

Redox State Modulatory Activity and Cytotoxicity of Olea europaea L. (Oleaceae) Leaves Extract Enriched in Polyphenols Using Macroporous Resin

 ,
,  , ,
, ,  ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of Olive Leaf Enriched Extract (OLEE)
2.3. Phytochemical Analysis
2.3.1. Spectrophotometric Determinations of Total Polyphenols and Total Flavonoids
2.3.2. HPLC-DAD and HPLC-ESI-MS Analyses
2.4. Antioxidant-Free Cell Assay
DPPH Test
2.5. Cell Lines
2.5.1. Cell Viability Assay
2.5.2. Light Microscopy and Morphological Analysis
2.5.3. LDH Release
2.6. Intracellular Redox State Evaluation
2.6.1. ROS Determination
2.6.2. RSH Determination
2.7. Statistical Analysis
3. Results
3.1. Phytochemical Analysis
3.1.1. Determination of Polyphenolic Profile
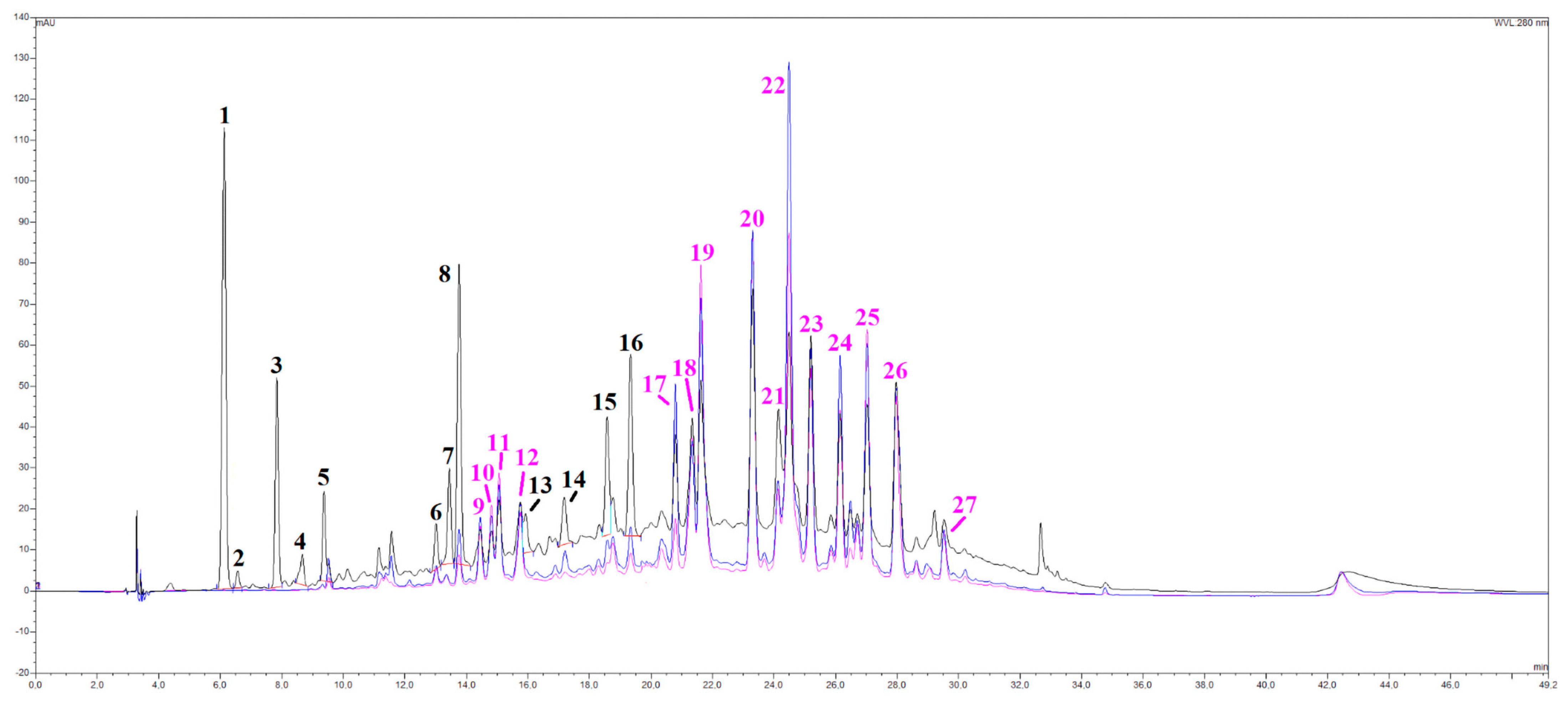
3.1.2. Compositional Analyses on OLEE
3.2. In Vitro Cell-Free Antioxidant Properties
DPPH Assay
3.3. Cytotoxicity on Normal and Cancer Cells
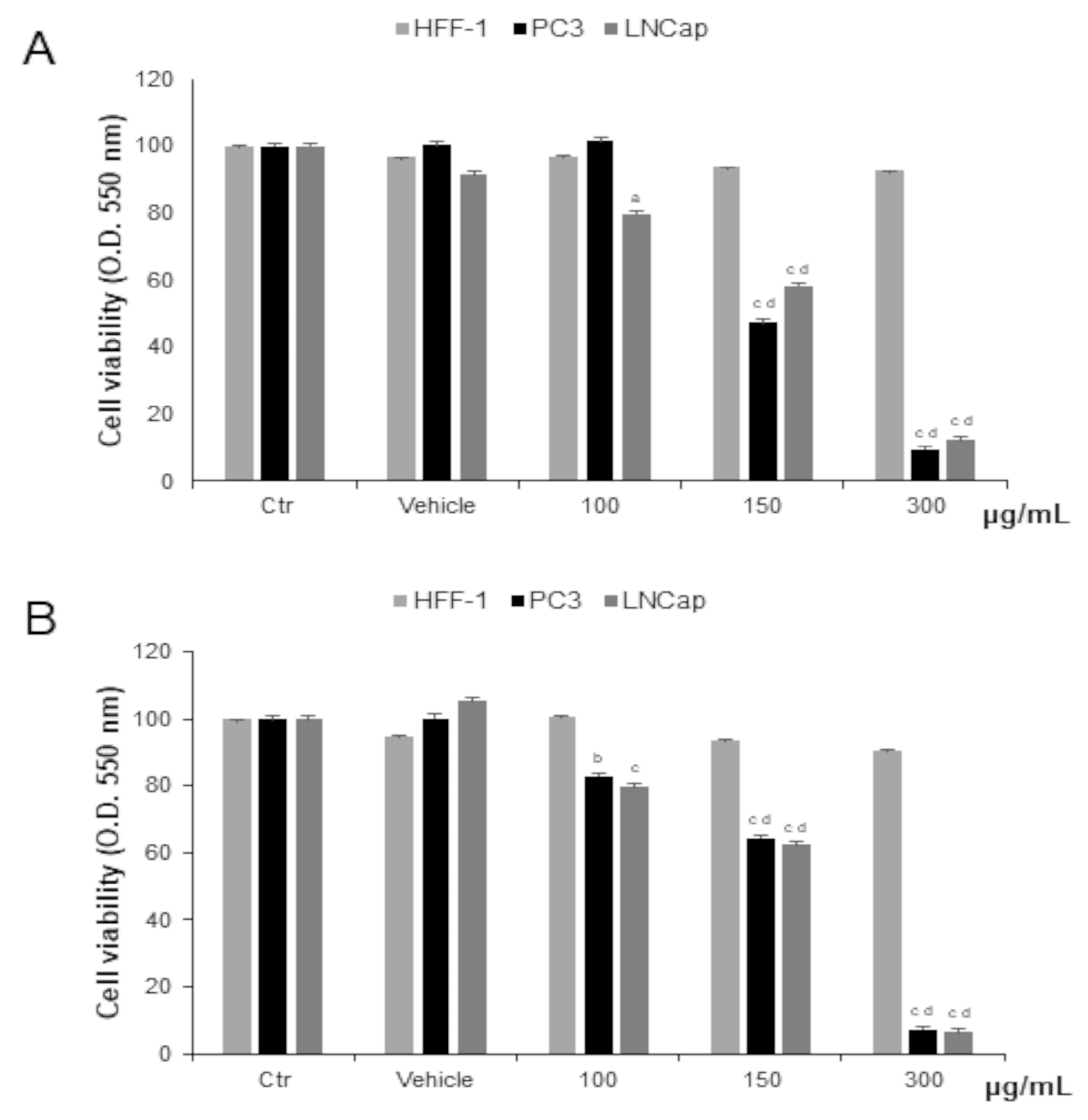
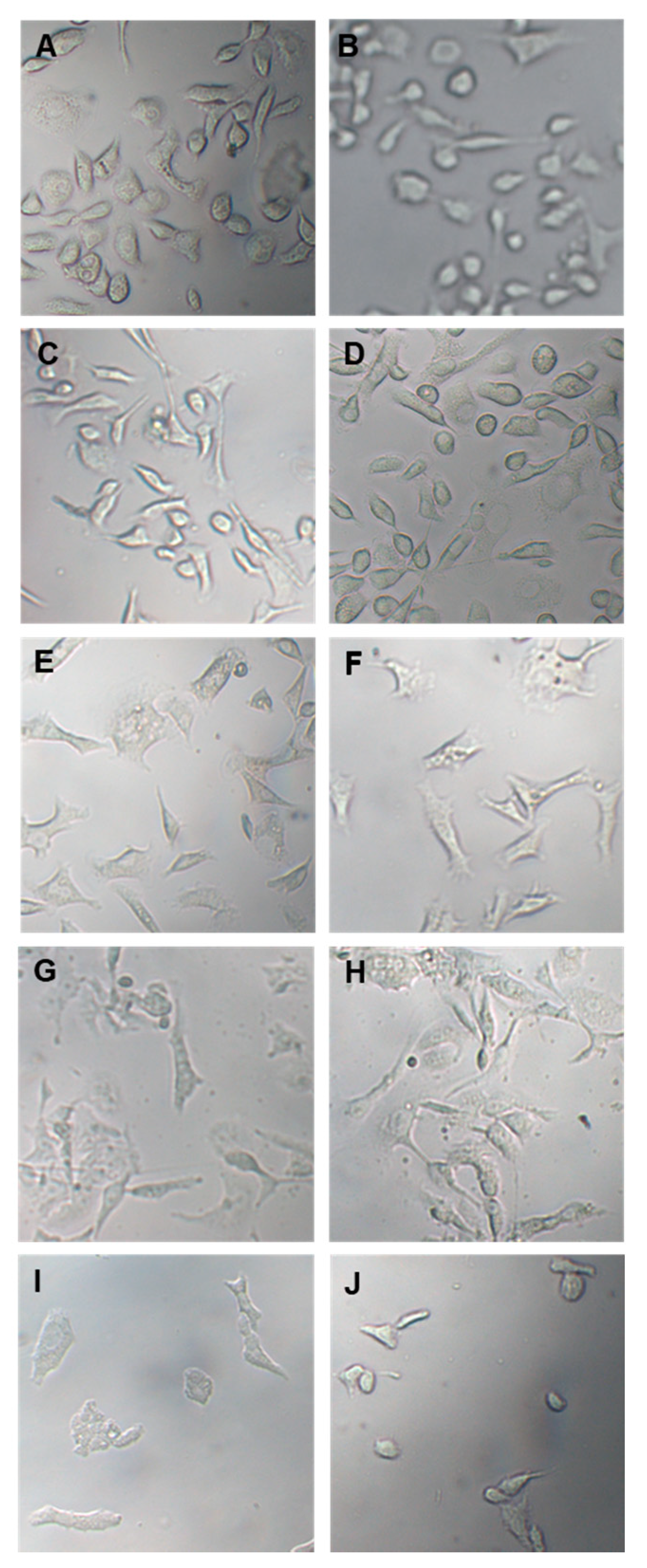
3.3.1. Effect of OLEE on Cell Viability and Morphology
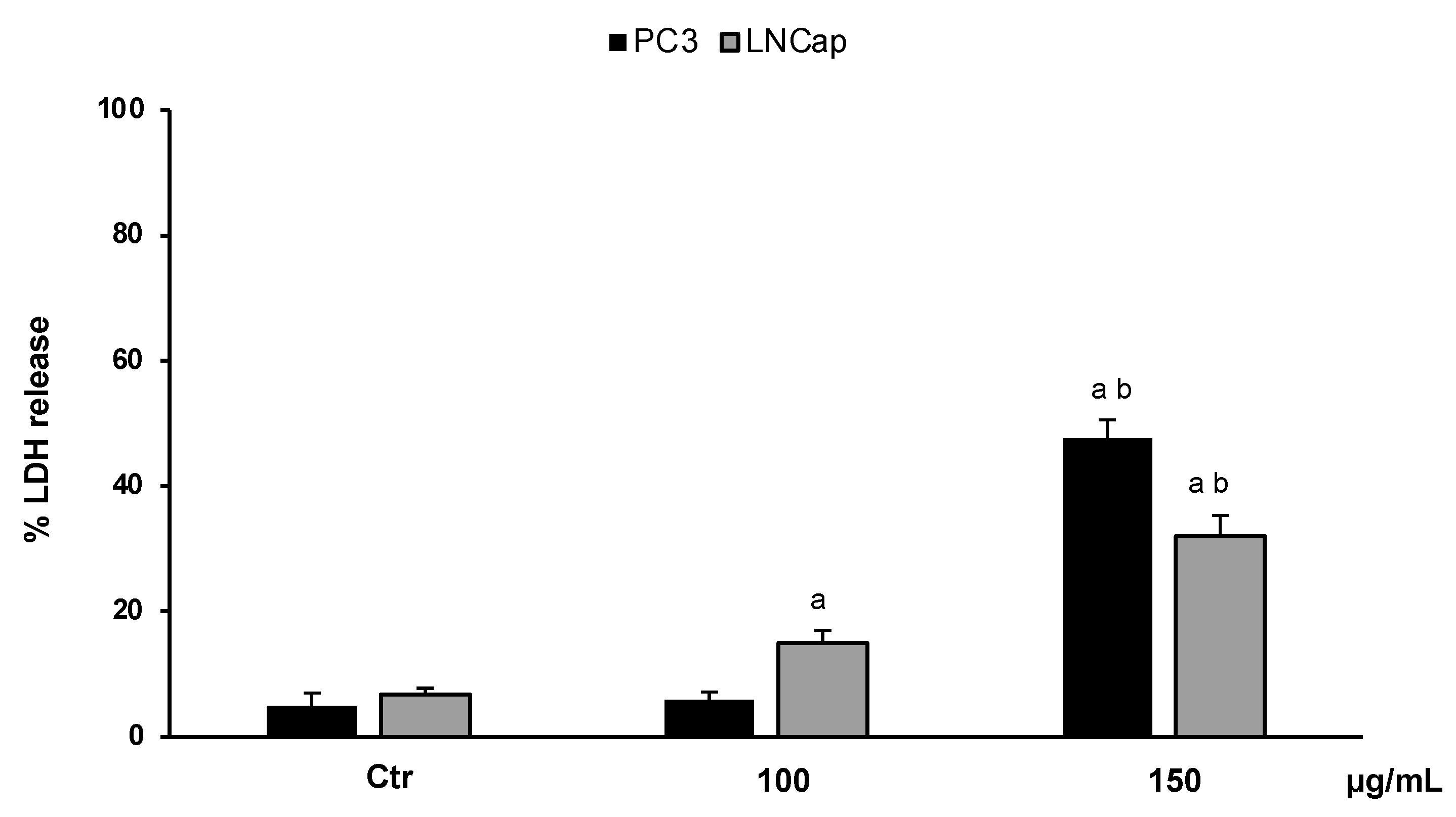
3.3.2. LDH Release
3.4. Intracellular Redox State Evaluation
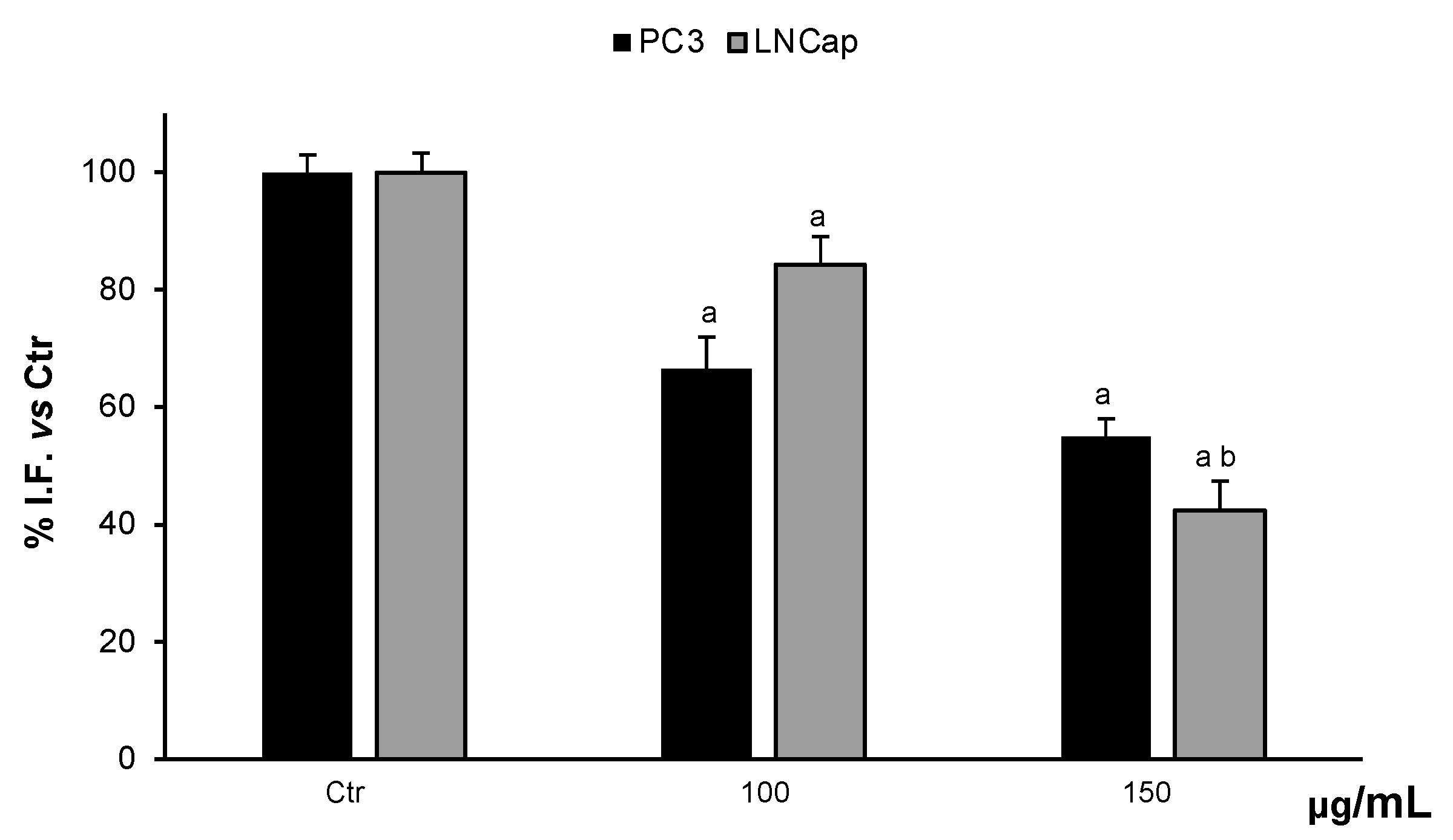
3.4.1. Determination of ROS Levels
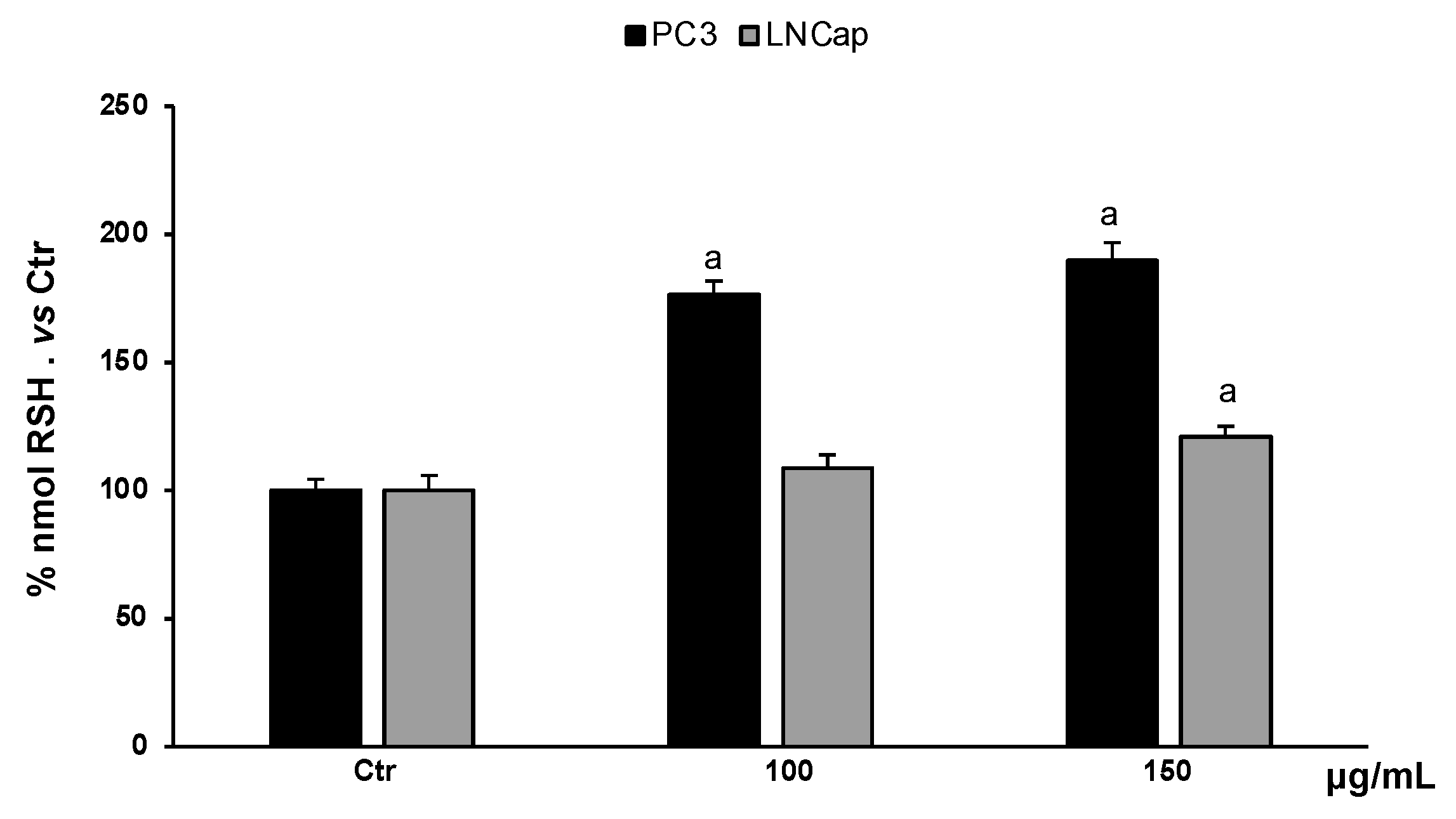
3.4.2. Determination of RSH Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mentella, M.C.; Scaldaferri, F.; Ricci, C.; Gasbarrini, A.; Miggiano, G.A.D. Cancer and Mediterranean Diet: A Review. Nutrients. 2019, 11, 2059. [Google Scholar] [CrossRef]
- Farinetti, A.; Zurlo, V.; Manenti, A.; Coppi, F.; Mattioli, A.V. Mediterranean diet and colorectal cancer: A systematic review. Nutrition 2017, 43, 83–88. [Google Scholar] [CrossRef]
- Gorzynik-Debicka, M.; Przychodzen, P.; Cappello, F.; Kuban-Jankowska, A.; Marino Gammazza, A.; Knap, N.; Wozniak, M.; Gorska-Ponikowska, M. Potential Health Benefits of Olive Oil and Plant Polyphenols. Int. J. Mol. Sci. 2018, 19, 686. [Google Scholar] [CrossRef] [PubMed]
- Cumaoğlu, A.; Ari, N.; Kartal, M.; Karasu, Ç. Polyphenolic extracts from Olea europea L. protect against cytokine-induced β-cell damage through maintenance of redox homeostasis. Rejuv. Res. 2011, 14, 325–334. [Google Scholar] [CrossRef] [PubMed]
