
Sedum middendorffianum Maxim Induces Apoptosis and Inhibits the Invasion of Human Ovarian Cancer Cells via Oxidative Stress Regulation

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample
2.2. Materials
2.3. Cell Culture and Cell Viability
2.4. Annexin V and PI Double Staining for Apoptosis Analysis
2.5. Western Blot Analysis
2.6. Measurement of Reactive Oxygen Species (ROS)
2.7. Invasion Assay
2.8. High Performance Liquid Chromatography-Mass Spectrometry Analysis
2.9. Statistical Analysis
3. Results
3.1. SMM Extract Inhibits the Growth of Human Ovarian Cancer Cells
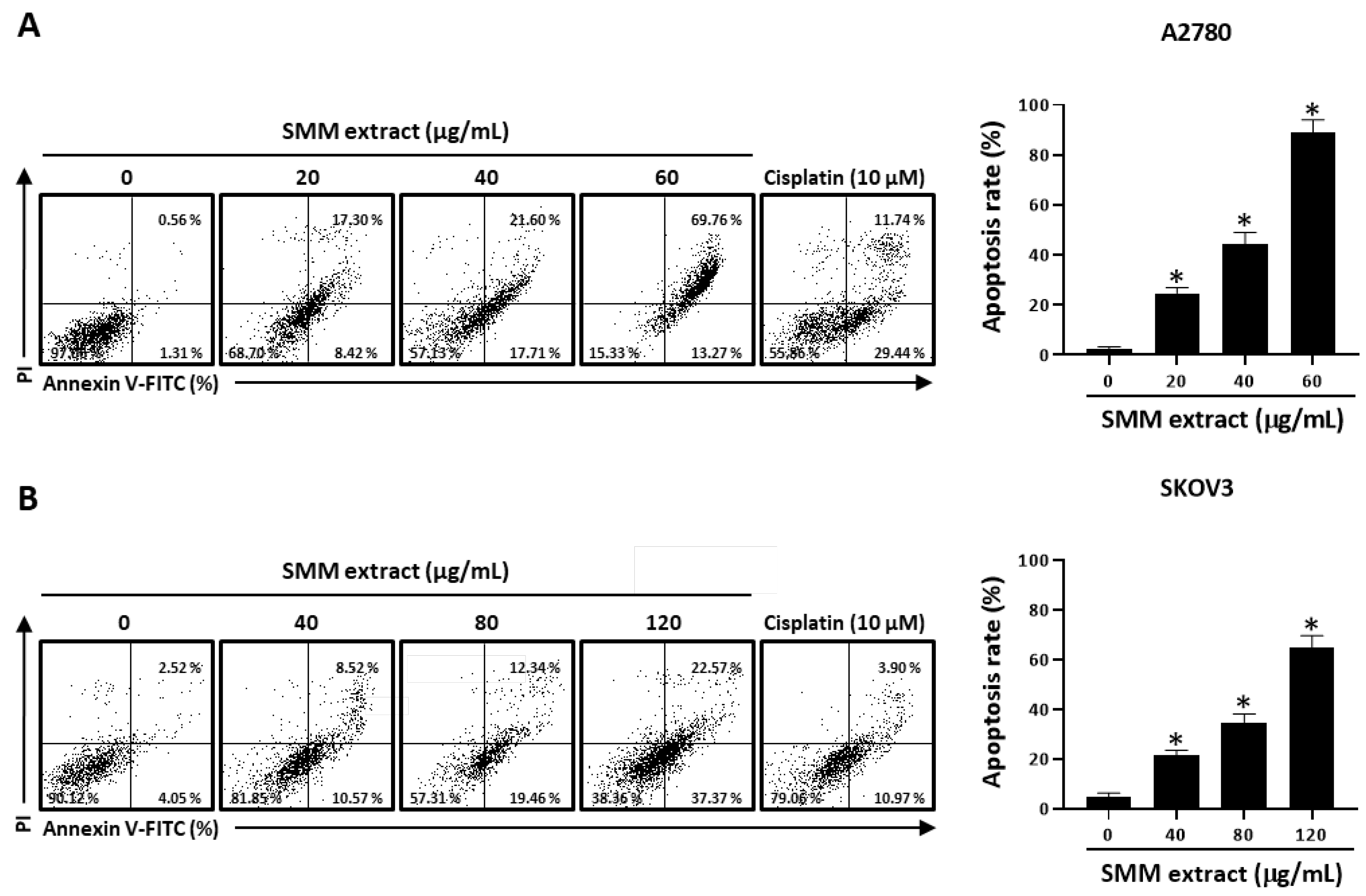
3.2. SMM Extract Induces Caspase-Dependent Apoptotic Cell Death in Human Ovarian Cancer Cells
3.3. SMM Extract Increases the Intracellular Levels of Reactive Oxygen Species (ROS), which are Associated with SMM-Induced Apoptotic Cell Death in Ovarian Cancer Cells
3.4. SMM Extract Inhibits Cell Invasion and MMP Expression in Human Ovarian Cancer Cells
3.5. Anti-Invasive Activity of SMM Extract is Associated with Intracellular ROS Levels
3.6. HPLC-MS Analysis of SMM Extract
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Das, P.M.; Bast, R.C., Jr. Early detection of ovarian cancer. Biomark. Med. 2008, 2, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Yeung, T.L.; Leung, C.S.; Yip, K.P.; Au Yeung, C.L.; Wong, S.T.; Mok, S.C. Cellular and molecular processes in ovarian cancer metastasis. A Review in the Theme: Cell and Molecular Processes in Cancer Metastasis. Am. J. Physiol. Cell Physiol. 2015, 309, C444–C456. [Google Scholar] [CrossRef] [Green Version]
- Pokhriyal, R.; Hariprasad, R.; Kumar, L.; Hariprasad, G. Chemotherapy Resistance in Advanced Ovarian Cancer Patients. Biomark. Cancer 2019, 11, 1179299X19860815. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the last 25 years. J. Nat. Prod. 2007, 70, 461–477. [Google Scholar] [CrossRef] [Green Version]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Volenzo, T.; Odiyo, J. Integrating endemic medicinal plants into the global value chains: The ecological degradation challenges and opportunities. Heliyon 2020, 6, e04970. [Google Scholar] [CrossRef]
- Avrelija, C.; Walter, C. Antimicrobial agents deriving from indigenous plants. Recent Pat. Food Nutr. Agric. 2010, 2, 83–92. [Google Scholar] [CrossRef]
- Koo, K.A.; Kong, W.S.; Nibbelink, N.P.; Hopkinson, C.S.; Lee, J.H. Potential Effects of Climate Change on the Distribution of Cold-Tolerant Evergreen Broadleaved Woody Plants in the Korean Peninsula. PLoS ONE 2015, 10, e0134043. [Google Scholar] [CrossRef]
