


Biological Activity and Metabolomics of Griffonia simplicifolia Seeds Extracted with Different Methodologies

 ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Preparation of Extracts
2.2.1. Soxhlet Extraction (SE)
2.2.2. Maceration (ME)
2.2.3. Microwave-Assisted Extraction (MAE)
2.3. Total Polyphenol Content (TPC)
2.4. Total Proanthocyanidin Content (TPAC)
2.5. Total Alkaloid Content (TAC)
2.6. Antioxidant Activity
2.6.1. ABTS Assay
2.6.2. DPPH Assay
2.6.3. FRAP Assay
2.7. HPLC-DAD-ESI-MS/MS
2.7.1. Alkaloids and N-Containing Compounds
2.7.2. Flavonoids
2.8. Evaluation of Antiproliferative Activity
2.8.1. Cell Culture
2.8.2. MTT Assay
2.9. Evaluation of Antibacterial Activity
2.9.1. Bacterial Strains
2.9.2. Evaluation of Antibacterial Properties
2.10. Statistical Analysis
3. Results and Discussion
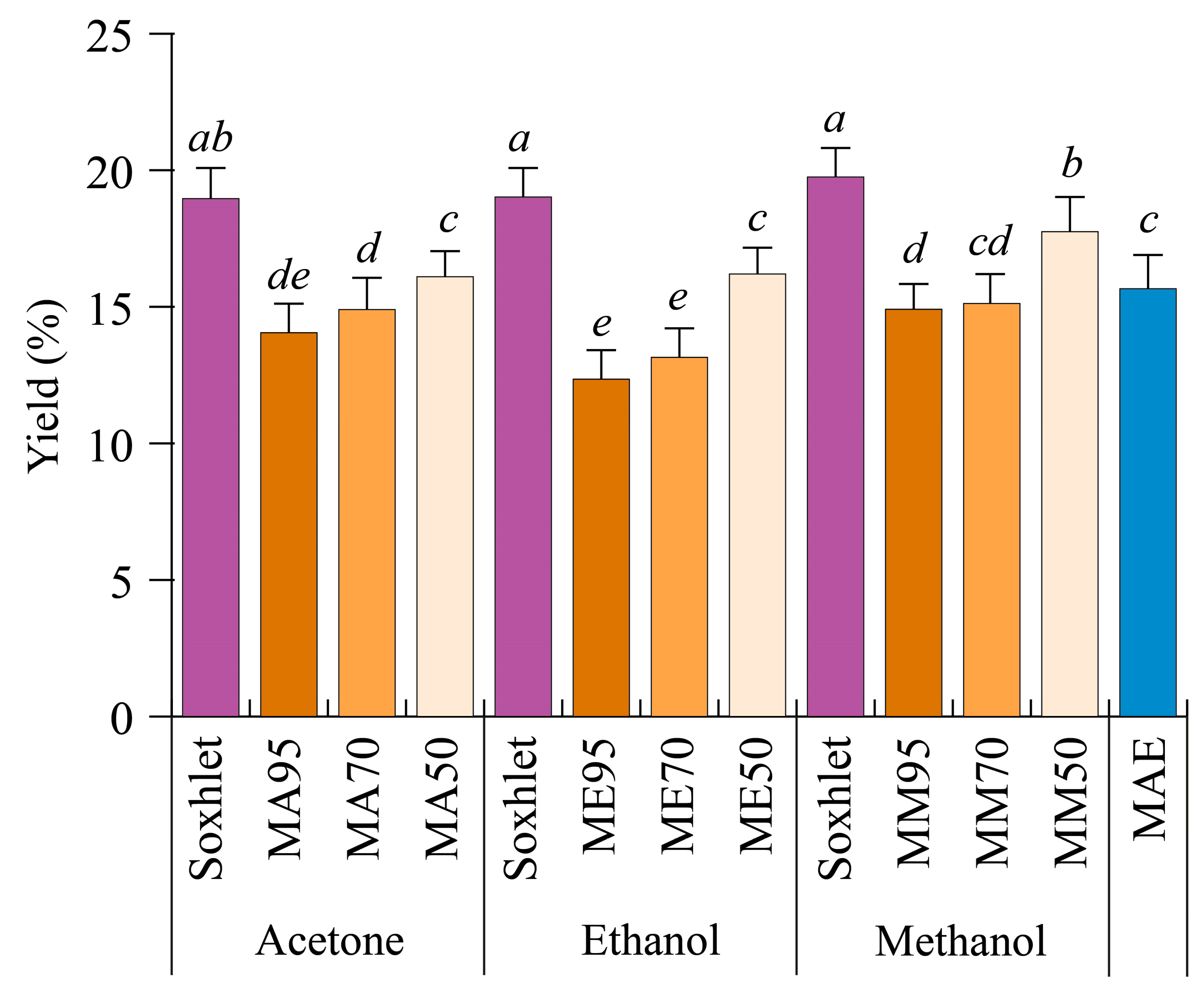
3.1. Extraction Yield of G. simplicifolia Seeds Depends on Solvent Polarity and Water Content
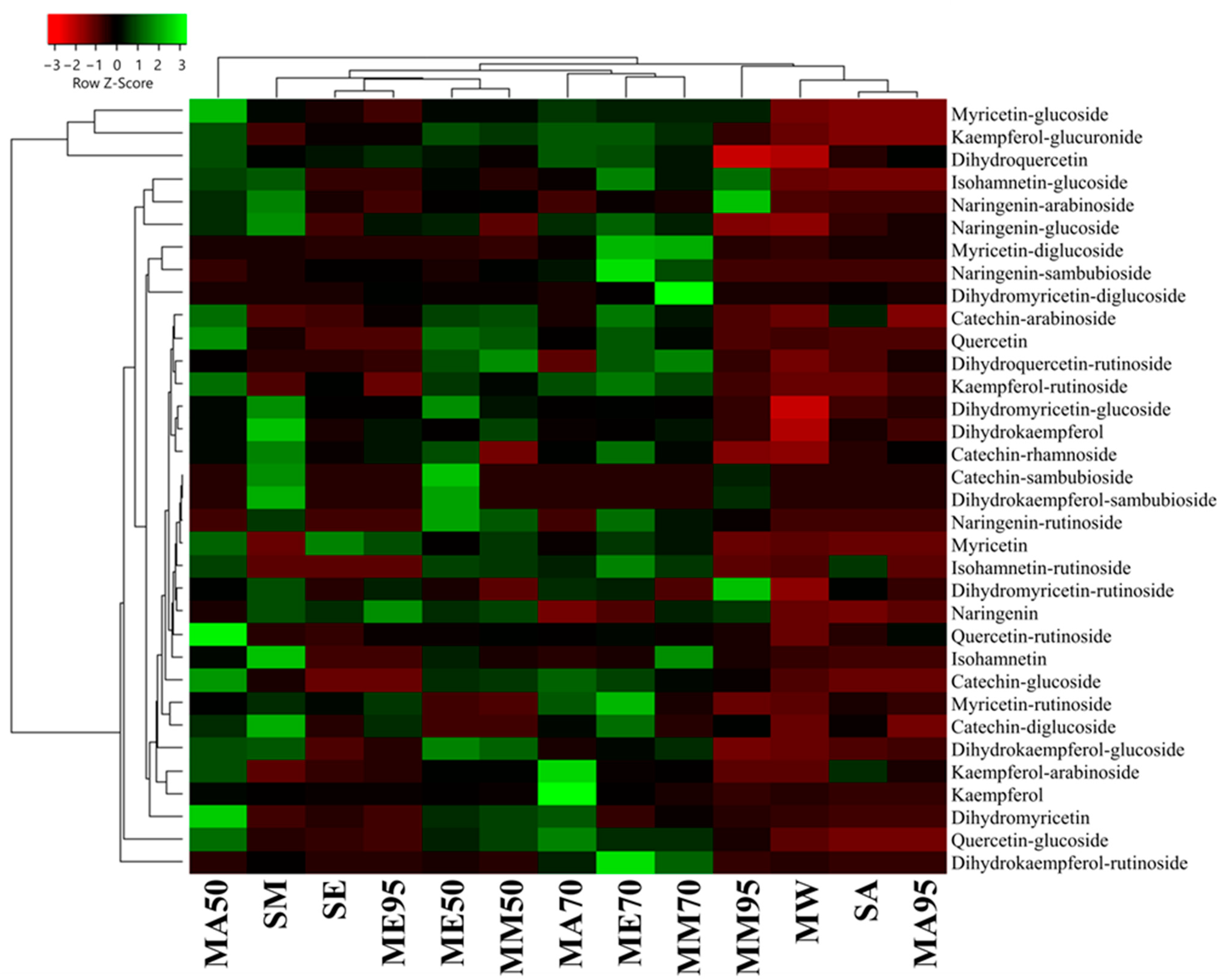
3.2. Acetone and Ethanol Are the Best Solvents for Flavonoids and Flavan-3-ols, whereas MAE Is More Efficient for N-Containing Compounds
3.3. Isomyricitrin, Taxifolin and a Flavonol Glucuronide Are the Main Polyphenols of G. simplicifolia
3.4. 5-HTP Is the Most Important N-Containing Compoud of G. simplicifolia and Is Best Extracted by Organic Solvent Maceration in the Presence of Water
3.5. Hydroalcoholic Extracts Show the Highest Radical Scavenging and Metal-Reducing Antioxidant Power
3.6. G. simplicifolia Extracts Exhibit a Concentration-Dependent Antiproliferative Activity, with a Significant Variability across the Different Cell Lines
3.7. Preliminary Tests Show That G. simplicifolia Seed Extracts Possess Antimicrobial Activity against S. aureus
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maffei, M. Plant Bioactive Molecules; Cambridge Scholars Publishing: Newcastle upon Tyne, UK, 2019; ISBN 1527526372. [Google Scholar]
- Jadhav, H.B.; Sablani, S.; Gogate, P.; Annapure, U.; Casanova, F.; Nayik, G.A.; Alaskar, K.; Sarwar, N.; Raina, I.A.; Ramniwas, S. Factors governing consumers buying behavior concerning nutraceutical product. Food Sci. Nutr. 2023; Early View. [Google Scholar] [CrossRef]
- Carnevale, G.; Di Viesti, V.; Zavatti, M.; Zanoli, P. Anxiolytic-like effect of Griffonia simplicifolia Baill. seed extract in rats. Phytomedicine 2011, 18, 848–851. [Google Scholar] [PubMed]
- Carnevale, G.; Di Viesti, V.; Zavatti, M.; Benelli, A.; Zanoli, P. Griffonia simplicifolia negatively affects sexual behavior in female rats. Phytomedicine 2010, 17, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Irvine, F.R. Woody plants of Ghana. In Woody plants of Ghana; Oxford University Press: Oxford, UK, 1961. [Google Scholar]
- Fellows, L.E.; Bell, E.A. 5-Hydroxy-L-tryptophan, 5-hydroxytryptamine and L-tryptophan-5-hydroxylase in Griffonia simplicifolia. Phytochemistry 1970, 9, 2389–2396. [Google Scholar] [CrossRef]
