
Boesenbergia rotunda and Its Pinostrobin for Atopic Dermatitis: Dual 5-Lipoxygenase and Cyclooxygenase-2 Inhibitor and Its Mechanistic Study through Steady-State Kinetics and Molecular Modeling


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Chemicals
2.2. Extraction of Essential Oils by Hydro Distillation
2.3. Ultrasound-Assisted Extraction (UAE)
2.4. Isolation and Structure Elucidation of Pinostrobin
2.5. Gas Chromatography–Mass Spectrometry Analysis
2.6. Antibacterial Activity
2.7. Determination of Antioxidant Activities
2.7.1. 2,2-Diphenyl-1-Picrylhydrazyl Radical (DPPH) Scavenging Activity
2.7.2. Nitric Oxide (NO) Radical Scavenging Activity
2.7.3. Ferric Reducing Antioxidant Power (FRAP)
2.7.4. Total Phenolic Content (TPC) Determination
2.7.5. Total Flavonoid Content (TFC) Determination
2.8. In Vitro Anti-Inflammatory Activities
2.8.1. Cyclooxygenase-2 Inhibitory Assay
2.8.2. 5-Lipoxygenase Inhibitory Assay
2.8.3. Steady-State Inhibition Kinetic Study of Pinostrobin to 5-Lipoxygenase
2.8.4. Molecular Modeling of Pinostrobin to 5-Lipoxygenase
2.9. Statistical Analysis
3. Results
3.1. Preparation of Plant Extracts and Essential Oils
3.2. Antibacterial Activity of Plant Extracts and Essential Oils
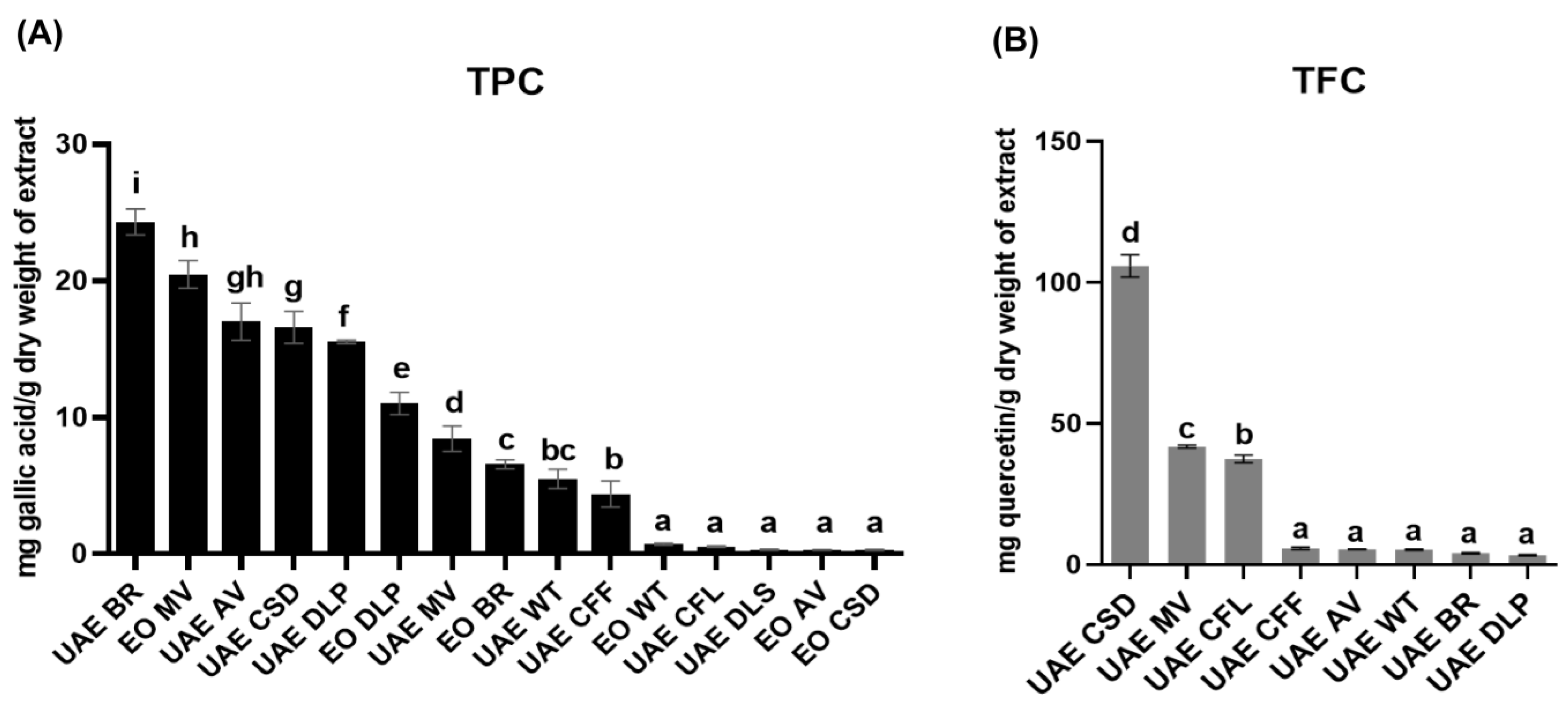
3.3. Antioxidant Potentials and Determination of Total Phenolic Content and Total Flavonoid Content
3.4. Gas Chromatography–Mass Spectrometry Analysis of Phytochemicals
3.5. Human Recombinant Cyclooxygenase-2 Inhibitory Activity of B. rotunda Essential Oils, Hexane Extract, and Pinostrobin
3.6. Human Recombinant 5-Lipoxygenase Inhibitory Activity of B. rotunda Essential Oils, Hexane Extract, and Pinostrobin, and Steady-State Inhibition Kinetic Study
3.7. Molecular Modeling of Pinostrobin and Nordihydroguaiaretic Acid to 5-Lipoxygenase
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Morgner, B.; Tittelbach, J.; Wiegand, C. Induction of psoriasis- and atopic dermatitis-like phenotypes in 3D skin equivalents with a fibroblast-derived matrix. Sci. Rep. 2023, 13, 1807. [Google Scholar] [CrossRef] [PubMed]
- Ramos Campos, E.V.; Proença, P.L.D.F.; Doretto-Silva, L.; Andrade-Oliveira, V.; Fraceto, L.F.; de Araujo, D.R. Trends in nanoformulations for atopic dermatitis treatment. Expert Opin. Drug Deliv. 2020, 17, 1615–1630. [Google Scholar] [CrossRef] [PubMed]
- Mayba, J.N.; Gooderham, M.J. Review of atopic dermatitis and topical therapies. J. Cutan. Med. Surg. 2017, 21, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, C.; Hipler, U.C.; Boldt, S.; Strehle, J.; Wollina, U. Skin-protective effects of a zinc oxide-functionalized textile and its relevance for atopic dermatitis. Clin. Cosmet. Investig. Dermatol. 2013, 6, 115–121. [Google Scholar] [PubMed]
