
Glutathione and a Pool of Metabolites Partly Related to Oxidative Stress Are Associated with Low and High Myopia in an Altered Bioenergetic Environment


 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Groups and Patient Selection
2.2. Clinical Exploration
2.3. Aqueous Humor Sample Recollection
2.4. NMR Assay
2.5. Glutathione Assay
2.6. Statistical Analysis
3. Results
3.1. The Clinical Data Recorded Were Very Consistent
3.2. PLS-DA Confirmed the Significant Differences between the Control and High-Myopia Groups
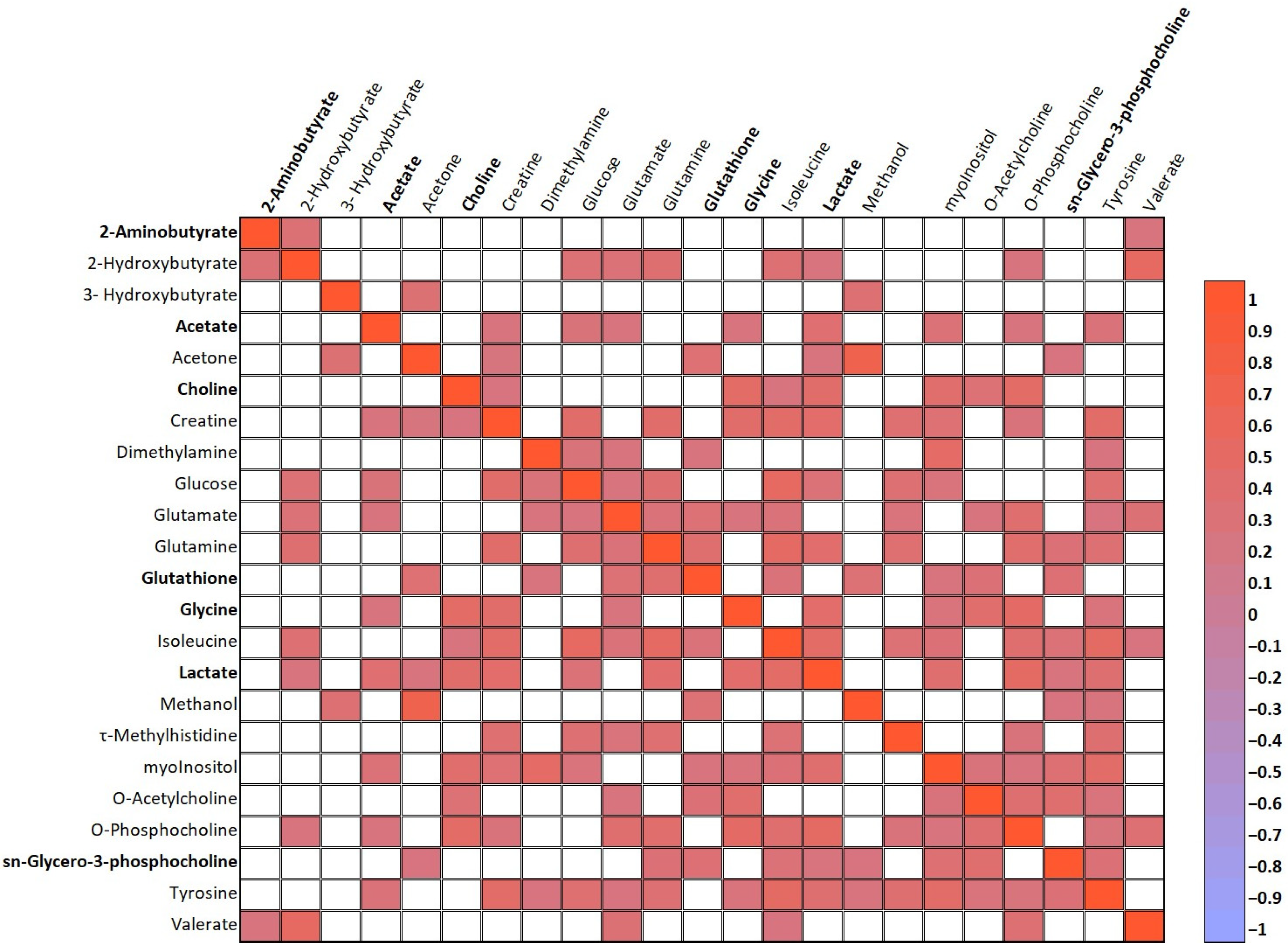
3.3. Main Metabolite Differences in the Aqueous Humor of the Study Groups
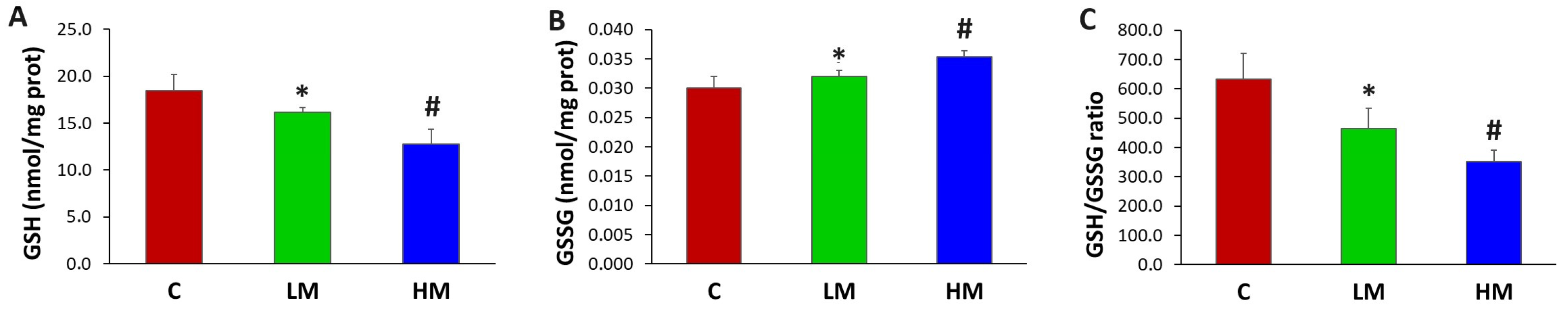
3.4. All Derivate Glutathione Biomarkers Showed Significant Differences among All Patient Groups
4. Discussion
4.1. NMR Metabolomic and Chemometric Approaches
4.2. Direct Oxidative-Stress-Related Metabolites
4.3. Scleral Remodeling and Metabolites Involved in Bioenergetic Pathways
4.4. Choline and Choline Derivates
4.5. Methylated Metabolites
4.6. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baird, P.N.; Saw, S.M.; Lanca, C.; Guggenheim, J.A.; Smith, E.L., III; Zhou, X.; Matsui, K.H.; Wu, P.C.; Sankaridurg, P.; Chia, A.; et al. Myopia. Nat. Rev. Dis. Primers 2020, 6, 99. [Google Scholar] [CrossRef]
- Holden, B.A.; Fricke, T.R.; Wilson, D.A.; Jong, M.; Naidoo, K.S.; Sankaridurg, P.; Wong, T.Y.; Naduvilath, T.J.; Resnikoff, S. Global prevalence of myopia and high myopia and temporal trends from 2000 through 2050. Ophthalmology 2016, 123, 1036–1042. [Google Scholar] [CrossRef]
- Han, X.; Liu, C.; Chen, Y.; He, M. Myopia prediction: A systematic review. Eye 2022, 36, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Zhou, J.B. Scleral remodeling in myopia development. Int. J. Ophthalmol. 2022, 15, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Ostrin, L.A.; Harb, E.; Nickla, D.L.; Read, S.A.; Alonso-Caneiro, D.; Schroedl, F.; Kaser-Eichberger, A.; Zhou, X.; Wildsoet, C.F. IMI-The Dynamic Choroid: New Insights, Challenges, and Potential Significance for Human Myopia. Investig. Ophthalmol. Vis. Sci. 2023, 64, 6. [Google Scholar] [CrossRef]
- Cai, X.B.; Shen, S.R.; Chen, D.F.; Zhang, Q.; Jin, Z.B. An overview of myopia genetics. Exp. Eye Res. 2019, 188, 107778. [Google Scholar] [CrossRef]
- Jiang, Y.; Xiao, X.; Sun, W.; Wang, Y.; Li, S.; Jia, X.; Wang, P.; Hejtmancik, J.F.; Zhang, Q. Clinical and genetic risk factors underlying severe consequence identified in 75 families with unilateral high myopia. J. Transl. Med. 2024, 22, 75. [Google Scholar] [CrossRef] [PubMed]
- Morgan, I.G.; Ohno-Matsui, K.; Saw, S.M. Myopia. Lancet 2012, 379, 1739–1748. [Google Scholar] [CrossRef]