- ALHaithloul, H.A.S.; Alotaibi, M.F.; Bin-Jumah, M.; Elgebaly, H.; Mahmoud, A.M. Olea europaea leaf extract up-regulates Nrf2/ARE/HO-1 signaling and attenuates cyclophosphamide-induced oxidative stress, inflammation and apoptosis in rat kidney. Biomed. Pharmacother. 2019, 111, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Hashmi, M.A.; Khan, A.; Hanif, M.; Farooq, U.; Perveen, S. Traditional Uses, Phytochemistry, and Pharmacology of Olea europaea (Olive). Evid. Based Complement. Alternat. Med. 2015, 2015, 541591. [Google Scholar] [CrossRef] [PubMed]
- Baci, D.; Gallazzi, M.; Cascini, C.; Tramacere, M.; De Stefano, D.; Bruno, A.; Noonan, D.M.; Albini, A. Downregulation of Pro-Inflammatory and Pro-Angiogenic Pathways in Prostate Cancer Cells by a Polyphenol-Rich Extract from Olive Mill Wastewater. Int. J. Mol. Sci. 2019, 20, 307. [Google Scholar] [CrossRef] [PubMed]
- Acquaviva, R.; Di Giacomo, C.; Sorrenti, V.; Galvano, F.; Santangelo, R.; Cardile, V.; Gangia, S.; D’Orazio, N.; Abraham, N.G.; Vanella, L. Antiproliferative effect of oleuropein in prostate cell lines. Int. J. Oncol. 2012, 41, 31–38. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Center, M.M.; Jemal, A.; Lortet-Tieulent, J.; Ward, E.; Ferlay, J.; Brawley, O.; Bray, F. International variation in prostate cancer incidence and mortality rates. Eur. Urol. 2012, 61, 1079–1092. [Google Scholar] [CrossRef]
- Rebbeck, T.R.; Devesa, S.S.; Chang, B.L.; Bunker, C.H.; Cheng, I.; Cooney, K.; Eeles, R.; Fernandez, P.; Giri, V.N.; Gueye, S.M.; et al. Global patterns of prostate cancer incidence, aggressiveness, and mortality in men of African descent. Prostate Cancer. 2013, 2013, 560857. [Google Scholar] [CrossRef] [PubMed]
- Vietri, M.T.; D’Elia, G.; Caliendo, G.; Resse, M.; Casamassimi, A.; Passariello, L.; Albanese, L.; Cioffi, M.; Molinari, A.M. Hereditary Prostate Cancer: Genes Related, Target Therapy and Prevention. Int. J. Mol. Sci. 2021, 22, 3753. [Google Scholar] [CrossRef] [PubMed]