- Atherton, D.J. Topical corticosteroids in atopic dermatitis. BMJ 2003, 327, 942. [Google Scholar] [CrossRef] [PubMed]
- Flower, R. Steroidal antiinflammatory drugs as inhibitors of phospholipase A2. Adv. Prostag. Thrombox. Res. 1978, 3, 105–112. [Google Scholar]
- Chew, Y.-L.; Khor, M.-A.; Xu, Z.; Lee, S.-K.; Keng, J.-W.; Sang, S.-H.; Akowuah, G.A.; Goh, K.W.; Liew, K.B.; Ming, L.C. Cassia alata, Coriandrum sativum, Curcuma longa and Azadirachta indica: Food Ingredients as Complementary and Alternative Therapies for Atopic Dermatitis-A Comprehensive Review. Molecules 2022, 27, 5475. [Google Scholar] [CrossRef]
- Reynolds, K.A.; Juhasz, M.L.W.; Mesinkovska, N.A. The role of oral vitamins and supplements in the management of atopic dermatitis: A systematic review. Int. J. Dermatol. 2019, 58, 1371–1376. [Google Scholar] [CrossRef]
- Javanbakht, M.H.; Keshavarz, S.A.; Djalali, M.; Siassi, F.; Eshraghian, M.R.; Firooz, A.; Seirafi, H.; Ehsani, A.H.; Chamari, M.; Mirshafiey, A. Randomized controlled trial using vitamins E and D supplementation in atopic dermatitis. J. Dermatol. Treat. 2011, 22, 144–150. [Google Scholar] [CrossRef]
- Oh, S.Y.; Chung, J.; Kim, M.K.; Kwon, S.O.; Cho, B.H. Antioxidant nutrient intakes and corresponding biomarkers associated with the risk of atopic dermatitis in young children. Eur. J. Clin. Nutr. 2010, 64, 245–252. [Google Scholar] [CrossRef]
- Schlichte, M.J.; Vandersall, A.; Katta, R. Diet and eczema: A review of dietary supplements for the treatment of atopic dermatitis. Dermatol. Pract. Concept. 2016, 6, 23–29. [Google Scholar] [CrossRef]
- Celik, V.; Beken, B.; Yazicioglu, M.; Ozdemir, P.G.; Sut, N. Do traditional fermented foods protect against infantile atopic dermatitis. Pediatr. Allergy Immunol. 2019, 30, 540–546. [Google Scholar] [CrossRef]
- Chen, D.; Chen, G.; Sun, Y.; Zeng, X.; Ye, H. Physiological genetics, chemical composition, health benefits and toxicology of tea (Camellia sinensis L.) flower: A review. Food Res. Int. 2020, 137, 109584. [Google Scholar] [CrossRef]
- Tang, Y.-Y.; He, X.-M.; Sun, J.; Li, C.-B.; Li, L.; Sheng, J.-F.; Xin, M.; Li, Z.-C.; Zheng, F.-J.; Liu, G.-M.; et al. Polyphenols and Alkaloids in Byproducts of Longan Fruits (Dimocarpus Longan Lour.) and Their Bioactivities. Molecules 2019, 24, 1186. [Google Scholar] [CrossRef]
- Eng-Chong, T.; Yean-Kee, L.; Chin-Fei, C.; Choon-Han, H.; Sher-Ming, W.; Li-Ping, C.T.; Gen-Teck, F.; Khalid, N.; Abd Rahman, N.; Karsani, S.A.; et al. Boesenbergia rotunda: From Ethnomedicine to Drug Discovery. Evid. Based Complement. Altern. Med. 2012, 2012, 473637. [Google Scholar] [CrossRef]