- Haarman, A.E.; Enthoven, C.A.; Tideman, J.W.L.; Tedja, M.S.; Verhoeven, V.J.; Klaver, C.C. The complications of myopia: A review and meta-analysis. Investig. Ophthalmol. Vis. Sci. 2020, 61, 49. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Kim, S.S. Factors associated with axial length elongation in high myopia in adults. Int. J. Opthalmol. 2021, 14, 1231–1236. [Google Scholar] [CrossRef]
- Ohno-Matsui, K.; Wu, P.C.; Yamashiro, K.; Vutipongsatorn, K.; Fang, Y.; Cheung, C.M.G.; Lai, T.Y.Y.; Ikuno, Y.; Cohen, S.Y.; Gaudric, A.; et al. IMI pathologic myopia. Investig. Ophthalmol. Vis. Sci. 2021, 62, 5. [Google Scholar] [CrossRef] [PubMed]
- Freddo, T.F. A contemporary concept of the blood-aqueous barrier. Prog. Retin. Eye Res. 2013, 32, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Barbas-Bernardos, C.; Armitage, E.G.; García, A.; Mérida, S.; Navea, A.; Bosch-Morell, F.; Barbas, C. Looking into aqueous humor through metabolomics spectacles− exploring its metabolic characteristics in relation to myopia. J. Pharm. Biomed. Anal. 2016, 127, 18–25. [Google Scholar] [CrossRef]
- Mayordomo-Febrer, A.; López-Murcia, M.; Morales-Tatay, J.M.; Monleón-Salvado, D.; Pinazo-Durán, M.D. Metabolomics of the aqueous humor in the rat glaucoma model induced by a series of intracamerular sodium hyaluronate injection. Exp. Eye Res. 2015, 131, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond Biomarkers and towards Mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef]
- Sengupta, A.; Narad, P. Metabolomics. In Metabolomics, Omics Approaches, Technologies and Applications: Integrative Approaches for Understanding OMICS Data; Arivaradarajan, P., Misra, G., Eds.; Springer: Singapore, 2018; pp. 75–97. ISBN 978-981-13-2924-1. [Google Scholar]
- Francisco, B.M.; Salvador, M.; Amparo, N. Oxidative stress in myopia. Oxid. Med. Cell. Longev. 2015, 2015, 750637. [Google Scholar] [CrossRef]
- Mérida, S.; Villar, V.M.; Navea, A.; Desco, C.; Sancho-Tello, M.; Peris, C.; Bosch-Morell, F. Imbalance between oxidative stress and growth factors in human high myopia. Front. Physiol. 2020, 11, 463. [Google Scholar] [CrossRef]
- Flores-Moreno, I.; Lugo, F.; Duker, J.S.; Ruiz-Moreno, J.M. The relationship between axial length and choroidal thickness in eyes with high myopia. Am. J. Ophthalmol. 2013, 155, 314–319.e1. [Google Scholar] [CrossRef]
- Hosoda, Y.; Yoshikawa, M.; Miyake, M.; Tabara, Y.; Shimada, N.; Zhao, W.; Oishi, A.; Nakanishi, H.; Masayuki, H.; Akagi, T.; et al. CCDC102B confers risk of low vision and blindness in high myopia. Nat. Commun. 2018, 9, 1782. [Google Scholar] [CrossRef]
- Ajenjo, M.A.D.; Domene, M.C.G.; Martínez, C.P. Refractive changes in nuclear, cortical and posterior subcapsular cataracts. Effect of the type and grade. J. Optom. 2015, 8, 86–92. [Google Scholar] [CrossRef]
