

Antifungal Action of Arabidopsis thaliana TCP21 via Induction of Oxidative Stress and Apoptosis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Cells and Growth Conditions
2.2. Purification and Characterization of Recombinant AtTCP21
2.3. Transmission Electron Microscopy (TEM)
2.4. Antifungal Assay
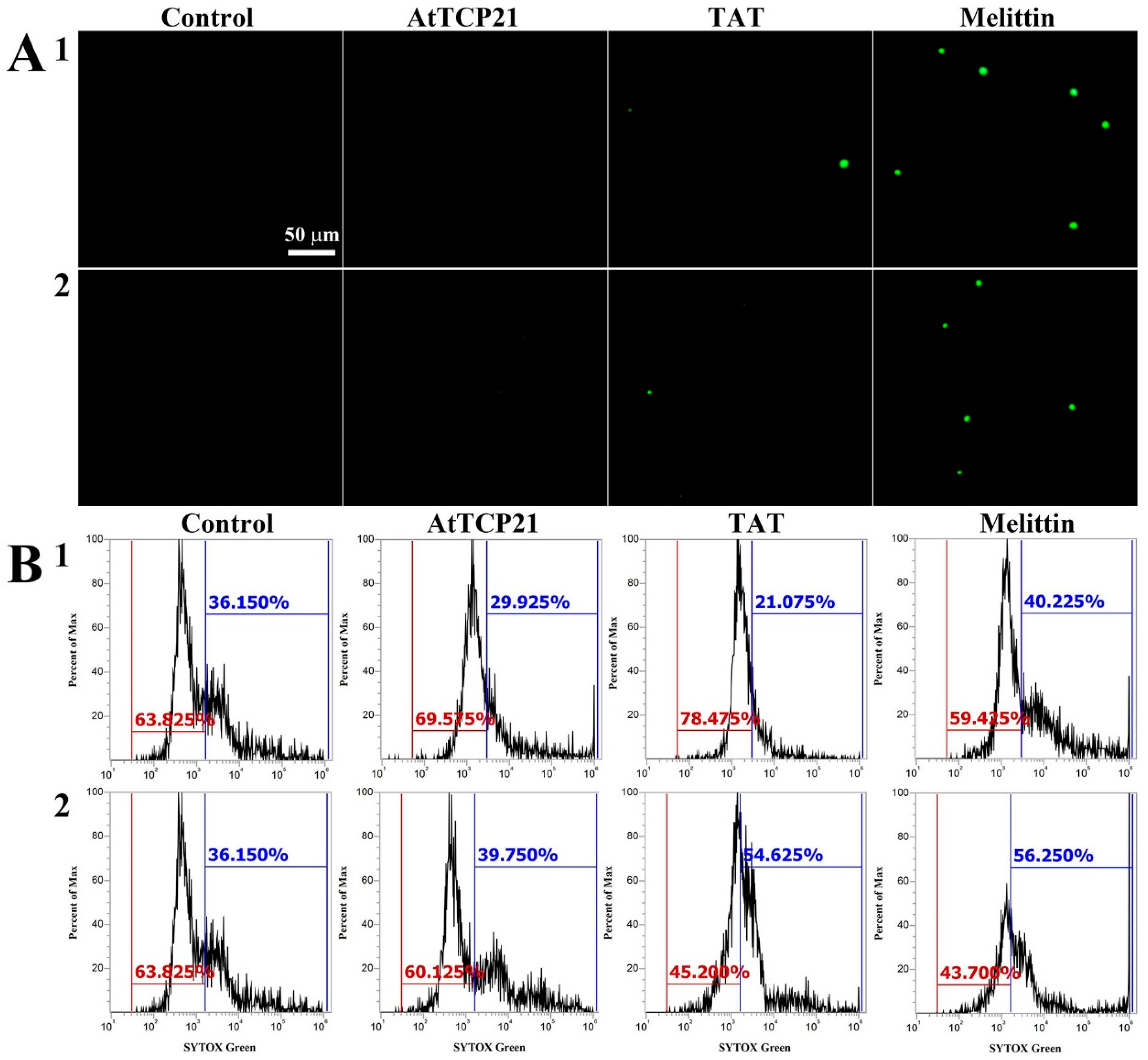
2.5. SYTOX Green Uptake Assay
2.6. Live/Dead Cell Assay
2.7. Confocal Laser Scanning Microscopy (CLSM)
2.8. Reactive Oxygen Species (ROS) Assessment
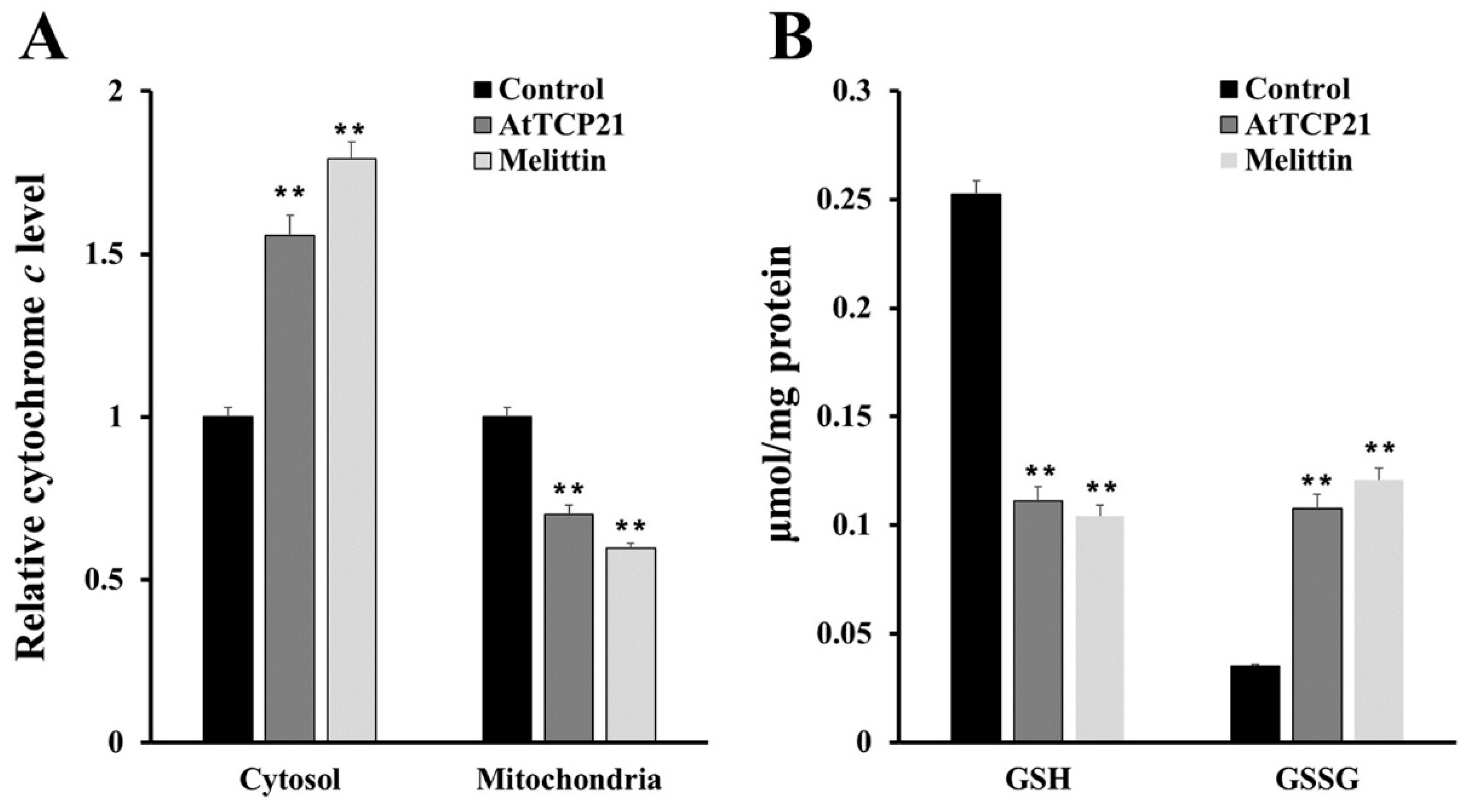
2.9. Quantification of Reduced Glutathione (GSH)
2.10. Measurement of Mitochondrial Superoxide and Membrane Potential
2.11. Cytochrome c Release Assay
2.12. Apoptosis Assay
2.13. Caspase 3/7 Activation Assay
2.14. Nuclear Staining with Hoechst 33342
2.15. Scanning Electron Microscopy (SEM)
2.16. Statistical Analysis
3. Results and Discussion
3.1. Purification and Identification of an Arabidopsis TCP Protein with Antifungal Activity
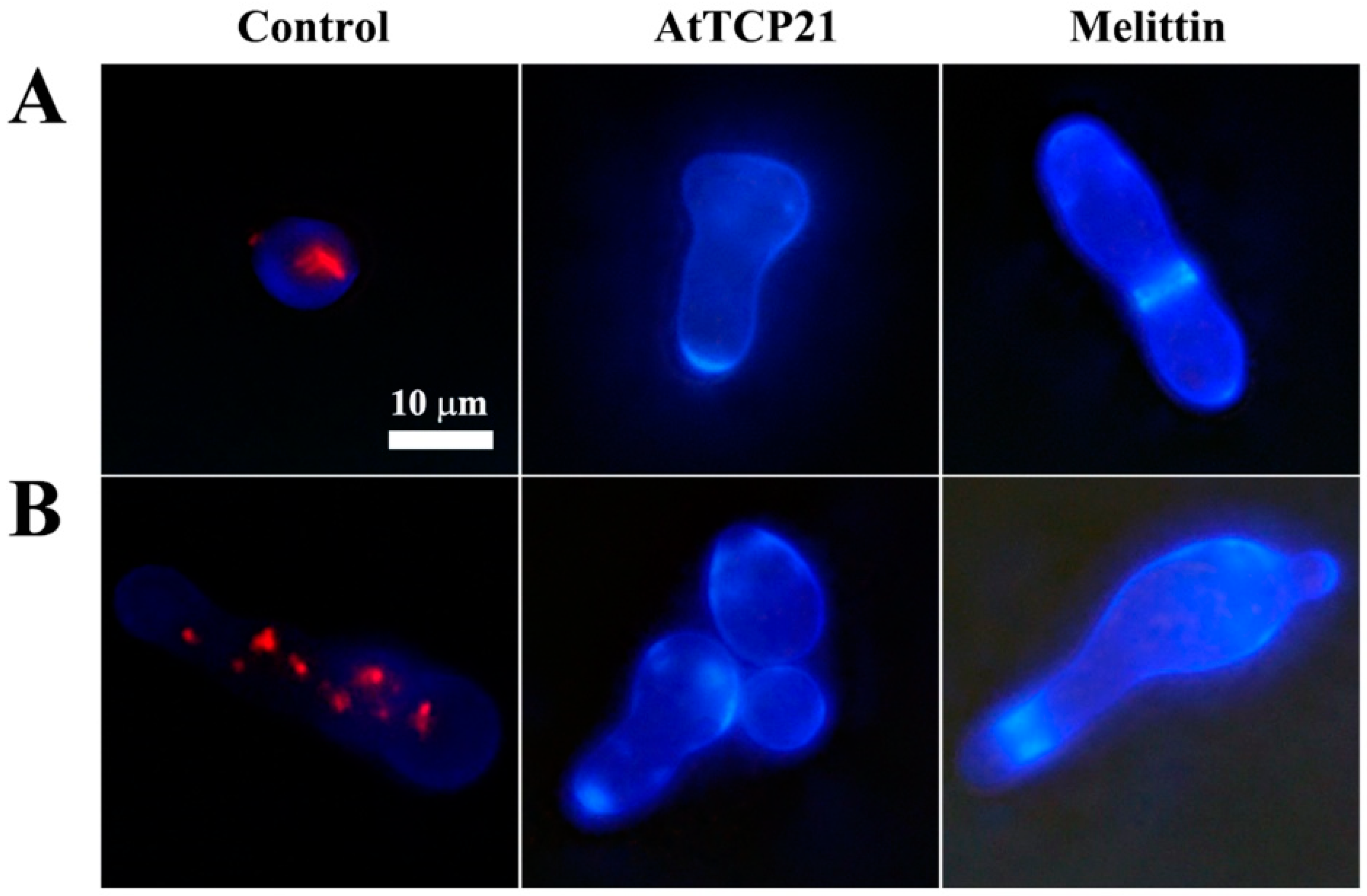
3.2. Cellular Distribution of AtTCP21 in Fungal Cells
3.3. Cellular Uptake of AtTCP21 without Alteration of Membrane Integrity
3.4. AtTCP21-Induced Intracellular ROS Generation
3.5. AtTCP21 Induces Apoptosis and Activates Caspase 3/7
3.6. Morphological Alterations Induced by AtTCP21 in C. gloeosporioides
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.; Wertheim, H.F.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H.; et al. Antibiotic resistance-the need for global solutions. Lancet Infect. Dis. 2013, 13, 1057–1098. [Google Scholar] [CrossRef] [PubMed]