- Rosdianto, A.M.; Puspitasari, I.M.; Lesmana, R.; Levita, J. Inhibitory Activity of Boesenbergia rotunda (L.) Mansf. Rhizome towards the Expression of Akt and NF-KappaB p65 in Acetic Acid-Induced Wistar Rats. Evid. Based Complement. Altern. Med. 2020, 2020, 6940313. [Google Scholar] [CrossRef]
- Subbaiah, K.V.; Savithramma, N. Bio-prospecting and documentation of traditional medicinal plants used to treat leprosy and eczema by ethnic groups of Kurnool district, Andhra Pradesh. Asian J. Pharm. Clin. Res. 2011, 4, 4488–4490. [Google Scholar]
- Maldini, M.; Sosa, S.; Montoro, P.; Giangaspero, A.; Balick, M.J.; Pizza, C.; Loggia, R.D. Screening of the topical anti-inflammatory activity of the bark of Acacia cornigera Willdenow, Byrsonima crassifolia Kunth, Sweetia panamensis Yakovlev and the leaves of Sphagneticola trilobata Hitchcock. J. Ethnopharmacol. 2009, 122, 430–433. [Google Scholar] [CrossRef]
- Saqib, S.; Ullah, F.; Naeem, M.; Younas, M.; Ayaz, A.; Ali, S.; Zaman, W. Mentha: Nutritional and Health Attributes to Treat Various Ailments Including Cardiovascular Diseases. Molecules 2022, 27, 6728. [Google Scholar] [CrossRef]
- El Menyiy, N.; Mrabti, H.N.; El Omari, N.; Bakili, A.E.; Bakrim, S.; Mekkaoui, M.; Balahbib, A.; Amiri-Ardekani, E.; Ullah, R.; Alqahtani, A.S.; et al. Medicinal Uses, Phytochemistry, Pharmacology, and Toxicology of Mentha spicata. Evid. Based Complement. Altern. Med. 2022, 2022, 7990508. [Google Scholar] [CrossRef] [PubMed]
- Hayat, M.Q.; Khan, M.A.; Ashraf, M.; Jabeen, S. Ethnobotany of the Genus artemisia L. (Asteraceae) in Pakistan. Ethnobot. Res. Appl. 2009, 7, 147–162. [Google Scholar] [CrossRef]
- Pribowo, A.; Girish, J.; Gustiananda, M.; Nandhira, R.G.; Hartrianti, P. Potential of Tamanu (Calophyllum inophyllum) Oil for Atopic Dermatitis Treatment. Evid. Based Complement. Altern. Med. 2021, 2021, 6332867. [Google Scholar] [CrossRef]
- Yanes, D.A.; Mosser-Goldfarb, J.L. Emerging therapies for atopic dermatitis: The prostaglandin/leukotriene pathway. J. Am. Acad. Dermatol. 2018, 78, S71–S75. [Google Scholar] [CrossRef]
- Andoh, T.; Haza, S.; Saito, A.; Kuraishi, Y. Involvement of leukotriene B4 in spontaneous itch-related behaviour in NC mice with atopic dermatitis-like skin lesions. Exp. Dermatol. 2011, 20, 894–898. [Google Scholar] [CrossRef]
- Chari, S.; Clark-Loeser, L.; Shupack, J.; Washenik, K. A Role for Leukotriene Antagonists in Atopic Dermatitis? Am. J. Clin. Dermatol. 2001, 2, 1–6. [Google Scholar] [CrossRef]