- Pan, C.W.; Boey, P.Y.; Cheng, C.Y.; Saw, S.M.; Tay, W.T.; Wang, J.J.; Tan, A.G.; Mitchell, P.; Wong, T.Y. Myopia, axial length, and age-related cataract: The Singapore Malay eye study. Investig. Ophthalmol. Vis. Sci. 2013, 54, 4498–4502. [Google Scholar] [CrossRef] [PubMed]
- Flitcroft, D.I.; He, M.; Jonas, J.B.; Jong, M.; Naidoo, K.; Ohno-Matsui, K.; Rahi, J.; Resnikoff, S.; Vitale, S.; Yannuzzi, L. IMI—Defining and Classifying Myopia: A Proposed Set of Standards for Clinical and Epidemiologic Studies. Investig. Ophthalmol. Vis. Sci. 2019, 60, M20–M30. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.J.; Babson, J.R.; Beatty, P.W.; Brodie, A.E.; Ellis, W.W.; Potter, D.W. High-performance liquid chromatography analysis of nanomole levels of glutathione, glutathione disulfide, and related thiols and disulfides. Anal. Biochem. 1980, 106, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Jin, N.; Zhu, X.; Lu, D.; Jin, C.; Li, Z.; Han, C.; Zhang, Y.; Lai, D.; Liu, K.; et al. A prospective study of serum metabolomic and lipidomic changes in myopic children and adolescents. Exp. Eye Res. 2020, 199, 108182. [Google Scholar] [CrossRef] [PubMed]
- Soga, T.; Baran, R.; Suematsu, M.; Ueno, Y.; Ikeda, S.; Sakurakawa, T.; Kakazu, Y.; Ishikawa, T.; Robert, M.; Nishioka, T.; et al. Differential metabolomics reveals ophthalmic acid as an oxidative stress biomarker indicating hepatic glutathione consumption. J. Biol. Chem. 2006, 281, 16768–16776. [Google Scholar] [CrossRef] [PubMed]
- Irino, Y.; Toh, R.; Nagao, M.; Mori, T.; Honjo, T.; Shinohara, M.; Tsuda, S.; Nakajima, S.; Satomi-Kobayashi, S.; Shinke, T.; et al. 2-Aminobutyric acid modulates glutathione homeostasis in the myocardium. Sci. Rep. 2016, 6, 36749. [Google Scholar] [CrossRef] [PubMed]
- Ueno, M.; Yoshii, K.; Yamashita, T.; Sonomura, K.; Asada, K.; Ito, E.; Fujita, T.; Sotozono, C.; Kinoshita, S.; Hamuro, J. The interplay between metabolites and microRNAs in aqueous humor to coordinate corneal endothelium integrity. Ophthalmol. Sci. 2023, 3, 100299. [Google Scholar] [CrossRef] [PubMed]
- Servillo, L.; Castaldo, D.; Giovane, A.; Casale, R.; D’Onofrio, N.; Cautela, D.; Balestrieri, M.L. Ophthalmic acid is a marker of oxidative stress in plants as in animals. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Tribble, J.R.; Otmani, A.; Sun, S.; Ellis, S.A.; Cimaglia, G.; Vohra, R.; Jöe, M.; Lardner, E.; Venkataraman, A.P.; Domínguez-Vicent, A.; et al. Nicotinamide provides neuroprotection in glaucoma by protecting against mitochondrial and metabolic dysfunction. Redox. Biol. 2021, 43, 101988. [Google Scholar] [CrossRef]
- Tuell, D.; Ford, G.; Los, E.; Stone, W. The Role of Glutathione and Its Precursors in Type 2 Diabetes. Antioxidants 2024, 13, 184. [Google Scholar] [CrossRef]