- Jeong, B.R.; Sivanesan, I. Micropropagation, berberine content and antitumor activity of Jeffersonia dubia (Maxim.) Benth et Hook. Plant Cell Tissue Org. 2016, 124, 453–458. [Google Scholar] [CrossRef]
- Kang, J.E.; Jeon, B.J.; Park, M.Y.; Kim, B.S. Inhibitory Activity of Sedum middendorffianum-Derived 4-Hydroxybenzoic Acid and Vanillic Acid on the Type III Secretion System of Pseudomonas syringae pv. tomato DC3000. Plant Pathol. J. 2020, 36, 608–617. [Google Scholar] [CrossRef]
- Kuete, V.; Seo, E.J.; Krusche, B.; Oswald, M.; Wiench, B.; Schroder, S.; Greten, H.J.; Lee, I.S.; Efferth, T. Cytotoxicity and pharmacogenomics of medicinal plants from traditional korean medicine. Evid. Based Complement. Alternat. Med. 2013, 2013, 341724. [Google Scholar] [CrossRef]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase functions in cell death and disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef] [Green Version]
- Hensley, P.; Mishra, M.; Kyprianou, N. Targeting caspases in cancer therapeutics. Biol. Chem. 2013, 394, 831–843. [Google Scholar] [CrossRef] [Green Version]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Zong, X.; Nephew, K.P. Ovarian Cancer Stem Cells: Role in Metastasis and Opportunity for Therapeutic Targeting. Cancers 2019, 11, 934. [Google Scholar] [CrossRef] [Green Version]
- Lengyel, E. Ovarian cancer development and metastasis. Am. J. Pathol. 2010, 177, 1053–1064. [Google Scholar] [CrossRef]
- Al-Alem, L.; Curry, T.E., Jr. Ovarian cancer: Involvement of the matrix metalloproteinases. Reproduction 2015, 150, R55–R64. [Google Scholar] [CrossRef] [Green Version]
- Peiris-Pages, M.; Martinez-Outschoorn, U.E.; Sotgia, F.; Lisanti, M.P. Metastasis and Oxidative Stress: Are Antioxidants a Metabolic Driver of Progression? Cell Metab. 2015, 22, 956–958. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Li, W.; Chen, J.; Zhang, X.; Zhang, W.; Duan, X.; Lei, B.; Huang, R. Transcriptomics Integrated with Metabolomics Reveals 2-Methoxy-1, 4-Naphthoquinone-Based Carbon Dots Induced Molecular Shifts in Penicillium italicum. J. Fungi 2022, 8, 420. [Google Scholar] [CrossRef]
- Yoon, J.; Park, J.; Kim, B. Skin whitening effect of Sedum kamtschaticum fisch. solvent fractions. J. Korean Appl. Sci. Technol. 2020, 37, 1239–1247. [Google Scholar] [CrossRef]
- Hosseini, A.; Ghorbani, A. Cancer therapy with phytochemicals: Evidence from clinical studies. Avicenna J. Phytomedicine 2015, 5, 84–97. [Google Scholar]
- Zaid, H.; Silbermann, M.; Amash, A.; Gincel, D.; Abdel-Sattar, E.; Sarikahya, N.B. Medicinal Plants and Natural Active Compounds for Cancer Chemoprevention/Chemotherapy. Evid. Based Complement. Alternat. Med. 2017, 2017, 7952417. [Google Scholar] [CrossRef]