- Fleischer, A.; Cincinnati, O.; Nam, K.; Kim, J.; Ahn, K.; Kang, H.; Choi, J.; Choi, B.J.; Lee, S.; Jung, C.; et al. Q301 (Zileuton) cream demonstrates superiority to vehicle in improving atopic dermatitis: Results from a phase 2A trial. J. Am. Acad. Dermatol. 2019, 81, AB112. [Google Scholar]
- Honda, T.; Kabashima, K. Prostanoids in allergy. Allergol. Int. 2015, 64, 11–16. [Google Scholar] [CrossRef]
- Moreno, A.S.; McPhee, R.; Arruda, L.K.; Howell, M.D. Targeting the T Helper 2 Inflammatory Axis in Atopic Dermatitis. Int. Arch. Allergy Immunol. 2016, 171, 71–80. [Google Scholar] [CrossRef]
- Tang, L.; Li, X.-L.; Deng, Z.-X.; Xiao, Y.; Cheng, Y.-H.; Li, J.; Ding, H. Conjugated linoleic acid attenuates 2, 4-dinitrofluorobenzene-induced atopic dermatitis in mice through dual inhibition of COX-2/5-LOX and TLR4/NF-κB signaling. J. Nut. Biochem. 2020, 81, 108379. [Google Scholar] [CrossRef]
- Carola, C.; Salazar, A.; Rakers, C.; Himbert, F.; Do, Q.T.; Bernard, P.; von Hagen, J. A Cornflower Extract Containing N-Feruloylserotonin Reduces Inflammation in Human Skin by Neutralizing CCL17 and CCL22 and Inhibiting COX-2 and 5-LOX. Mediat. Inflamm. 2021, 2021, 6652791. [Google Scholar] [CrossRef] [PubMed]
- Commission, B.P. British Pharmacopoeia; The Stationary Office: London, UK, 1970. [Google Scholar]
- European Committee for Antimicrobial Susceptibility Testing of the European Society of Clinical Microbiology and Infectious Diseases. Determination of minimum inhibitory concentrations (MICs) of antibacterial agents by broth dilution. Clin. Microbiol. Infect. 2003, 9, 9–15. [Google Scholar]
- Thadhani, V.M.; Choudhary, M.I.; Ali, S.; Omar, I.; Siddique, H.; Karunaratne, V. Antioxidant activity of some lichen metabolites. Nat. Prod. Res. 2011, 25, 1827–1837. [Google Scholar] [CrossRef]
- Chew, C.Y.; Chua, L.S.; Soontorngun, N.; Lee, C.T. Discovering potential bioactive compounds from Tualang honey. Agric. Nat. Res. 2018, 52, 361–365. [Google Scholar] [CrossRef]
- Mogana, R.; Teng-Jin, K.; Wiart, C. The Medicinal Timber Canarium patentinervium Miq. (Burseraceae Kunth.) Is an Anti-Inflammatory Bioresource of Dual Inhibitors of Cyclooxygenase (COX) and 5-Lipoxygenase (5-LOX). ISRN Biotechnol. 2013, 2013, 986361. [Google Scholar] [CrossRef] [PubMed]
- Vo, N.N.Q.; Nomura, Y.; Muranaka, T.; Fukushima, E.O. Structure–Activity Relationships of Pentacyclic Triterpenoids as Inhibitors of Cyclooxygenase and Lipoxygenase Enzymes. J. Nat. Prod. 2019, 82, 3311–3320. [Google Scholar] [CrossRef]