- Corkey, B.E.; Deeney, J.T. The redox communication network as a regulator of metabolism. Front. Physiol. 2020, 11, 567796. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.Y.; Chen, C.; Chang, J.; Chien, L.; Shih, Y.F.; Lin, L.L.; Pang, C.P.; Wang, I.J. Pharmacotherapeutic candidates for myopia: A review. Biomed. Pharmacother. 2021, 133, 111092. [Google Scholar] [CrossRef] [PubMed]
- Penha, A.M.; Schaeffel, F.; Feldkaemper, M. Insulin, insulin-like growth factor–1, insulin receptor, and insulin-like growth factor–1 receptor expression in the chick eye and their regulation with imposed myopic or hyperopic defocus. Mol. Vis. 2011, 17, 1436–1448. [Google Scholar] [PubMed]
- Vessey, K.A.; Rushforth, D.A.; Stell, W.K. Glucagon-and secretin-related peptides differentially alter ocular growth and the development of form-deprivation myopia in chicks. Investig. Ophthalmol. Vis. Sci. 2005, 46, 3932–3942. [Google Scholar] [CrossRef] [PubMed]
- Razak, M.A.; Begum, P.S.; Viswanath, B.; Rajagopal, S. Multifarious beneficial effect of nonessential amino acid, glycine: A review. Oxid. Med. Cell. Longev. 2017, 2017, 1716701. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, E.; Müller, U.; Wässle, H. Characterization of the glycinergic input to bipolar cells of the mouse retina. Eur. J. Neurosci. 2006, 23, 350–364. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Wang, Q.; Zhao, F.; Tang, G.; So, C.; Tse, D.; To, C.H.; Feng, Y.; Zhou, X.; Pan, F. Increased Connexin36 phosphorylation in AII amacrine cell coupling of the mouse myopic retina. Front. Cell. Neurosci. 2020, 14, 124. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Tong, P.; Chen, G.; Deng, M.; Cai, T.; Tian, R.; Zhang, Z.; Zhang, Z.; Xia, K.; Hu, Z. GLRA2 gene mutations cause high myopia in humans and mice. J. Med. Genet. 2023, 60, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Li, F.F.; Zhu, M.C.; Shao, Y.L.; Lu, F.; Yi, Q.Y.; Huang, X.F. Causal relationships between glycemic traits and myopia. Investig. Ophthalmol. Vis. Sci. 2023, 64, 7. [Google Scholar] [CrossRef]
- Zhu, B.T. Biochemical mechanism underlying the pathogenesis of diabetic retinopathy and other diabetic complications in humans: The methanol-formaldehyde-formic acid hypothesis. Acta Biochim. Biophys. Sin. 2022, 54, 415–451. [Google Scholar] [CrossRef]
- Rabinowitz, J.D.; Enerbäck, S. Lactate: The ugly duckling of energy metabolism. Nat. Metab. 2020, 2, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Jha, M.K.; Lee, I.K.; Suk, K. Metabolic reprogramming by the pyruvate dehydrogenase kinase–lactic acid axis: Linking metabolism and diverse neuropathophysiologies. Neurosci. Biobehav. Rev. 2016, 68, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Lei, Y.; Pan, M.; Hu, C.; Xie, B.; Wu, W.; Su, J.; Li, Y.; Tan, Y.; Wei, X.; et al. Augmentation of scleral glycolysis promotes myopia through histone lactylation. Cell Metab. 2024, 36, 511–525. [Google Scholar] [CrossRef] [PubMed]