- Lemaire, P.A.; Adosraku, R.K. An HPLC method for the direct assay of the serotonin precursor, 5-hydroxytrophan, in seeds of Griffonia simplicifolia. Phytochem. Anal. 2002, 13, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Xin-Zhi, W.; Fei-Hua, W.U.; Wei, Q.U.; Liang, J.-Y. A new β-carboline alkaloid from the seeds of Griffonia simplicifolia. Chin. J. Nat. Med. 2013, 11, 401–405. [Google Scholar]
- Maffei, M.E. 5-Hydroxytryptophan (5-HTP): Natural occurrence, analysis, biosynthesis, biotechnology, physiology and toxicology. Int. J. Mol. Sci. 2020, 22, 181. [Google Scholar] [CrossRef]
- Jha, A.K.; Sit, N. Extraction of bioactive compounds from plant materials using combination of various novel methods: A review. Trends Food Sci. Technol. 2022, 119, 579–591. [Google Scholar] [CrossRef]
- Vigliante, I.; Mannino, G.; Maffei, M.E. Chemical characterization and DNA fingerprinting of Griffonia simplicifolia baill. Molecules 2019, 24, 1032. [Google Scholar] [CrossRef]
- Xie, D.-T.; Wang, Y.-Q.; Kang, Y.; Hu, Q.-F.; Su, N.-Y.; Huang, J.-M.; Che, C.-T.; Guo, J.-X. Microwave-assisted extraction of bioactive alkaloids from Stephania sinica. Sep. Purif. Technol. 2014, 130, 173–181. [Google Scholar] [CrossRef]
- Belwal, T.; Pandey, A.; Bhatt, I.D.; Rawal, R.S. Optimized microwave assisted extraction (MAE) of alkaloids and polyphenols from Berberis roots using multiple-component analysis. Sci. Rep. 2020, 10, 917. [Google Scholar] [CrossRef]
- Mannino, G.; Ricciardi, M.; Gatti, N.; Serio, G.; Vigliante, I.; Contartese, V.; Gentile, C.; Bertea, C.M. Changes in the Phytochemical Profile and Antioxidant Properties of Prunus persica Fruits after the Application of a Commercial Biostimulant Based on Seaweed and Yeast Extract. Int. J. Mol. Sci. 2022, 23, 15911. [Google Scholar] [CrossRef]
- Prior, R.L.; Fan, E.; Ji, H.; Howell, A.; Nio, C.; Payne, M.J.; Reed, J. Multi-laboratory validation of a standard method for quantifying proanthocyanidins in cranberry powders. J. Sci. Food Agric. 2010, 90, 1473–1478. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.K.; Srivastava, B.; Sahu, A. Spectrophotometric determination of Rauwolfia alkaloids: Estimation of reserpine in pharmaceuticals. Anal. Sci. 2004, 20, 571–573. [Google Scholar] [CrossRef] [PubMed]
- Agliassa, C.; Mannino, G.; Molino, D.; Cavalletto, S.; Contartese, V.; Bertea, C.M.; Secchi, F. A new protein hydrolysate-based biostimulant applied by fertigation promotes relief from drought stress in Capsicum annuum L. Plant Physiol. Biochem. 2021, 166, 1076–1086. [Google Scholar] [CrossRef] [PubMed]
- Mannino, G.; Maffei, M.E. Metabolomics-Based Profiling, Antioxidant Power, and Uropathogenic Bacterial Anti-Adhesion Activity of SP4TM, a Formulation with a High Content of Type-A Proanthocyanidins. Antioxidants 2022, 11, 1234. [Google Scholar] [CrossRef]