- Erdem Büyükkiraz, M.; Kesmen, Z. Antimicrobial peptides (AMPs): A promising class of antimicrobial compounds. J. Appl. Microbiol. 2022, 132, 1573–1596. [Google Scholar] [CrossRef] [PubMed]
- Stotz, H.U.; Thomson, J.G.; Wang, Y. Plant defensins: Defense, development and application. Plant Signal Behav. 2009, 4, 1010–1012. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Kaur, J.; Sharma, G.; Kashyap, P. Plant derived antimicrobial peptides: Mechanism of target, isolation techniques, sources and pharmaceutical applications. J. Food Biochem. 2022, 46, e14348. [Google Scholar] [CrossRef]
- Bakare, O.O.; Gokul, A.; Fadaka, A.O.; Wu, R.; Niekerk, L.A.; Barker, A.M.; Keyster, M.; Klein, A. Plant Antimicrobial Peptides (PAMPs): Features, Applications, Production, Expression, and Challenges. Molecules 2022, 27, 3703. [Google Scholar] [CrossRef]
- Lima, A.M.; Azevedo, M.I.G.; Sousa, L.M.; Oliveira, N.S.; Andrade, C.R.; Freitas, C.D.T.; Souza, P.F.N. Plant antimicrobial peptides: An overview about classification, toxicity and clinical applications. Int. J. Biol. Macromol. 2022, 214, 10–21. [Google Scholar] [CrossRef]
- Doebley, J. The genetics of maize evolution. Annu. Rev. Genet. 2004, 38, 37–59. [Google Scholar] [CrossRef]
- Hileman, L.C.; Baum, D.A. Why do paralogs persist? Molecular evolution of CYCLOIDEA and related floral symmetry genes in Antirrhineae (Veronicaceae). Mol. Biol. Evol. 2003, 20, 591–600. [Google Scholar] [CrossRef]