- Allinger, N.L. Conformational analysis. 130. MM2. A hydrocarbon force field utilizing V1 and V2 torsional terms. J. Am. Chem. Soc. 1977, 99, 8127–8134. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Gilbert, N.C.; Gerstmeier, J.; Schexnaydre, E.E.; Börner, F.; Garscha, U.; Neau, D.B.; Werz, O.; Newcomer, M.E. Structural and mechanistic insights into 5-lipoxygenase inhibition by natural products. Nat. Chem. Biol. 2020, 16, 783–790. [Google Scholar] [CrossRef]
- Eldridge, M.D.; Murray, C.W.; Auton, T.R.; Paolini, G.V.; Mee, R.P. Empirical scoring functions: I. The development of a fast empirical scoring function to estimate the binding affinity of ligands in receptor complexes. J. Comput. Aided Mol. Des. 1997, 11, 425–445. [Google Scholar] [CrossRef]
- Sayre, C.L.; Alrushaid, S.; Martinez, S.E.; Anderson, H.D.; Davies, N.M. Pre-Clinical Pharmacokinetic and Pharmacodynamic Characterization of Selected Chiral Flavonoids: Pinocembrin and Pinostrobin. J. Pharm. Pharm. Sci. 2015, 18, 368–395. [Google Scholar] [CrossRef]
- Radmark, O. Formation of eicosanoids and other oxylipins in human macrophages. Biochem. Pharmacol. 2022, 204, 115210. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.W.; Zhang, Y. DockRMSD: An open-source tool for atom mapping and RMSD calculation of symmetric molecules through graph isomorphism. J. Cheminform. 2019, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, D.; Caballero, J. Is It Reliable to Take the Molecular Docking Top Scoring Position as the Best Solution without Considering Available Structural Data? Molecules 2018, 23, 1038. [Google Scholar] [CrossRef] [PubMed]
- Jaipetch, T.; Kanghae, S.; Pancharoen, O.; Patrick, V.; Reutrakul, V.; Tuntiwachwuttikul, P.; White, A. Constituents of Boesenbergia pandurata (syn. Kaempferia pandurata): Isolation, Crystal Structure and Synthesis of (±)-Boesenbergin A. Aust. J. Chem. 1982, 35, 351–361. [Google Scholar] [CrossRef]
- Jitvaropas, R.; Saenthaweesuk, S.; Somparn, N.; Thuppia, A.; Sireeratawong, S.; Phoolcharoen, W. Antioxidant, Antimicrobial and Wound Healing Activities of Boesenbergia rotunda. Nat. Prod. Commun. 2012, 7, 1934578X1200700727. [Google Scholar] [CrossRef]
- Teethaisong, Y.; Pimchan, T.; Srisawat, R.; Hobbs, G.; Eumkeb, G. Boesenbergia rotunda (L.) Mansf. extract potentiates the antibacterial activity of some β-lactams against β-lactam-resistant staphylococci. J. Glob. Antimicrob. Resist. 2018, 12, 207–213. [Google Scholar] [CrossRef]
- Dorman, H.D.; Deans, S.G. Antimicrobial agents from plants: Antibacterial activity of plant volatile oils. J. Appl. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef]