- Allott, E.H.; Masko, E.M.; Freedland, S.J. Obesity and prostate cancer: Weighing the evidence. Eur. Urol. 2013, 63, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Xu, Z.; Xu, T.; Yu, B.; Zou, Q. Diabetes mellitus is associated with elevated risk of mortality amongst patients with prostate cancer: A metaanalysis of 11 cohort studies. Diabetes Metab. Res Rev. 2015, 31, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Peisch, S.F.; Van Blarigan, E.L.; Chan, J.M.; Stampfer, M.J.; Kenfield, S.A. Prostate cancer progression and mortality: A review of diet and lifestyle factors. World J. Urol. 2017, 35, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Tikkinen, K.A.O.; Dahm, P.; Lytvyn, L.; Heen, A.F.; Vernooij, R.W.M.; Siemieniuk, R.A.C.; Wheeler, R.; Vaughan, B.; Fobuzi, A.C.; Blanker, M.H.; et al. Prostate cancer screening with prostate-specific antigen (PSA) test: A clinical practice guideline. BMJ. 2018, 362, k3581. [Google Scholar] [CrossRef] [PubMed]
- Zargar, H.; van den Bergh, R.; Moon, D.; Lawrentschuk, N.; Costello, A.; Murphy, D. The impact of the United States Preventive Services Task Force (USPTSTF) recommendations against prostate-specific antigen (PSA) testing on PSA testing in Australia. BJU Int. 2017, 119, 110–115. [Google Scholar] [CrossRef]
- Culp, M.B.; Soerjomataram, I.; Efstathiou, J.A.; Bray, F.; Jemal, A. Recent global patterns in prostate cancer incidence and mortality rates. Eur. Urol. 2020, 77, 38–52. [Google Scholar] [CrossRef]
- Chen, R.C.; Rumble, R.B.; Loblaw, D.A.; Finelli, A.; Ehdaie, B.; Cooperberg, M.R.; Morgan, S.C.; Tyldesley, S.; Haluschak, J.J.; Tan, W.; et al. Active surveillance for the management of localized prostate cancer (Cancer Care Ontario Guideline): American Society of Clinical Oncology Clinical Practice Guideline Endorsement. J. Clin. Oncol. 2016, 34, 2182–2190. [Google Scholar] [CrossRef]
- Gravis, G.; Audenet, F.; Irani, J.; Timsit, M.O.; Barthelemy, P.; Beuzeboc, P.; Fléchon, A.; Linassier, C.; Oudard, S.; Rebillard, X.; et al. Chemotherapy in hormone-sensitive metastatic prostate cancer: Evidences and uncertainties from the literature. Cancer Treat. Rev. 2017, 55, 211–217. [Google Scholar] [CrossRef]
- Lorente, D.; Fizazi, K.; Sweeney, C.; de Bono, J.S. Optimal treatment sequence for metastatic castration-resistant prostate cancer. Eur. Urol. Focus 2016, 2, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Yeku, O.; Slovin, S.F. Immune therapy for prostate cancer. Cancer J. 2016, 22, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Yacoubian, A.; Dargham, R.A.; Khauli, R.B.; Bachir, B.G. Overview of dietary supplements in prostate cancer. Curr. Urol. Rep. 2016, 17, 78. [Google Scholar] [CrossRef] [PubMed]