- Kosugi, S.; Ohashi, Y. PCF1 and PCF2 specifically bind to cis elements in the rice proliferating cell nuclear antigen gene. Plant Cell 1997, 9, 1607–1619. [Google Scholar]
- Viola, I.L.; Alem, A.L.; Jure, R.M.; Gonzalez, D.H. Physiological Roles and Mechanisms of Action of Class I TCP Transcription Factors. Int. J. Mol. Sci. 2023, 24, 5437. [Google Scholar] [CrossRef]
- Li, S. The Arabidopsis thaliana TCP transcription factors: A broadening horizon beyond development. Plant Signal Behav. 2015, 10, e1044192. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Hwarari, D.; Ma, H.; Xu, H.; Yang, L.; Luo, Y. Genomic survey of TCP transcription factors in plants: Phylogenomics, evolution and their biology. Front. Genet. 2022, 13, 1060546. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wen, X.; Huang, X.; Wei, M.; Chen, H.; Yu, Y.; Dai, S. Genome-Wide Identification and Characterization of TCP Gene Family Members in Melastoma Candidum. Molecules 2022, 27, 9036. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, S.; Khan, S.; Usharani, T.R.; Tiwari, S. Analysis of TCP Transcription Factors Revealed Potential Roles in Plant Growth and Fusarium oxysporum f.sp. cubense Resistance in Banana (cv. Rasthali). Appl. Biochem. Biotechnol. 2022, 194, 5456–5473. [Google Scholar] [CrossRef]
- Wu, F.; Qi, J.; Meng, X.; Jin, W. miR319c acts as a positive regulator of tomato against Botrytis cinerea infection by targeting TCP29. Plant Sci. 2020, 300, 110610. [Google Scholar] [CrossRef]