- Mannino, G.; Iovino, P.; Lauria, A.; Genova, T.; Asteggiano, A.; Notarbartolo, M.; Porcu, A.; Serio, G.; Chinigò, G.; Occhipinti, A. Bioactive triterpenes of protium heptaphyllum gum resin extract display cholesterol-lowering potential. Int. J. Mol. Sci. 2021, 22, 2664. [Google Scholar] [CrossRef]
- Demirci, F.; Guven, K.; Demirci, B.; Dadandi, M.Y.; Baser, K.H.C. Antibacterial activity of two Phlomis essential oils against food pathogens. Food Control 2008, 19, 1159–1164. [Google Scholar] [CrossRef]
- Gaglio, R.; Barbera, M.; Aleo, A.; Lommatzsch, I.; La Mantia, T.; Settanni, L. Inhibitory activity and chemical characterization of Daucus carota subsp. maximus essential oils. Chem. Biodivers. 2017, 14, e1600477. [Google Scholar] [CrossRef]
- Sati, S.C.; Joshi, S. Antibacterial activity of the Himalayan lichen Parmotrema nilgherrense extracts. Br. Microbiol. Res. J. 2011, 1, 26. [Google Scholar] [CrossRef]
- Muhamad, N.; Muhmed, S.A.; Yusoff, M.M.; Gimbun, J. Influence of solvent polarity and conditions on extraction of antioxidant, flavonoids and phenolic content from Averrhoa bilimbi. J. Food Sci. Eng. 2014, 4, 255–260. [Google Scholar]
- Hossain, M.A.; Shah, M.D.; Gnanaraj, C.; Iqbal, M. In vitro total phenolics, flavonoids contents and antioxidant activity of essential oil, various organic extracts from the leaves of tropical medicinal plant Tetrastigma from Sabah. Asian Pac. J. Trop. Med. 2011, 4, 717–721. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Singh, V.; Shukla, S.; Rai, A.K. Phenolic content and antioxidant capacity of selected cucurbit fruits extracted with different solvents. J. Nutr. Food Sci. 2016, 6, 565. [Google Scholar] [CrossRef]
- Chiavaro, E.; Cerretani, L.; Paradiso, V.M.; Summo, C.; Paciulli, M.; Gallina Toschi, T.; Caponio, F. Thermal and chemical evaluation of naturally auto-oxidised virgin olive oils: A correlation study. J. Sci. Food Agric. 2013, 93, 2909–2916. [Google Scholar] [CrossRef] [PubMed]
- Calinescu, I.; Asofiei, I.; Gavrila, A.I.; Trifan, A.; Ighigeanu, D.; Martin, D.; Matei, C.; Buleandra, M. Integrating microwave-assisted extraction of essential oils and polyphenols from rosemary and thyme leaves. Chem. Eng. Commun. 2017, 204, 965–973. [Google Scholar] [CrossRef]
- Chowdhary, P.; Gupta, A.; Gnansounou, E.; Pandey, A.; Chaturvedi, P. Current trends and possibilities for exploitation of Grape pomace as a potential source for value addition. Environ. Pollut. 2021, 278, 116796. [Google Scholar] [CrossRef]
- Zou, S.; Wang, C.; Li, J.; He, J.; Gao, X.; Chang, Y. Microwave assisted solid phase microextraction for extraction and selective enrichment of four alkaloids in lotus leaf. Sustain. Chem. Pharm. 2020, 18, 100345. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, Z.; Han, F.; Kang, X.; Gu, H.; Yang, L. Microwave-assisted method for simultaneous hydrolysis and extraction in obtaining ellagic acid, gallic acid and essential oil from Eucalyptus globulus leaves using Brönsted acidic ionic liquid [HO3S(CH2)4mim] HSO4. Ind. Crops Prod. 2016, 81, 152–161. [Google Scholar] [CrossRef]