- Rajagopalan, U.; Samarakoon, S.R.; Tennekoon, K.H.; Malavige, N.; de Silva, E.D. Screening of five Sri Lankan endemic plants for anti-cancer effects on breast cancer stem cells isolated from MCF-7 and MDA-MB-231 cell lines. Trop. J. Pharm. Res. 2018, 17, 1825–1832. [Google Scholar] [CrossRef]
- Manukyan, A.; Lumlerdkij, N.; Heinrich, M. Caucasian endemic medicinal and nutraceutical plants: In-vitro antioxidant and cytotoxic activities and bioactive compounds. J. Pharm. Pharm. 2019, 71, 1152–1161. [Google Scholar] [CrossRef]
- Neergheen, V.S.; Bahorun, T.; Jen, L.S.; Aruoma, O.I. Bioefficacy of mauritian endemic medicinal plants: Assessment of their phenolic contents and antioxidant potential. Pharm. Biol. 2007, 45, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.-B.; Deng, T.; Dou, Q.-L.; He, L.; Lv, X.-Y.; Jiang, H.J.P. Sedum lipingense (Crassulaceae) identifying a new stonecrop species in SE Guizhou, China, based on morphological and molecular evidence. PhytoKeys 2019, 134, 125. [Google Scholar] [CrossRef]
- Qiu, Q.; Jiang, L.; Zhen, H.; Huang, F.; Zhen, D.; Ye, M.; Meng, X.; Liu, Y.; Qin, X. Promotion of HepG2 cell apoptosis by Sedum emarginatum Migo and the mechanism of action. BMC Complement. Med. Ther. 2022, 22, 31. [Google Scholar] [CrossRef]
- Huang, D.; Zhang, W.; Huang, D.; Wu, J. Antitumor activity of the aqueous extract from Sedum sarmentosum Bunge in vitro. Cancer Biother. Radiopharm. 2010, 25, 81–88. [Google Scholar] [CrossRef]
- Shin, J.A.; Won, D.H.; Swarup, N.; Ahn, M.H.; Yang, S.O.; Chawla, K.; Kim, J.; Choi, S.J.; Ahn, C.H.; Oh, K.Y.; et al. Methanol extract of Sedum oryzifolium and its constituent, trehalose, impede the invasiveness of oral squamous cell carcinoma cell lines via downregulation of Slug. Phytomedicine 2021, 91, 153670. [Google Scholar] [CrossRef]
- Shnyakina, G.P.; Zapesochnaya, G.G. Flavonols and phenolic compounds of Sedum middendorffianum. Chem. Nat. Compd. 1973, 9, 645. [Google Scholar] [CrossRef]
- Ren, M.X.; Deng, X.H.; Ai, F.; Yuan, G.Y.; Song, H.Y. Effect of quercetin on the proliferation of the human ovarian cancer cell line SKOV-3 in vitro. Exp. Ther. Med. 2015, 10, 579–583. [Google Scholar] [CrossRef] [Green Version]
- Gibellini, L.; Pinti, M.; Nasi, M.; De Biasi, S.; Roat, E.; Bertoncelli, L.; Cossarizza, A. Interfering with ROS Metabolism in Cancer Cells: The Potential Role of Quercetin. Cancers 2010, 2, 1288. [Google Scholar] [CrossRef] [Green Version]