- Acquaviva, R.; D’Angeli, F.; Malfa, G.A.; Ronsisvalle, S.; Garozzo, A.; Stivala, A.; Ragusa, S.; Nicolosi, D.; Salmeri, M.; Genovese, C. Antibacterial and anti-biofilm activities of walnut pellicle extract (Juglans regia L.) against coagulase-negative staphylococci. Nat. Prod. Res. 2021, 35, 2076–2081. [Google Scholar] [CrossRef] [PubMed]
- Gambacorta, G.; Faccia, M.; Previtali, M.A.; Pati, S.; La Notte, E.; Baino, A. Effect of olive maturation and stoning on quality indices and antioxidant content of extra virgin olive oil (cv. Coratina) during storage. J. Food Sci. 2010, 75, C229–C235. [Google Scholar] [CrossRef] [PubMed]
- Napoli, E.; Siracusa, L.; Ruberto, G.; Carrubba, A.; Lazzara, S.; Speciale, A.; Cimino, F.; Saija, A.; Cristani, M. Phytochemical profiles, phototoxic and antioxidant properties of eleven Hypericum species—A comparative study. Phytochemistry 2018, 152, 162–173. [Google Scholar] [CrossRef]
- Campisi, A.; Acquaviva, R.; Raciti, G.; Duro, A.; Rizzo, M.; Santagati, N.A. Antioxidant Activities of Solanum nigrum L. Leaf Extracts Determined in in vitro Cellular Models. Foods 2019, 8, 63. [Google Scholar] [CrossRef]
- Luca, T.; Napoli, E.; Privitera, G.; Musso, N.; Ruberto, G.; Castorina, S. Antiproliferative Effect and Cell Cycle Alterations Induced by Salvia officinalis Essential Oil and Its Three Main Components in Human Colon Cancer Cell Lines. Chem. Biodiver. 2020, 2017, e2000309. [Google Scholar] [CrossRef]
- Taviano, M.F.; Miceli, N.; Acquaviva, R.; Malfa, G.A.; Ragusa, S.; Giordano, D.; Cásedas, G.; Les, F.; López, V. Cytotoxic, Antioxidant, and Enzyme Inhibitory Properties of the Traditional Medicinal Plant Matthiola incana (L.) R. Br. Biology 2020, 9, 163. [Google Scholar] [CrossRef]
- Di Giacomo, C.; Acquaviva, R.; Piva, A.; Sorrenti, V.; Vanella, L.; Piva, G.; Casadei, G.; La Fauci, L.; Ritieni, A.; Bognanno, M.; et al. Protective effect of cyanidin 3-O-beta-D-glucoside on ochratoxin A-mediated damage in the rat. Br. J. Nutr. 2007, 98, 937–943. [Google Scholar] [CrossRef]
