, 8, 219–237. [Google Scholar] [CrossRef]
- Villanueva-Paz, M.; Moran, L.; Lopez-Alcantara, N.; Freixo, C.; Andrade, R.J.; Lucena, M.I.; Cubero, F.J. Oxidative Stress in Drug-Induced Liver Injury (DILI): From Mechanisms to Biomarkers for Use in Clinical Practice. Antioxidants 2021, 10, 390. [Google Scholar] [CrossRef]
- Gerussi, A.; Natalini, A.; Antonangeli, F.; Mancuso, C.; Agostinetto, E.; Barisani, D.; Di Rosa, F.; Andrade, R.; Invernizzi, P. Immune-Mediated Drug-Induced Liver Injury: Immunogenetics and Experimental Models. Int. J. Mol. Sci. 2021, 22, 4557. [Google Scholar] [CrossRef]
- Kato, R.; Uetrecht, J. Supernatant from Hepatocyte Cultures with Drugs That Cause Idiosyncratic Liver Injury Activates Macrophage Inflammasomes. Chem. Res. Toxicol. 2017, 30, 1327–1332. [Google Scholar] [CrossRef] [PubMed]
- Andrade, R.J.; Tulkens, P.M. Hepatic safety of antibiotics used in primary care. J. Antimicrob. Chemother. 2011, 66, 1431–1446. [Google Scholar] [CrossRef] [Green Version]
- Shaw, P.J.; Hopfensperger, M.J.; Ganey, P.E.; Roth, R.A. Lipopolysaccharide and trovafloxacin coexposure in mice causes idiosyncrasy-like liver injury dependent on tumor necrosis factor-alpha. Toxicol. Sci. 2007, 100, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Waring, J.F.; Liguori, M.J.; Luyendyk, J.P.; Maddox, J.F.; Ganey, P.E.; Stachlewitz, R.F.; North, C.; Blomme, E.A.; Roth, R.A. Microarray analysis of lipopolysaccharide potentiation of trovafloxacin-induced liver injury in rats suggests a role for proinflammatory chemokines and neutrophils. J. Pharmacol. Exp. Ther. 2006, 316, 1080–1087. [Google Scholar] [CrossRef] [PubMed]
- Beggs, K.M.; Maiuri, A.R.; Fullerton, A.M.; Poulsen, K.L.; Breier, A.B.; Ganey, P.E.; Roth, R.A. Trovafloxacin-induced replication stress sensitizes HepG2 cells to tumor necrosis factor-alpha-induced cytotoxicity mediated by extracellular signal-regulated kinase and ataxia telangiectasia and Rad3-related. Toxicology 2015, 331, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Liguori, M.J.; Anderson, M.G.; Bukofzer, S.; McKim, J.; Pregenzer, J.F.; Retief, J.; Spear, B.B.; Waring, J.F. Microarray analysis in human hepatocytes suggests a mechanism for hepatotoxicity induced by trovafloxacin. Hepatology 2005, 41, 177–186. [Google Scholar] [CrossRef]
- Fujiwara, R.; Sumida, K.; Kutsuno, Y.; Sakamoto, M.; Itoh, T. UDP-glucuronosyltransferase (UGT) 1A1 mainly contributes to the glucuronidation of trovafloxacin. Drug Metab. Pharmacokinet. 2015, 30, 82–88. [Google Scholar] [CrossRef]
- Mitsugi, R.; Sumida, K.; Fujie, Y.; Tukey, R.H.; Itoh, T.; Fujiwara, R. Acyl-glucuronide as a Possible Cause of Trovafloxacin-Induced Liver Toxicity: Induction of Chemokine (C-X-C Motif) Ligand 2 by Trovafloxacin Acyl-glucuronide. Biol. Pharm. Bull. 2016, 39, 1604–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, X.; Luyendyk, J.P.; Ganey, P.E.; Roth, R.A. Inflammatory stress and idiosyncratic hepatotoxicity: Hints from animal models. Pharmacol. Rev. 2009, 61, 262–282. [Google Scholar] [CrossRef] [Green Version]
- Murphy, E.J.; Davern, T.J.; Shakil, A.O.; Shick, L.; Masharani, U.; Chow, H.; Freise, C.; Lee, W.M.; Bass, N.M. Troglitazone-induced fulminant hepatic failure. Acute Liver Failure Study Group. Dig. Dis. Sci. 2000, 45, 549–553. [Google Scholar] [CrossRef]