- Tauffenberger, A.; Fiumelli, H.; Almustafa, S.; Magistretti, P.J. Lactate and pyruvate promote oxidative stress resistance through hormetic ROS signaling. Cell Death Dis. 2019, 10, 653. [Google Scholar] [CrossRef] [PubMed]
- Tassinari, I.D.; Andrade, M.K.G.; da Rosa, L.A.; Hoff, M.L.M.; Nunes, R.R.; Vogt, E.L.; Fabres, R.B.; Sanches, E.F.; Netto, C.A.; Paz, A.H.; et al. Lactate administration reduces brain injury and ameliorates behavioral outcomes following neonatal hypoxia-ischemia. Neuroscience 2020, 448, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Cai, M.; Liu, Y.; Liu, B.; Xue, X.; Ji, R.; Bian, X.; Lou, S. The roles of GRP81 as a metabolic sensor and inflammatory mediator. J. Cell. Physiol. 2020, 235, 8938–8950. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Wang, Z.; Liang, X.; Wang, Y.; Gao, L.; Ma, C. Monocarboxylate transporter 1 promotes classical microglial activation and pro-inflammatory effect via 6-phosphofructo-2-kinase/fructose-2, 6-biphosphatase 3. J. Neuroinflamm. 2019, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jensen, N.J.; Wodschow, H.Z.; Nilsson, M.; Rungby, J. Effects of ketone bodies on brain metabolism and function in neurodegenerative diseases. Int. J. Mol. Sci. 2020, 21, 8767. [Google Scholar] [CrossRef] [PubMed]
- Izuta, Y.; Imada, T.; Hisamura, R.; Oonishi, E.; Nakamura, S.; Inagaki, E.; Ito, M.; Soga, T.; Tsubota, K. Ketone body 3-hydroxybutyrate mimics calorie restriction via the Nrf2 activator, fumarate, in the retina. Aging Cell 2018, 17, e12699. [Google Scholar] [CrossRef]
- Nelson, A.B.; Queathem, E.D.; Puchalska, P.; Crawford, P.A. Metabolic Messengers: Ketone bodies. Nat. Metab. 2023, 5, 2062–2074. [Google Scholar] [CrossRef]
- Rojas-Morales, P.; Pedraza-Chaverri, J.; Tapia, E. Ketone bodies, stress response, and redox homeostasis. Redox Biol. 2020, 29, 101395. [Google Scholar] [CrossRef]
- Acosta, M.L.; Kalloniatis, M.; Christie, D.L. Creatine transporter localization in developing and adult retina: Importance of creatine to retinal function. Am. J. Physiol. Cell Physiol. 2005, 289, C1015–C1023. [Google Scholar] [CrossRef] [PubMed]
- Sia, P.I.; Wood, J.P.; Chidlow, G.; Casson, R. Creatine is neuroprotective to retinal neurons in vitro but not in vivo. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4360–4377. [Google Scholar] [CrossRef]
- Ke, C.; Xu, H.; Chen, Q.; Zhong, H.; Pan, C.W. Serum metabolic signatures of high myopia among older Chinese adults. Eye 2021, 35, 817–824. [Google Scholar] [CrossRef]
- Cui, D.; Trier, K.; Ribel-Madsen, S.M. Effect of day length on eye growth, myopia progression, and change of corneal power in myopic children. Ophthalmology 2013, 120, 1074–1079. [Google Scholar] [CrossRef]
- Cui, D.; Trier, K.; Zeng, J.; Wu, K.; Yu, M.; Ge, J. Adenosine receptor protein changes in guinea pigs with form deprivation myopia. Acta Ophthalmol. 2010, 88, 759–765. [Google Scholar] [CrossRef]