- Huang, C.B.; George, B.; Ebersole, J.L. Antimicrobial activity of n-6, n-7 and n-9 fatty acids and their esters for oral microorganisms. Arch. Oral Biol. 2010, 55, 555–560. [Google Scholar] [CrossRef]
- Simões, M.; Rocha, S.; Coimbra, M.A.; Vieira, M.J. Enhancement of Escherichia coli and Staphylococcus aureus antibiotic susceptibility using sesquiterpenoids. Med. Chem. 2008, 4, 616–623. [Google Scholar] [CrossRef]
- Platzer, M.; Kiese, S.; Tybussek, T.; Herfellner, T.; Schneider, F.; Schweiggert-Weisz, U.; Eisner, P. Radical Scavenging Mechanisms of Phenolic Compounds: A Quantitative Structure-Property Relationship (QSPR) Study. Front. Nutr. 2022, 9, 663. [Google Scholar] [CrossRef]
- Foti, M.C. Use and Abuse of the DPPH• Radical. J. Agric. Food Chem. 2015, 63, 8765–8776. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.K.; Jaiswal, G.; Bhutani, K.K. A review on biological sources, chemistry and pharmacological activities of pinostrobin. Nat. Prod. Res. 2016, 30, 2017–2027. [Google Scholar] [CrossRef] [PubMed]
- Tanjung, M.; Tjahjandarie, T.S.; Sentosa, M.H. Antioxidant and cytotoxic agent from the rhizomes of Kaempferia pandurata. Asian Pac. J. Trop. Dis. 2013, 3, 401–404. [Google Scholar] [CrossRef]
- Hadisaputra, S.; Purwoko, A.A.; Hakim, A.; Wati, R.; Asnawati, D.; Prananto, Y.P. Experimental and Theoretical Study of Pinostrobin as Copper Corrosion Inhibitor at 1 M H2SO4 Medium. IOP Conf. Ser. Mater. Sci. Eng. 2020, 833, 012010. [Google Scholar] [CrossRef]
- Atun, S.; Handayani, S.; Frindryani, L.F. Identification and antioxidant activity test of bioactive compound produced from ethanol extract of temukunci (Boesenbergia rotunda). AIP Conf. Proc. 2017, 1868, 020007. [Google Scholar]
- Fakhrudin, N.; Pramiastuti, O.; Wahyuono, S. A Simple, Fast, and Inexpensive Nonchromatographic Method for the Isolation of Pinostrobin from Boesenbergia Rotunda Rhizome. Rasayan J. Chem. 2021, 14, 735–740. [Google Scholar] [CrossRef]
- Manda, G.; Rojo, A.I.; Martínez-Klimova, E.; Pedraza-Chaverri, J.; Cuadrado, A. Nordihydroguaiaretic Acid: From Herbal Medicine to Clinical Development for Cancer and Chronic Diseases. Front. Pharmacol. 2020, 11, 151. [Google Scholar] [CrossRef]
- Pufahl, R.A.; Kasten, T.P.; Hills, R.; Gierse, J.K.; Reitz, B.A.; Weinberg, R.A.; Masferrer, J.L. Development of a fluorescence-based enzyme assay of human 5-lipoxygenase. Anal. Biochem. 2007, 364, 204–212. [Google Scholar] [CrossRef]
- Dzoyem, J.P.; Nkuete, A.H.L.; Ngameni, B.; Eloff, J.N. Anti-inflammatory and anticholinesterase activity of six flavonoids isolated from Polygonum and Dorstenia species. Arch. Pharm. Res. 2017, 40, 1129–1134. [Google Scholar] [CrossRef]