- Lopez, J.A.; Sun, Y.; Blair, P.B.; Mukhtar, M.S. TCP three-way handshake: Linking developmental processes with plant immunity. Trends Plant Sci. 2015, 20, 238–245. [Google Scholar] [CrossRef]
- Park, S.C.; Kim, I.R.; Kim, J.Y.; Lee, Y.; Kim, E.J.; Jung, J.H.; Jung, Y.J.; Jang, M.K.; Lee, J.R. Molecular mechanism of Arabidopsis thaliana profilins as antifungal proteins. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2545–2554. [Google Scholar] [CrossRef]
- Son, H.; Jung, Y.J.; Park, S.C.; Kim, I.R.; Park, J.H.; Jang, M.K.; Lee, J.R. Functional Characterization of an Arabidopsis Profilin Protein as a Molecular Chaperone under Heat Shock Stress. Molecules 2022, 27, 5771. [Google Scholar] [CrossRef]
- Park, S.C.; Lim, H.S.; Mun, S.E.; Jung, Y.J.; Yoon, A.M.; Son, H.; Kim, C.M.; Choo, Y.K.; Lee, J.R. Potent Antifungal Functions of a Living Modified Organism Protein, CP4-EPSPS, against Pathogenic Fungal Cells. Molecules 2023, 28, 4289. [Google Scholar] [CrossRef]
- Park, S.C.; Kim, I.R.; Kim, J.Y.; Lee, Y.; Yoo, S.H.; Jung, J.H.; Cheong, G.W.; Lee, S.Y.; Jang, M.K.; Lee, J.R. Functional Characterization of a Rice Thioredoxin Protein OsTrxm and Its Cysteine Mutant Variant with Antifungal Activity. Antioxidants 2019, 8, 598. [Google Scholar] [CrossRef]
- Petre, B. Toward the Discovery of Host-Defense Peptides in Plants. Front. Immunol. 2020, 11, 1825. [Google Scholar] [CrossRef] [PubMed]