- Bonkovsky, H.L.; Azar, R.; Bird, S.; Szabo, G.; Banner, B. Severe cholestatic hepatitis caused by thiazolidinediones: Risks associated with substituting rosiglitazone for troglitazone. Dig. Dis. Sci. 2002, 47, 1632–1637. [Google Scholar] [CrossRef]
- Yokoi, T. Troglitazone. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 419–435. [Google Scholar]
- Shen, C.; Meng, Q.; Zhang, G. Species-specific toxicity of troglitazone on rats and human by gel entrapped hepatocytes. Toxicol. Appl. Pharmacol. 2012, 258, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Ong, M.M.; Latchoumycandane, C.; Boelsterli, U.A. Troglitazone-induced hepatic necrosis in an animal model of silent genetic mitochondrial abnormalities. Toxicol. Sci. 2007, 97, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, P.K.; Hart, T.; Elcock, F.; Zhang, C.; Hahn, L.; McFarland, D.; Schwartz, L.; Morgan, D.G.; Bugelski, P. Troglitazone-induced intracellular oxidative stress in rat hepatoma cells: A flow cytometric assessment. Cytom. A 2003, 52, 28–35. [Google Scholar] [CrossRef]
- Pu, S.; Pan, Y.; Zhang, Q.; You, T.; Yue, T.; Zhang, Y.; Wang, M. Endoplasmic Reticulum Stress and Mitochondrial Stress in Drug-Induced Liver Injury. Molecules 2023, 28, 3160. [Google Scholar] [CrossRef] [PubMed]
- Edling, Y.; Sivertsson, L.K.; Butura, A.; Ingelman-Sundberg, M.; Ek, M. Increased sensitivity for troglitazone-induced cytotoxicity using a human in vitro co-culture model. Toxicol. Vitr. 2009, 23, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Tasnim, F.; Huang, X.; Lee, C.Z.W.; Ginhoux, F.; Yu, H. Recent Advances in Models of Immune-Mediated Drug-Induced Liver Injury. Front. Toxicol. 2021, 3, 605392. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | CAS Number | Supplier | Concentrations (µM) | Cmax (µM) [18] | iDILI Classification |
|---|---|---|---|---|---|
| Trovafloxacin | 147059-72-1 | Merck | 25–800 | 4.1 | + |
| Levofloxacin | 100986-85-4 | Merck | 25–1000 | 15.7 | − |
| Troglitazone | 97322-87-7 | Merck | 50–250 | 6.4 | + |
| Rosiglitazone | 122320-73-4 | Merck | 50–250 | 1 | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villanueva-Badenas, E.; Donato, M.T.; Tolosa, L. Mechanistic Understanding of Idiosyncratic Drug-Induced Hepatotoxicity Using Co-Cultures of Hepatocytes and Macrophages. Antioxidants 2023, 12, 1315. https://doi.org/10.3390/antiox12071315
Villanueva-Badenas E, Donato MT, Tolosa L. Mechanistic Understanding of Idiosyncratic Drug-Induced Hepatotoxicity Using Co-Cultures of Hepatocytes and Macrophages. Antioxidants. 2023; 12(7):1315. https://doi.org/10.3390/antiox12071315
Chicago/Turabian StyleVillanueva-Badenas, Estela, M. Teresa Donato, and Laia Tolosa. 2023. "Mechanistic Understanding of Idiosyncratic Drug-Induced Hepatotoxicity Using Co-Cultures of Hepatocytes and Macrophages" Antioxidants 12, no. 7: 1315. https://doi.org/10.3390/antiox12071315
APA StyleVillanueva-Badenas, E., Donato, M. T., & Tolosa, L. (2023). Mechanistic Understanding of Idiosyncratic Drug-Induced Hepatotoxicity Using Co-Cultures of Hepatocytes and Macrophages. Antioxidants, 12(7), 1315. https://doi.org/10.3390/antiox12071315