- Román, G.C.; Jackson, R.E.; Gadhia, R.; Román, A.N.; Reis, J. Mediterranean diet: The role of long-chain ω-3 fatty acids in fish; polyphenols in fruits, vegetables, cereals, coffee, tea, cacao and wine; probiotics and vitamins in prevention of stroke, age-related cognitive decline, and Alzheimer disease. Rev. Neurol. 2019, 175, 724–741. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.G.; Allkanjari, O.; Busetto, G.M.; Cai, T.; Larganà, G.; Magri, V.; Perletti, G.; Robustelli Della Cuna, F.S.; Russo, G.I.; Stamatiou, K.; et al. Nutraceutical treatment and prevention of benign prostatic hyperplasia and prostate cancer. Arch Ital Urol Androl. 2019, 91. [Google Scholar] [CrossRef] [PubMed]
- Leri, M.; Scuto, M.; Ontario, M.L.; Calabrese, V.; Calabrese, E.J.; Bucciantini, M.; Stefani, M. Healthy Effects of Plant Polyphenols: Molecular Mechanisms. Int. J. Mol. Sci. 2020, 21, 1250. [Google Scholar] [CrossRef] [PubMed]
- El, S.N.; Karakaya, S. Olive tree (Olea europaea) leaves: Potential beneficial effects on human health. Nutr. Rev. 2009, 67, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Papoti, V.T.; Papageorgiou, M.; Dervisi, K.; Alexopoulos, E.; Apostolidis, K.; Petridis, D. Screening Olive Leaves from Unexploited Traditional Greek Cultivars for Their Phenolic Antioxidant Dynamic. Foods 2018, 7, 197. [Google Scholar] [CrossRef] [PubMed]
- Lins, P.G.; Marina Piccoli Pugine, S.; Scatolini, A.M.; de Melo, M.P. In vitro antioxidant activity of olive leaf extract (Olea europaea L.) and its protective effect on oxidative damage in human erythrocytes. Heliyon 2018, 4, e00805. [Google Scholar] [CrossRef] [PubMed]
- Aresta, M.; Dibenedetto, A.; Narracci, M.; Tommasi, I. A technology for the treatment of olive-mill waste water in a continuously fed plant. Environ. Chem. Lett. 2003, 1, 13–18. [Google Scholar] [CrossRef]
- Wang, T.Y.; Li, Q.; Bi, K.S. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef]
- Deng, D.; Zhang, J.; Cooney, J.M.; Skinner, M.A.; Adaim, A.; Jensen, D.J.; Stevenson, D.E. Methylated polyphenols are poor “chemical” antioxidants but can still effectively protect cells from hydrogen peroxide-induced cytotoxicity. FEBS Lett. 2006, 580, 5247–5250. [Google Scholar] [CrossRef]
- Mansour, H.M.M.; Zeitoun, A.A.; Abd-Rabou, H.S.; El Enshasy, H.A.; Dailin, D.J.; Zeitoun, M.A.A.; El-Sohaimy, S.A. Antioxidant and Anti-Diabetic Properties of Olive (Olea europaea) Leaf Extracts: In Vitro and In Vivo Evaluation. Antioxidants 2023, 12, 1275. [Google Scholar] [CrossRef]