- Cao, J.; Chen, L.; Li, M.; Cao, F.; Zhao, L.; Su, E. Efficient extraction of proanthocyanidin from Ginkgo biloba leaves employing rationally designed deep eutectic solvent-water mixture and evaluation of the antioxidant activity. J. Pharm. Biomed. Anal. 2018, 158, 317–326. [Google Scholar] [CrossRef]
- Letaief, T.; Garzoli, S.; Laghezza Masci, V.; Mejri, J.; Abderrabba, M.; Tiezzi, A.; Ovidi, E. Chemical composition and biological activities of tunisian Ziziphus lotus extracts: Evaluation of drying effect, solvent extraction, and extracted plant parts. Plants 2021, 10, 2651. [Google Scholar] [CrossRef]
- Towanou, R.; Konmy, B.; Yovo, M.; Dansou, C.C.; Dougnon, V.; Loko, F.S.; Akpovi, C.D.; Baba-Moussa, L. Phytochemical Screening, Antioxidant Activity, and Acute Toxicity Evaluation of Senna italica Extract Used in Traditional Medicine. J. Toxicol. 2023, 2023, 6405415. [Google Scholar] [CrossRef]
- Harrison, K.; Were, L.M. Effect of gamma irradiation on total phenolic content yield and antioxidant capacity of almond skin extracts. Food Chem. 2007, 102, 932–937. [Google Scholar] [CrossRef]
- Rusu, M.E.; Gheldiu, A.-M.; Mocan, A.; Moldovan, C.; Popa, D.-S.; Tomuta, I.; Vlase, L. Process optimization for improved phenolic compounds recovery from walnut (Juglans regia L.) septum: Phytochemical profile and biological activities. Molecules 2018, 23, 2814. [Google Scholar] [CrossRef] [PubMed]
- Diniyah, N.; Alam, M.B.; Lee, S.-H. Antioxidant potential of non-oil seed legumes of Indonesian’s ethnobotanical extracts. Arab. J. Chem. 2020, 13, 5208–5217. [Google Scholar] [CrossRef]
- Feliciano, R.P.; Shea, M.P.; Shanmuganayagam, D.; Krueger, C.G.; Howell, A.B.; Reed, J.D. Comparison of isolated cranberry (Vaccinium macrocarpon Ait.) proanthocyanidins to catechin and procyanidins A2 and B2 for use as standards in the 4-(dimethylamino) cinnamaldehyde assay. J. Agric. Food Chem. 2012, 60, 4578–4585. [Google Scholar] [CrossRef]
- Yanagida, A.; Murao, H.; Ohnishi-Kameyama, M.; Yamakawa, Y.; Shoji, A.; Tagashira, M.; Kanda, T.; Shindo, H.; Shibusawa, Y. Retention behavior of oligomeric proanthocyanidins in hydrophilic interaction chromatography. J. Chromatogr. A 2007, 1143, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Ramos, P.A.B.; Pereira, C.; Gomes, A.P.; Neto, R.T.; Almeida, A.; Santos, S.A.O.; Silva, A.M.S.; Silvestre, A.J.D. Chemical characterisation, antioxidant and antibacterial activities of Pinus pinaster ait. and Pinus pinea L. Bark polar extracts: Prospecting forestry by-products as renewable sources of bioactive compounds. Appl. Sci. 2022, 12, 784. [Google Scholar] [CrossRef]