- Teekaraman, D.; Elayapillai, S.P.; Viswanathan, M.P.; Jagadeesan, A. Quercetin inhibits human metastatic ovarian cancer cell growth and modulates components of the intrinsic apoptotic pathway in PA-1cell line. Chem. Biol. Interact. 2019, 300, 91–100. [Google Scholar] [CrossRef]
- Abourashed, E.A. Bioavailability of Plant-Derived Antioxidants. Antioxidants 2013, 2, 309. [Google Scholar] [CrossRef] [Green Version]
- Thilakarathna, S.H.; Rupasinghe, H.P. Flavonoid bioavailability and attempts for bioavailability enhancement. Nutrients 2013, 5, 3367. [Google Scholar] [CrossRef] [Green Version]
- Gerl, R.; Vaux, D.L. Apoptosis in the development and treatment of cancer. Carcinogenesis 2005, 26, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Yuan, J. Caspases in apoptosis and beyond. Oncogene 2008, 27, 6194–6206. [Google Scholar] [CrossRef] [Green Version]
- Boice, A.; Bouchier-Hayes, L. Targeting apoptotic caspases in cancer. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118688. [Google Scholar] [CrossRef]
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L1005–L1028. [Google Scholar] [CrossRef] [Green Version]
- Bae, Y.S.; Oh, H.; Rhee, S.G.; Yoo, Y.D. Regulation of reactive oxygen species generation in cell signaling. Mol. Cells 2011, 32, 491–509. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, M.A.; Sethi, G. Role of Reactive Oxygen Species in Cancer Progression: Molecular Mechanisms and Recent Advancements. Biomolecules 2019, 9, 735. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Kim, H.S.; Seo, Y.R. Understanding of ROS-Inducing Strategy in Anticancer Therapy. Oxid. Med. Cell. Longev. 2019, 2019, 5381692. [Google Scholar] [CrossRef]
- Xi, X.; Wang, J.; Qin, Y.; You, Y.; Huang, W.; Zhan, J. The Biphasic Effect of Flavonoids on Oxidative Stress and Cell Proliferation in Breast Cancer Cells. Antioxidants 2022, 11, 622. [Google Scholar] [CrossRef]
- Kim, T.H.; Park, J.H.; Woo, J.S. Resveratrol induces cell death through ROS-dependent downregulation of Notch1/PTEN/Akt signaling in ovarian cancer cells. Mol. Med. Rep. 2019, 19, 3353–3360. [Google Scholar] [CrossRef]
- Yadav, P.; Yadav, R.; Jain, S.; Vaidya, A. Caspase-3: A primary target for natural and synthetic compounds for cancer therapy. Chem. Biol. Drug Des. 2021, 98, 144–165. [Google Scholar] [CrossRef]
- Alonso, M.M.; Asumendi, A.; Villar, J.; Gil, M.J.; Martinez-Merino, V.; Encio, I.J.; Migliaccio, M. New benzo(b)thiophenesulphonamide 1,1-dioxide derivatives induce a reactive oxygen species-mediated process of apoptosis in tumour cells. Oncogene 2003, 22, 3759–3769. [Google Scholar] [CrossRef] [Green Version]