- Makowska-Wąs, J.; Galanty, A.; Gdula-Argasińska, J.; Tyszka-Czochara, M.; Szewczyk, A.; Nunes, R.; Carvalho, I.S.; Michalik, M.; Paśko, P. Identification of Predominant Phytochemical Compounds and Cytotoxic Activity of Wild Olive Leaves (Olea europaea L. ssp. sylvestris) Harvested in South Portugal. Chem. Biodivers. 2017, 14, e1600331. [Google Scholar] [CrossRef] [PubMed]
- Albogami, S.; Hassan, A.M. Assessment of the Efficacy of Olive Leaf (Olea europaea L.) Extracts in the Treatment of Colorectal Cancer and Prostate Cancer Using In Vitro Cell Models. Molecules 2021, 26, 4069. [Google Scholar] [CrossRef] [PubMed]
- Freitas, D.D.S.; Morgado-Díaz, J.A.; Gehren, A.S.; Vidal, F.C.B.; Fernandes, R.M.T.; Romão, W.; Tose, L.V.; Frazão, F.N.S.; Costa, M.C.P.; Silva, D.F.; et al. Cytotoxic analysis and chemical characterization of fractions of the hydroalcoholic extract of the Euterpe oleracea Mart. seed in the MCF-7 cell line. J. Pharm. Pharmacol. 2017, 69, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Tongyi, S. Role of Bax/Bcl-2 family members in green tea polyphenol induced necroptosis of p53-deficient Hep3B cells. Tumour Biol. 2014, 35, 8065–8075. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.G.; Jiao, D. Necroptosis, tumor necrosis and tumorigenesis. Cell Stress 2019, 4, 1–8. [Google Scholar] [CrossRef]
- Gupta-Elera, G.; Garrett, A.R.; Robison, R.A.; O’Neill, K.L. The role of oxidative stress in prostate cancer. Eur. J. Cancer Prev. 2012, 21, 155–162. [Google Scholar] [CrossRef]
- Chowdhury, S.K.; Gemin, A.; Singh, G. High activity of mitochondrial glycerophosphate dehydrogenase and glycerophosphate-dependent ROS production in prostate cancer cell lines. Biochem. Biophys. Res. Commun. 2005, 333, 1139–1145. [Google Scholar] [CrossRef]
- Chaiswing, L.; Bourdeau-Heller, J.M.; Zhong, W.; Oberley, T.D. Characterization of redox state of two human prostate carcinoma cell lines with different degrees of aggressiveness. Free Radic. Biol. Med. 2007, 43, 202–215. [Google Scholar] [CrossRef]