- Flórez, N.; Conde, E.; Domínguez, H. Microwave assisted water extraction of plant compounds. J. Chem. Technol. Biotechnol. 2015, 90, 590–607. [Google Scholar] [CrossRef]
- Mannino, G.; Chinigò, G.; Serio, G.; Genova, T.; Gentile, C.; Munaron, L.; Bertea, C.M. Proanthocyanidins and where to find them: A meta-analytic approach to investigate their chemistry, biosynthesis, distribution, and effect on human health. Antioxidants 2021, 10, 1229. [Google Scholar] [CrossRef]
- Hertzman, P.A.; Blevins, W.L.; Mayer, J.; Greenfield, B.; Ting, M.; Gleich, G.J. Association of the eosinophilia–myalgia syndrome with the ingestion of tryptophan. N. Engl. J. Med. 1990, 322, 869–873. [Google Scholar] [CrossRef]
- GODA, Y.; SUZUKI, J.; MAITANI, T.; YOSHIHIRA, K.; TAKEDA, M.; UCHIYAMA, M. 3-Anilino-L-alanine, structural determination of UV-5, a contaminant in EMS-associated L-tryptophan samples. Chem. Pharm. Bull. 1992, 40, 2236–2238. [Google Scholar] [CrossRef]
- Yamaoka, K.A.; Miyasaka, N.; Inuo, G.; Saito, I.; Kolb, J.-P.; Fujita, K.; Kashiwazaki, S. 1,1′-Ethylidenebis (tryptophan)(peak E) induces functional activation of human eosinophils and interleukin 5 production from T lymphocytes: Association of eosinophilia-myalgia syndrome with al-tryptophan contaminant. J. Clin. Immunol. 1994, 14, 50–60. [Google Scholar] [CrossRef]
- Sato, F.; Hagiwara, Y.; Kawase, Y. Subchronic toxicity of 3-phenylamino alanine, an impurity in L-tryptophan reported to be associated with eosinophilia-myalgia syndrome. Arch. Toxicol. 1995, 69, 444–449. [Google Scholar] [CrossRef]
- Williamson, B.L.; Johnson, K.L.; Tomlinson, A.J.; Gleich, G.J.; Naylor, S. On-line HPLC-tandem mass spectrometry structural characterization of case-associated contaminants of L-tryptophan implicated with the onset of eosinophilia myalgia syndrome. Toxicol. Lett. 1998, 99, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Klarskov, K.; Johnson, K.L.; Benson, L.M.; Cragun, J.D.; Gleich, G.J.; Wrona, M.; Jiang, X.-R.; Dryhurst, G.; Naylor, S. Structural characterization of a case-implicated contaminant," Peak X," in commercial preparations of 5-hydroxytryptophan. J. Rheumatol. 2003, 30, 89–95. [Google Scholar] [PubMed]
- Tomek, P.; Palmer, B.D.; Flanagan, J.U.; Fung, S.-P.S.; Bridewell, D.J.A.; Jamie, J.F.; Ching, L.-M. Formation of an N-formylkynurenine-derived fluorophore and its use for measuring indoleamine 2, 3-dioxygenase 1 activity. Anal. Bioanal. Chem. 2013, 405, 2515–2524. [Google Scholar] [CrossRef] [PubMed]
- Humphries, K.; Dryhurst, G. Electrochemical oxidation of 5-hydroxytryptophan in acid solution. J. Pharm. Sci. 1987, 76, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Humphries, K.A.; Wrona, M.Z.; Dryhurst, G. Electrochemical and enzymatic oxidation of 5-hydroxytryptophan. J. Electroanal. Chem. 1993, 346, 377–403. [Google Scholar] [CrossRef]
- Mehta, A.; Prabhakar, M.; Kumar, P.; Deshmukh, R.; Sharma, P.L. Excitotoxicity: Bridge to various triggers in neurodegenerative disorders. Eur. J. Pharmacol. 2013, 698, 6–18. [Google Scholar] [CrossRef]