- Denning, T.L.; Takaishi, H.; Crowe, S.E.; Boldogh, I.; Jevnikar, A.; Ernst, P.B. Oxidative stress induces the expression of Fas and Fas ligand and apoptosis in murine intestinal epithelial cells. Free Radic. Biol. Med. 2002, 33, 1641–1650. [Google Scholar] [CrossRef]
- Inoue, A.; Muranaka, S.; Fujita, H.; Kanno, T.; Tamai, H.; Utsumi, K. Molecular mechanism of diclofenac-induced apoptosis of promyelocytic leukemia: Dependency on reactive oxygen species, Akt, Bid, cytochrome and caspase pathway. Free Radic. Biol. Med. 2004, 37, 1290–1299. [Google Scholar] [CrossRef]
- Maraldi, T. Natural Compounds as Modulators of NADPH Oxidases. Oxid. Med. Cell. Longev. 2013, 2013, 271602. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.Y.; Lee, N.K.; Wang, Y.Y.; Hong, J.P.; Son, S.R.; Gu, D.H.; Jang, D.S.; Choi, J.H. 1′-Acetoxyeugenol Acetate Isolated from Thai Ginger Induces Apoptosis in Human Ovarian Cancer Cells by ROS Production via NADPH Oxidase. Antioxidants 2022, 11, 293. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.H.; Jeong, M.; Jang, D.S.; Choi, J.H. Gomisin L1, a Lignan Isolated from Schisandra Berries, Induces Apoptosis by Regulating NADPH Oxidase in Human Ovarian Cancer Cells. Life 2021, 11, 858. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Ahn, J.H.; Cho, Y.J.; Kim, H.Y.; Yang, Y.I.; Lee, K.T.; Jang, D.S.; Choi, J.H. alpha-Terthienylmethanol, isolated from Eclipta prostrata, induces apoptosis by generating reactive oxygen species via NADPH oxidase in human endometrial cancer cells. J. Ethnopharmacol. 2015, 169, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Zhang, Y.; Dusting, G.J. NADPH oxidase-mediated redox signaling: Roles in cellular stress response, stress tolerance, and tissue repair. Pharmacol. Rev. 2011, 63, 218–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintero-Fabian, S.; Arreola, R.; Becerril-Villanueva, E.; Torres-Romero, J.C.; Arana-Argaez, V.; Lara-Riegos, J.; Ramirez-Camacho, M.A.; Alvarez-Sanchez, M.E. Role of Matrix Metalloproteinases in Angiogenesis and Cancer. Front. Oncol. 2019, 9, 1370. [Google Scholar] [CrossRef] [Green Version]
- Shay, G.; Lynch, C.C.; Fingleton, B. Moving targets: Emerging roles for MMPs in cancer progression and metastasis. Matrix Biol. 2015, 44–46, 200–206. [Google Scholar] [CrossRef]
- Nishikawa, M. Reactive oxygen species in tumor metastasis. Cancer Lett. 2008, 266, 53–59. [Google Scholar] [CrossRef]