- Santiago, A.R.; Madeira, M.H.; Boia, R.; Aires, I.D.; Rodrigues-Neves, A.C.; Santos, P.F.; Ambrosio, A.F. Keep an eye on adenosine: Its role in retinal inflammation. Pharmacol. Ther. 2020, 210, 107513. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.; Patzak, A.; Pfeiffer, N.; Gericke, A. Muscarinic acetylcholine receptors in the retina—Therapeutic implications. Int. J. Mol. Sci. 2021, 22, 4989. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ouyang, X.; Fu, H.; Hou, X.; Liu, Y.; Xie, Y.; Yu, H.; Wang, G. Advances in biomedical study of the myopia-related signaling pathways and mechanisms. Biomed. Pharmacother. 2022, 145, 112472. [Google Scholar] [CrossRef]
- Ford, K.J.; Feller, M.B. Assembly and disassembly of a retinal cholinergic network. Vis. Neurosci. 2012, 29, 61–71. [Google Scholar] [CrossRef]
- Laspas, P.; Zhutdieva, M.B.; Brochhausen, C.; Musayeva, A.; Zadeh, J.K.; Pfeiffer, N.; Xia, N.; Li, H.; Wess, J.; Gericke, A. The M1 muscarinic acetylcholine receptor subtype is important for retinal neuron survival in aging mice. Sci. Rep. 2019, 9, 5222. [Google Scholar] [CrossRef]
- Barathi, V.A.; Kwan, J.L.; Tan, Q.S.; Weon, S.R.; Seet, L.F.; Goh, L.K.; Vithana, E.N.; Beuerman, R.W. Muscarinic cholinergic receptor (M2) plays a crucial role in the development of myopia in mice. Dis Model Mech 2013, 6, 1146–1158. [Google Scholar] [CrossRef]
- Gericke, A.; Steege, A.; Manicam, C.; Böhmer, T.; Wess, J.; Pfeiffer, N. Role of the M3 muscarinic acetylcholine receptor subtype in murine ophthalmic arteries after endothelial removal. Investig. Ophthalmol. Vis. Sci. 2014, 55, 625–631. [Google Scholar] [CrossRef]
- Kiuchi, K.; Matsuoka, M.; Wu, J.C.; Silva, R.L.; Kengatharan, M.; Verghese, M.; Ueno, S.; Yokoi, K.; Khu, N.H.; Cooke, J.P.; et al. Mecamylamine suppresses basal and nicotine-stimulated choroidal neovascularization. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1705–1711. [Google Scholar] [CrossRef] [PubMed]
- Gallazzini, M.; Ferraris, J.D.; Burg, M.B. GDPD5 is a glycerophosphocholine phosphodiesterase that osmotically regulates the osmoprotective organic osmolyte GPC. Proc. Natl. Acad. Sci. USA 2008, 105, 11026–11031. [Google Scholar] [CrossRef]
- Wu, W.; Song, Y.; Sun, M.; Li, Y.; Xu, Y.; Xu, M.; Yang, Y.; Li, S.; Zhang, F. Corneal metabolic biomarkers for moderate and high myopia in human. Exp. Eye Res. 2023, 237, 109689. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Xu, Y.; Pang, Z.; Mu, G. The influence of the choroid on the onset and development of myopia: From perspectives of choroidal thickness and blood flow. Acta Ophthalmol. 2021, 99, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Cunha-Vaz, J.G. The blood-ocular barriers: Past, present, and future. Doc. Ophthalmol. 1997, 93, 149–157. [Google Scholar] [CrossRef]