- Matute, C.; Domercq, M.; Sánchez-Gómez, M. Glutamate-mediated glial injury: Mechanisms and clinical importance. Glia 2006, 53, 212–224. [Google Scholar] [CrossRef]
- Pellegrini, N.; Vitaglione, P.; Granato, D.; Fogliano, V. Twenty-five years of total antioxidant capacity measurement of foods and biological fluids: Merits and limitations. J. Sci. Food Agric. 2020, 100, 5064–5078. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Negulescu, G.P. Methods for total antioxidant activity determination: A review. Biochem. Anal. Biochem. 2011, 1, 106. [Google Scholar] [CrossRef]
- Liang, N.; Kitts, D.D. Antioxidant property of coffee components: Assessment of methods that define mechanisms of action. Molecules 2014, 19, 19180–19208. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.P.; Martelli-Tosi, M.; Massarioli, A.P.; Melo, P.S.; Alencar, S.M.; Favaro-Trindade, C.S. Co-encapsulation of guaraná extracts and probiotics increases probiotic survivability and simultaneously delivers bioactive compounds in simulated gastrointestinal fluids. LWT 2022, 161, 113351. [Google Scholar] [CrossRef]
- Cardoso, R.R.; Neto, R.O.; dos Santos D’Almeida, C.T.; do Nascimento, T.P.; Pressete, C.G.; Azevedo, L.; Martino, H.S.D.; Cameron, L.C.; Ferreira, M.S.L.; de Barros, F.A.R. Kombuchas from green and black teas have different phenolic profile, which impacts their antioxidant capacities, antibacterial and antiproliferative activities. Food Res. Int. 2020, 128, 108782. [Google Scholar] [CrossRef]
- Lauria, A.; Mannino, S.; Gentile, C.; Mannino, G.; Martorana, A.; Peri, D. DRUDIT: Web-Based DRUgs DIscovery Tools to Design Small Molecules as Modulators of Biological Targets. Bioinformatics 2020, 36, 1562–1569. [Google Scholar] [CrossRef] [PubMed]
- Mannino, G.; Serio, G.; Gaglio, R.; Busetta, G.; La Rosa, L.; Lauria, A.; Settanni, L.; Gentile, C. Phytochemical Profile and Antioxidant, Antiproliferative, and Antimicrobial Properties of Rubus idaeus Seed Powder. Foods 2022, 11, 2605. [Google Scholar] [CrossRef]
- Vu, V.; Szewczyk, M.M.; Nie, D.Y.; Arrowsmith, C.H.; Barsyte-Lovejoy, D. Validating small molecule chemical probes for biological discovery. Annu. Rev. Biochem. 2022, 91, 61–87. [Google Scholar] [CrossRef]
- Regassa, H.; Sourirajan, A.; Kumar, V.; Pandey, S.; Kumar, D.; Dev, K. A review of medicinal plants of the himalayas with anti-proliferative activity for the treatment of various cancers. Cancers 2022, 14, 3898. [Google Scholar] [CrossRef]
- Akoto, C.O.; Acheampong, A.; Tagbor, P.D.; Bortey, K. Determination of the antimicrobial and antioxidant activities of the leaf extracts of Griffonia simplicifolia. J. Pharmacogn. Phytochem. 2020, 9, 537–545. [Google Scholar]
- Donkor, E.S. Cockroaches and food-borne pathogens. Environ. Health Insights 2020, 14, 1178630220913365. [Google Scholar] [CrossRef]