- Roman, W.A.; Gomes, D.B.; Zanchet, B.; Schönell, A.P.; Diel, K.A.P.; Banzato, T.P.; Ruiz, A.L.T.G.; Carvalho, J.E.; Neppel, A.; Barison, A.; et al. Antiproliferative effects of pinostrobin and 5,6-dehydrokavain isolated from leaves of Alpinia zerumbet. Rev. Bras. Farmacogn. 2017, 27, 592–598. [Google Scholar]
- González, A.S.; Soto Tellini, V.H.; Benjumea Gutiérrez, D.M. Study of the dermal anti-inflammatory, antioxidant, and analgesic activity of pinostrobin. Heliyon 2022, 8, e10413. [Google Scholar] [CrossRef] [PubMed]
- Sivaranjani, N.; Rao, S.V.; Rajeev, G. Role of reactive oxygen species and antioxidants in atopic dermatitis. J. Clin. Diagn. Res. 2013, 7, 2683–2685. [Google Scholar] [CrossRef] [PubMed]
- Bertino, L.; Guarneri, F.; Cannavò, S.P.; Casciaro, M.; Pioggia, G.; Gangemi, S. Oxidative Stress and Atopic Dermatitis. Antioxidants 2020, 9, 196. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.M.; Kang, M.C.; Jin, M.; Lee, T.H.; Lim, B.O.; Kim, S.Y. Fermented blueberry and black rice containing Lactobacillus plantarum MG4221: A novel functional food for particulate matter (PM2.5)/dinitrochlorobenzene (DNCB)-induced atopic dermatitis. Food Funct. 2021, 12, 3611–3623. [Google Scholar] [CrossRef]
- Sanabria-de la Torre, R.; Fernández-González, A.; Quiñones-Vico, M.I.; Montero-Vilchez, T.; Arias-Santiago, S. Bioengineered Skin Intended as In Vitro Model for Pharmacosmetics, Skin Disease Study and Environmental Skin Impact Analysis. Biomedicines 2020, 8, 464. [Google Scholar] [CrossRef]
- Jang, H.J.; Lee, J.B.; Yoon, J.K. Advanced In Vitro Three-Dimensional Skin Models of Atopic Dermatitis. Tissue Eng. Regen. Med. 2023, 20, 539–552. [Google Scholar] [CrossRef]
- Seo, H.-S.; Seong, K.H.; Kim, C.-D.; Seo, S.J.; Park, B.C.; Kim, M.H.; Hong, S.-P. Adiponectin Attenuates the Inflammation in Atopic Dermatitis-Like Reconstructed Human Epidermis. Ann. Dermatol. 2019, 31, 186–195. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Species | Part Used | Zone of Inhibition (mm) | MIC (µg/mL) | ||
|---|---|---|---|---|---|---|
| Essential Oils (c = 1000 µg/Disk) | Hexane Extract (c = 250 µg/Disk) | Essential Oils | Hexane Extract | |||
| 1 | A. vulgaris | Aerial part | - | - | nd | nd |
| 2 | B. rotunda | Rhizome | 13 | 9 | 100 | 10 |
| 3 | C. sinensis | Young leaves | 12 | - | nd | nd |
| 4 | C. fistula | Leaves | nd | - | nd | nd |
| Pods | nd | - | nd | nd | ||
| 5 | D. longan | Peels | - | - | nd | nd |
| Seeds | nd | - | nd | nd | ||
| 6 | M. villosa | Leaves | - | - | nd | nd |
| 7 | W. trilobata | Leaves | 9 | - | nd | nd |
| Pinostrobin | nd | >200 | ||||