- Wu, X.; Zhu, Y.; Yan, H.; Liu, B.; Li, Y.; Zhou, Q.; Xu, K. Isothiocyanates induce oxidative stress and suppress the metastasis potential of human non-small cell lung cancer cells. BMC Cancer 2010, 10, 269. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Zhang, Y.; Luo, Y.; Xu, B.; Yao, Y.; Deng, Y.; Yang, F.; Ye, T.; Wang, G.; Cheng, Z.J.B.; et al. Hinokiflavone induces apoptosis in melanoma cells through the ROS-mitochondrial apoptotic pathway and impairs cell migration and invasion. Biomed. Pharmacother. 2018, 103, 101–110. [Google Scholar] [CrossRef]
- Liu, F.; Gomez Garcia, A.M.; Meyskens, F.L., Jr. NADPH oxidase 1 overexpression enhances invasion via matrix metalloproteinase-2 and epithelial-mesenchymal transition in melanoma cells. J. Investig. Dermatol. 2012, 132, 2033–2041. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.H.; Jin, S.W.; Kim, S.J.; Pham, T.H.; Choi, J.H.; Jeong, H.G. Tetrabromobisphenol A Induces MMP-9 Expression via NADPH Oxidase and the activation of ROS, MAPK, and Akt Pathways in Human Breast Cancer MCF-7 Cells. Toxicol. Res. 2019, 35, 93–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Name | a IC50 (μg/mL) | |
|---|---|---|---|
| A2780 | IOSE80PC | ||
| Oleaceae | Forsythia saxatilis Nakai | 165.32 ± 5.98 | 138.71 ± 5.80 |
| Oleaceae | Forsythia velutina Nakai | 133.15 ± 10.43 | >200 |
| Ranunculaceae | Ranunculus crucilobus H.Lév | 153.62 ± 4.68 | >200 |
| Berberidaceae | Jeffersonia dubia Maxim | 98.35 ± 6.51 | 89.20 ± 5.91 |
| Crassulaceae | Sedum middendorffianum Maxim | 50.25 ± 4.35 | >200 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.-Y.; Jeong, M.; Lee, K.; Kim, J.-O.; Lee, W.H.; Park, I.; Kwon, H.C.; Choi, J.-H. Sedum middendorffianum Maxim Induces Apoptosis and Inhibits the Invasion of Human Ovarian Cancer Cells via Oxidative Stress Regulation. Antioxidants 2023, 12, 1386. https://doi.org/10.3390/antiox12071386
Choi J-Y, Jeong M, Lee K, Kim J-O, Lee WH, Park I, Kwon HC, Choi J-H. Sedum middendorffianum Maxim Induces Apoptosis and Inhibits the Invasion of Human Ovarian Cancer Cells via Oxidative Stress Regulation. Antioxidants. 2023; 12(7):1386. https://doi.org/10.3390/antiox12071386
Chicago/Turabian StyleChoi, Ju-Yeon, Miran Jeong, Kijun Lee, Jin-Ok Kim, Wan Hee Lee, InWha Park, Hak Cheol Kwon, and Jung-Hye Choi. 2023. "Sedum middendorffianum Maxim Induces Apoptosis and Inhibits the Invasion of Human Ovarian Cancer Cells via Oxidative Stress Regulation" Antioxidants 12, no. 7: 1386. https://doi.org/10.3390/antiox12071386
APA StyleChoi, J. -Y., Jeong, M., Lee, K., Kim, J. -O., Lee, W. H., Park, I., Kwon, H. C., & Choi, J. -H. (2023). Sedum middendorffianum Maxim Induces Apoptosis and Inhibits the Invasion of Human Ovarian Cancer Cells via Oxidative Stress Regulation. Antioxidants, 12(7), 1386. https://doi.org/10.3390/antiox12071386