- Mbata, T.I.; Debiao, L.U.; Saikia, A. Antibacterial activity of the crude extract of Chinese green tea (Camellia sinensis) on Listeria monocytogenes. Afr. J. Biotechnol. 2008, 7, 1571–1573. [Google Scholar]
- Akoto, C.O.; Acheampong, A.; Boakye, Y.D.; Akwata, D.; Okine, M. In vitro anthelminthic, antimicrobial and antioxidant activities and FTIR analysis of extracts of Alchornea cordifolia leaves. J. Pharmacogn. Phytochem. 2019, 8, 2432–2442. [Google Scholar]
- Srivastava, P.; Upreti, D.K.; Dhole, T.N.; Srivastava, A.K.; Nayak, M.T. Antimicrobial property of extracts of Indian lichen against human pathogenic bacteria. Interdiscip. Perspect. Infect. Dis. 2013, 2013, 709348. [Google Scholar] [CrossRef]
- Teng, H.; Lee, W.Y. Antibacterial and antioxidant activities and chemical compositions of volatile oils extracted from Schisandra chinensis Baill. seeds using simultaneous distillation extraction method, and comparison with Soxhlet and microwave-assisted extraction. Biosci. Biotechnol. Biochem. 2014, 78, 79–85. [Google Scholar] [CrossRef]
- De Castro, C.; Parrilli, M.; Holst, O.; Molinaro, A. Microbe-associated molecular patterns in innate immunity: Extraction and chemical analysis of gram-negative bacterial lipopolysaccharides. Methods Enzymol. 2010, 480, 89–115. [Google Scholar] [PubMed]
- Chan, E.W.C.; Lim, Y.Y.; Omar, M. Antioxidant and antibacterial activity of leaves of Etlingera species (Zingiberaceae) in Peninsular Malaysia. Food Chem. 2007, 104, 1586–1593. [Google Scholar] [CrossRef]
- Alves, F.S.; Cruz, J.N.; de Farias Ramos, I.N.; do Nascimento Brandão, D.L.; Queiroz, R.N.; da Silva, G.V.; da Silva, G.V.; Dolabela, M.F.; da Costa, M.L.; Khayat, A.S. Evaluation of antimicrobial activity and cytotoxicity effects of extracts of piper nigrum L. and piperine. Separations 2022, 10, 21. [Google Scholar] [CrossRef]
- Aguilar-Villalva, R.; Molina, G.A.; España-Sánchez, B.L.; Díaz-Peña, L.F.; Elizalde-Mata, A.; Valerio, E.; Azanza-Ricardo, C.; Estevez, M. Antioxidant capacity and antibacterial activity from Annona cherimola phytochemicals by ultrasound-assisted extraction and its comparison to conventional methods. Arab. J. Chem. 2021, 14, 103239. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DPPH | ABTS | FRAP | ||
|---|---|---|---|---|
| Acetone | Soxhlet | 0.201 (0.001) g | 0.196 (0.004) l | 0.003 (0.001) i |
| MA95 | 0.197 (0.003) g | 0.151 (0.001) l | 0.025 (0.001) i | |
| MA70 | 0.749 (0.023) e | 8.884 (0.217) f | 1.775 (0.055) c | |
| MA50 | 1.568 (0.036) d | 22.493 (0.146) b | 1.153 (0.023) d | |
| Ethanol | Soxhlet | 0.618 (0.020) ef | 4.482 (0.049) h | 0.421 (0.010) f |
| ME95 | 0.373 (0.035) fd | 10.646 (0.196) e | 0.255 (0.005) g | |
| ME70 | 3.811 (0.157) b | 19.889 (0.882) d | 3.111 (0.032) b | |
| ME50 | 5.187 (0.356) a | 22.561 (0.265) b | 4.568 (0.047) a | |