- Kumar, B.; Koul, S.; Khandrika, L.; Meacham, R.B.; Koul, H.K. Oxidative stress is inherent in prostate cancer cells and is required for aggressive phenotype. Cancer Res. 2008, 68, 1777–1785. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.W.; Hong, S.; Jin, W.; Lim, S.; Kim, S.J.; Kang, H.J.; Park, E.H.; Ahn, K.; Lim, C.J. Up-regulation of defense enzymes is responsible for low reactive oxygen species in malignant prostate cancer cells. Exp. Mol. Med. 2005, 37, 497–506. [Google Scholar] [CrossRef]
- Li, L.; Jin, P.; Guan, Y.; Luo, M.; Wang, Y.; He, B.; Li, B.; He, K.; Cao, J.; Huang, C.; et al. Exploiting Polyphenol-Mediated Redox Reorientation in Cancer Therapy. Pharmaceuticals 2022, 15, 1540. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Polyphenols (mg GAE/g Extract) | Total Flavonoids (mg CE/g Extract) | DPPH Test IC50 (μg/mL) | |
|---|---|---|---|
| OLEE | 130.02 ± 2.3 | 70.13 ± 1.2 | 100.00 ± 1.8 |
| Trolox | 15 µM ± 0.62 |
| Peak # | Rt, Min a | Compound Tentative Identification | UV-Vis Data, nm b | ESI- Data, m/z c |
|---|---|---|---|---|
| 1 | 6.1 | hydroxytyrosol hexoside | 280.2 | 315.0952 * (M-H); 153.0987 |
| 2 | 6.6 | 3,4 dihydroxyphenylacetic acid (DOPAC) d | 280.6 | 167.0419 (M-H); 190.0309 * (M-H+Na) |
| 3 | 7.8 | Hydroxytyrosol d | 275.9 | 153.0627 (M-H) |
| 4 | 8.7 | hydroxytyrosol derivative | 279.1 | 315.0595 (M-H) (tr) |
| 5 | 9.4 | DOPAC derivative | 280.4 | 315.0462; 181.0391 * (M-H) |
| 6 | 13.0 | eriodictyol hexoside-deoxyhexoside | 283; 336 | 595.1698 * (M-H); 449.0234 (M-H-deoxyhex) |
| 7 | 13.4 | hydroxytyrosol derivative 2 | 280.1 | 315.0603 * (M-H) |
| 8 | 13.8 | naringenin hexoside-deoxyhexoside | 284; 332sh | 579.1677 * (M-H); 433.1583(M-H-deoxyhex) |
| 9 | 14.5 | apigenin hexoside derivative | 266; 336 | 611.1039 * (M-H) |
| 10 | 14.8 | luteolin di-hexoside | 257; 265; 346 | 609.1529 *(M-H); 447.1531 (M-H-hex) |
| 11 | 15.1 | luteolin hexoside-pentoside | 253.4; 266.2; 345 | 579.1419 * (M-H) |
| 12 | 15.8 | methylapigenin di-hexoside | 268; 337 | 607.1747 *(M-H); 445.1672 (M-H-hex) |
| 13 | 15.9 | ligstroside d | 288.2 | 523.1778 (M-H); 361.1532 (M-H-glc) |
| 14 | 18.0 | oleuropein aglycone | 287 | 377.1309 (M-H) |
| 15 | 18.6 | eriodictyol deoxyhexoside | 289; 335sh | 433.1039 * (M-H) |
| 16 | 19.3 | naringenin deoxyhexoside | 287; 332sh | 417.1275 * (M-H) |
| 17 | 20.8 | luteolin 7-O-glucoside d | 248; 267;311 | 447.0097 * (M-H); 470.0989 (M-H+Na) |