- Ninkuu, V.; Yan, J.; Fu, Z.; Yang, T.; Ziemah, J.; Ullrich, M.S.; Kuhnert, N.; Zeng, H. Lignin and Its Pathway-Associated Phytoalexins Modulate Plant Defense against Fungi. J. Fungi 2022, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Maximiano, M.R.; Franco, O.L. Biotechnological applications of versatile plant lipid transfer proteins (LTPs). Peptides 2021, 140, 170531. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, M.; de la Canal, L.; Regente, M. Plant Antifungal Lectins: Mechanism of Action and Targets on Human Pathogenic Fungi. Curr. Protein Pept. Sci. 2020, 21, 284–294. [Google Scholar] [CrossRef]
- Shen, J.; Zeng, Y.; Zhuang, X.; Sun, L.; Yao, X.; Pimpl, P.; Jiang, L. Organelle pH in the Arabidopsis endomembrane system. Mol. Plant 2013, 6, 1419–1437. [Google Scholar] [CrossRef]
- Li, T.; Li, L.; Du, F.; Sun, L.; Shi, J.; Long, M.; Chen, Z. Activity and Mechanism of Action of Antifungal Peptides from Microorganisms: A Review. Molecules 2021, 26, 3438. [Google Scholar] [CrossRef]
- Parisi, K.; Shafee, T.M.A.; Quimbar, P.; van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. The evolution, function and mechanisms of action for plant defensins. Semin. Cell Dev. Biol. 2019, 88, 107–118. [Google Scholar] [CrossRef]
- Schutters, K.; Reutelingsperger, C. Phosphatidylserine targeting for diagnosis and treatment of human diseases. Apoptosis 2010, 15, 1072–1082. [Google Scholar] [CrossRef]
- Sharon, A.; Finkelstein, A.; Shlezinger, N.; Hatam, I. Fungal apoptosis: Function, genes and gene function. FEMS Microbiol. Rev. 2009, 33, 833–854. [Google Scholar] [CrossRef]