| Methanol | Soxhlet | 0.318 (0.023) fg | 3.753 (0.025) h | 0.553 (0.008) e |
| MM95 | 1.956 (0.075) c | 2.297 (0.058) i | 0.317 (0.006) g | |
| MM70 | 1.923 (0.014) c | 25.088 (0.218) a | 0.275 (0.008) g | |
| MM50 | 3.824 (0.097) b | 21.235 (0.244) c | 0.259 (0.011) g | |
| MAE | 0.145 (0.011) g | 5.987 (0.448) g | 0.102 (0.002) h | |
| HeLa | HepG2 | MCF-7 | ||
|---|---|---|---|---|
| Acetone | Soxhlet | >2000 | >2000 | >2000 |
| MA95 | >2000 | >2000 | >2000 | |
| MA70 | 1141.97 (35.7) a | 1076.74 (42.16) a | 1332.55 (33.01) a | |
| MA50 | 1189.55 (47.43) a | 1152.66 (37.77) a | 922.96 (35.26) b | |
| Ethanol | Soxhlet | >2000 | >2000 | >2000 |
| ME95 | >2000 | >2000 | >2000 | |
| ME70 | 784.6 (10.55) b | 634.32 (7.74) c | 760.5 (20.26) c | |
| ME50 | 1042.43 (11.53) a | 792.4 (22.13) b | 789.4 (18.21) c | |
| Methanol | Soxhlet | 649.88 (13.82) c | 809.52 (11.22) b | 701.84 (11.06) d |
| MM95 | 693.45 (22.68) c | 809.65 (25.03) b | 1029.25 (34.72) e | |
| MM70 | 603.15 (21.08) c | 803.69 (23.88) b | 719.59 (22.49) cd | |
| MM50 | 1072.89 (30.66) a | 777.16 (24.01) b | 1460.73 (39.11) a | |
| MAE | 1580.32 (37.27) d | 1744.3 (40.34) d | 1654.32 (48.62) f | |
| Solvents | Samples | Inhibition (mm) | |
|---|---|---|---|
| E. coli ATCC25922 | S. aureus ATCC33862 | ||
| Acetone | Soxhlet | n.d. | 16.9 ± 0.2 |
| MA50 | n.d. | n.d. | |
| MA70 | n.d. | 11.6 ± 0.1 | |
| MA95 | n.d. | 14.4 ± 0.2 | |
| n.d. | |||
| Ethanol | Soxhlet | n.d. | 18.2 ± 0.3 |
| ME50 | n.d. | n.d. | |
| ME70 | n.d. | 13.3 ± 0.2 | |
| ME95 | n.d. | 16.1 ± 0.4 | |
| n.d. | |||
| Methanol | Soxhlet | n.d. | 17.5 ± 0.3 |
| MM50 | n.d. | n.d. | |
| MM70 | n.d. | 12.3 ± 0.2 | |
| MM95 | n.d. | 15.8 ± 0.4 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mannino, G.; Serio, G.; Gaglio, R.; Maffei, M.E.; Settanni, L.; Di Stefano, V.; Gentile, C. Biological Activity and Metabolomics of Griffonia simplicifolia Seeds Extracted with Different Methodologies. Antioxidants 2023, 12, 1709. https://doi.org/10.3390/antiox12091709
Mannino G, Serio G, Gaglio R, Maffei ME, Settanni L, Di Stefano V, Gentile C. Biological Activity and Metabolomics of Griffonia simplicifolia Seeds Extracted with Different Methodologies. Antioxidants. 2023; 12(9):1709. https://doi.org/10.3390/antiox12091709
Chicago/Turabian StyleMannino, Giuseppe, Graziella Serio, Raimondo Gaglio, Massimo E. Maffei, Luca Settanni, Vita Di Stefano, and Carla Gentile. 2023. "Biological Activity and Metabolomics of Griffonia simplicifolia Seeds Extracted with Different Methodologies" Antioxidants 12, no. 9: 1709. https://doi.org/10.3390/antiox12091709
APA StyleMannino, G., Serio, G., Gaglio, R., Maffei, M. E., Settanni, L., Di Stefano, V., & Gentile, C. (2023). Biological Activity and Metabolomics of Griffonia simplicifolia Seeds Extracted with Different Methodologies. Antioxidants, 12(9), 1709. https://doi.org/10.3390/antiox12091709