| 18 | 21.3 | methylapigenin hexoside | 272; 337 | 445.1209 * (M-H) |
| 19 | 21.6 | methylluteolin hexoside | 249; 266; 345 | 461.1158 * (M-H) |
| 20 | 23.3 | luteolin deoxyhexoside | 248; 269; 341 | 431.1049 * (M-H) |
| 21 | 24.1 | methylapigenin deoxyhexoside | 274; 338 | 429.1258 * (M-H) |
| 22 | 24.5 | methylluteolin isomer 1 | 241; 263sh; 330 | 599.0658 * (2M-H) |
| 23 | 25.2 | methylapigenin isomer 1 | 270; 337 | 283.0679 * (M-H) |
| 24 | 26.1 | methylluteolin isomer 2 | 249; 269; 333 | 299.0629 (M-H); 599.0657 * (2M-H) |
| 25 | 27.0 | methylluteolin isomer 3 | 245; 269; 343 | 299.0621 * (M-H) |
| 26 | 28.0 | methylapigenin isomer 2 | 273; 337 | 590.1941 * (2M-H+Na); 283.0678 * |
| 27 | 29.5 | di-methylluteolin isomer 1 | 249; 268; 343 | 313.0773 (M-H); 627.1066 (2M-H) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luca, T.; Malfa, G.A.; Siracusa, L.; La Mantia, A.; Bianchi, S.; Napoli, E.; Puleo, S.; Sergi, A.; Acquaviva, R.; Castorina, S. Redox State Modulatory Activity and Cytotoxicity of Olea europaea L. (Oleaceae) Leaves Extract Enriched in Polyphenols Using Macroporous Resin. Antioxidants 2024, 13, 73. https://doi.org/10.3390/antiox13010073
Luca T, Malfa GA, Siracusa L, La Mantia A, Bianchi S, Napoli E, Puleo S, Sergi A, Acquaviva R, Castorina S. Redox State Modulatory Activity and Cytotoxicity of Olea europaea L. (Oleaceae) Leaves Extract Enriched in Polyphenols Using Macroporous Resin. Antioxidants. 2024; 13(1):73. https://doi.org/10.3390/antiox13010073
Chicago/Turabian StyleLuca, Tonia, Giuseppe Antonio Malfa, Laura Siracusa, Alfonsina La Mantia, Simone Bianchi, Edoardo Napoli, Stefano Puleo, Angelo Sergi, Rosaria Acquaviva, and Sergio Castorina. 2024. "Redox State Modulatory Activity and Cytotoxicity of Olea europaea L. (Oleaceae) Leaves Extract Enriched in Polyphenols Using Macroporous Resin" Antioxidants 13, no. 1: 73. https://doi.org/10.3390/antiox13010073
APA StyleLuca, T., Malfa, G. A., Siracusa, L., La Mantia, A., Bianchi, S., Napoli, E., Puleo, S., Sergi, A., Acquaviva, R., & Castorina, S. (2024). Redox State Modulatory Activity and Cytotoxicity of Olea europaea L. (Oleaceae) Leaves Extract Enriched in Polyphenols Using Macroporous Resin. Antioxidants, 13(1), 73. https://doi.org/10.3390/antiox13010073