- Lakhani, S.A.; Masud, A.; Kuida, K.; Porter, G.A., Jr.; Booth, C.J.; Mehal, W.Z.; Inayat, I.; Flavell, R.A. Caspases 3 and 7: Key mediators of mitochondrial events of apoptosis. Science 2006, 311, 847–851. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungi | IC50 (μg/mL) | |
|---|---|---|
| AtTCP21 | Melittin | |
| Mold | ||
| C. gloeosporioides | 3.1 | 12.5 |
| F. graminearum | 25 | 12.5 |
| F. moniliforme | 50 | 25 |
| F. oxysporum | 25 | 12.5 |
| F. solani | 25 | 12.5 |
| T. harzianum | 25 | 25 |
| T. viride | 50 | 25 |
| Yeast | ||
| C. albicans | 100 | 6.3 |
| C. tropicalis | 100 | 12.5 |
| T. beigelii | 50 | 6.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.-C.; Yoon, A.-M.; Kim, Y.-M.; Lee, M.-Y.; Lee, J.R. Antifungal Action of Arabidopsis thaliana TCP21 via Induction of Oxidative Stress and Apoptosis. Antioxidants 2023, 12, 1767. https://doi.org/10.3390/antiox12091767
Park S-C, Yoon A-M, Kim Y-M, Lee M-Y, Lee JR. Antifungal Action of Arabidopsis thaliana TCP21 via Induction of Oxidative Stress and Apoptosis. Antioxidants. 2023; 12(9):1767. https://doi.org/10.3390/antiox12091767
Chicago/Turabian StylePark, Seong-Cheol, A-Mi Yoon, Young-Min Kim, Min-Young Lee, and Jung Ro Lee. 2023. "Antifungal Action of Arabidopsis thaliana TCP21 via Induction of Oxidative Stress and Apoptosis" Antioxidants 12, no. 9: 1767. https://doi.org/10.3390/antiox12091767
APA StylePark, S. -C., Yoon, A. -M., Kim, Y. -M., Lee, M. -Y., & Lee, J. R. (2023). Antifungal Action of Arabidopsis thaliana TCP21 via Induction of Oxidative Stress and Apoptosis. Antioxidants, 12(9), 1767. https://doi.org/10.3390/antiox12091767