| Tetracycline | nd | 3.125 | ||||
| No. | Species | Part Used | DPPH Radical; IC50 (µg/mL) | NO Radical; IC50 (µg/mL) | FRAP (mM FeSO4/g Extract) | |||
|---|---|---|---|---|---|---|---|---|
| EOs | Hexane Extract | EOs | Hexane Extract | EOs | Hexane Extract | |||
| 1 | A. vulgaris | Aerial part | - | - | - | - | 0.37 ± 0.05 ab | 0.36 ± 0.14 ab |
| 2 | B. rotunda | Rhizome | 4085.67 ± 545.13 d | 557.97 ± 27.86 a | 1695.00 ± 128.18 b | 2651.67 ± 138.68 d | 0.24 ± 0.08 a | 2.26 ± 0.17 h |
| 3 | C. sinensis | Young leaves | - | 208.90 ± 4.81 a | - | 1809.67 ± 91.22 bc | 0.51 ± 0.09 bc | 3.87 ± 0.16 j |
| 4 | C. fistula | Leaves | nd | 3993.67 ± 175.25 d | nd | 4620.33 ± 285.35 e | nd | 2.61 ± 0.20 i |
| Pods | nd | - | nd | - | nd | 1.12 ± 0.13 f | ||
| 5 | D. longan | Peels | - | 1123.00 ± 115.21 b | - | 675.70 ± 54.37 a | 1.09 ± 0.11 ef | 1.13 ± 0.14 f |
| Seeds | nd | - | nd | - | nd | 0.97 ± 0.08 def | ||
| 6 | M. villosa | Aerial part | - | 2885.33 ± 280.08 c | - | - | 0.99 ± 0.07 def | 1.41 ± 0.27 g |
| 7 | W. trilobata | Leaves | - | - | 2849.67 ± 82.80 d | 2434.33 ± 984.08 cd | 0.84 ± 0.09 de | 0.73 ± 0.22 cd |
| Quercetin | 19.08 ± 1.34 (63.13 ± 4.43 µM) | 93.58 ± 7.93 (309.62 ± 26.24 µM) | - | - | ||||
| Pinostrobin | >1000 | >1000 | 1.04 ± 0.01 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liana, D.; Eurtivong, C.; Phanumartwiwath, A. Boesenbergia rotunda and Its Pinostrobin for Atopic Dermatitis: Dual 5-Lipoxygenase and Cyclooxygenase-2 Inhibitor and Its Mechanistic Study through Steady-State Kinetics and Molecular Modeling. Antioxidants 2024, 13, 74. https://doi.org/10.3390/antiox13010074
Liana D, Eurtivong C, Phanumartwiwath A. Boesenbergia rotunda and Its Pinostrobin for Atopic Dermatitis: Dual 5-Lipoxygenase and Cyclooxygenase-2 Inhibitor and Its Mechanistic Study through Steady-State Kinetics and Molecular Modeling. Antioxidants. 2024; 13(1):74. https://doi.org/10.3390/antiox13010074
Chicago/Turabian StyleLiana, Desy, Chatchakorn Eurtivong, and Anuchit Phanumartwiwath. 2024. "Boesenbergia rotunda and Its Pinostrobin for Atopic Dermatitis: Dual 5-Lipoxygenase and Cyclooxygenase-2 Inhibitor and Its Mechanistic Study through Steady-State Kinetics and Molecular Modeling" Antioxidants 13, no. 1: 74. https://doi.org/10.3390/antiox13010074
APA StyleLiana, D., Eurtivong, C., & Phanumartwiwath, A. (2024). Boesenbergia rotunda and Its Pinostrobin for Atopic Dermatitis: Dual 5-Lipoxygenase and Cyclooxygenase-2 Inhibitor and Its Mechanistic Study through Steady-State Kinetics and Molecular Modeling. Antioxidants, 13(1), 74. https://doi.org/10.3390/antiox13010074