- Marshall, D.L.; Criscuolo, A.; Young, R.S.; Poad, B.L.; Zeller, M.; Reid, G.E.; Mitchell, T.W.; Blanksby, S.J. Mapping unsaturation in human plasma lipids by data-independent ozone-induced dissociation. J. Am. Soc. Mass Spectrom. 2019, 30, 1621–1630. [Google Scholar] [CrossRef]
- Wu, J.; Liu, L.L.; Cao, M.; Hu, A.; Hu, D.; Luo, Y.; Wang, H.; Zhong, J.N. DNA Methylation plays important roles in retinal development and diseases. Exp. Eye Res. 2021, 211, 108733. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Grenell, A.; Zhong, F.; Yam, M.; Hauer, A.; Gregor, E.; Zhu, S.; Lohner, D.; Zhu, J.; Du, J. Metabolic signature of the aging eye in mice. Neurobiol. Aging 2018, 71, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Advani, J.; Mehta, P.A.; Hamel, A.R.; Mehrotra, S.; Kiel, C.; Strunz, T.; Corso-Díaz, X.; Kwicklis, M.; van Asten, F.; Ratnapriya, R.; et al. QTL mapping of human retina DNA methylation identifies 87 gene-epigenome interactions in age-related macular degeneration. Nat. Commun. 2024, 15, 1972. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Axial Length (mm) | Spherical Equivalent | Macular Thickness (μm) | Choroidal Thickness (μm) | |
|---|---|---|---|---|---|
| C | 40 | 22.6 ± 0.4 | −1.1 ± 1.5 | 259.2 ± 33.0 | 237.1 ± 64.5 |
| LM | 48 | 24.2 ± 0.7 * | −3.4 ± 3.0 * | 249.7 ± 26.3 | 196.2 ± 77.3 |
| HM | 28 | 28.4 ± 1.9 *# | −9.7 ± 7.6 *# | 269.7 ± 42.4 | 125.5 ± 106.7 *# |
| Degrees of Maculopathy | C | LM | HM |
|---|---|---|---|
| No myopic degenerative retinal lesion | 29.5 | 32.8 | 0.8 |
| Tessellated fundus | 1.6 | 5.7 | 3.3 |
| Diffuse chorioretinal atrophy | 0 | 0 | 5.7 |
| Patchy chorioretinal atrophy | 0.2 | 0.5 | 1.3 |
| Macular atrophy | 0 | 0 | 5.7 |
| PLUS: NEOVASCULARIZATION/FUCHS | 0 | 0 | 4.1 |
| PLUS: LACQUER CRACKS | 0 | 0 | 4.9 |
| ID | Formula | LM %change | HM %change |
|---|---|---|---|
| 2-Aminobutyrate | C4H9NO2 | −12.54 | +18.64 # |
| Acetate | C2H4O2 | +40.87 * | +12.72 |
| Choline | C5H14NO | +119.66 * | +9.39 *# |
| Glycine | C2H5NO2 | +5.13 | −21.45 # |
| Lactate | C3H6O3 | +2.22 | −8.40 # |
| sn-Glycero-3-phosphocholine | C8H21NO6P | −19.19 * | −15.47 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mérida, S.; Návea, A.; Desco, C.; Celda, B.; Pardo-Tendero, M.; Morales-Tatay, J.M.; Bosch-Morell, F. Glutathione and a Pool of Metabolites Partly Related to Oxidative Stress Are Associated with Low and High Myopia in an Altered Bioenergetic Environment. Antioxidants 2024, 13, 539. https://doi.org/10.3390/antiox13050539
Mérida S, Návea A, Desco C, Celda B, Pardo-Tendero M, Morales-Tatay JM, Bosch-Morell F. Glutathione and a Pool of Metabolites Partly Related to Oxidative Stress Are Associated with Low and High Myopia in an Altered Bioenergetic Environment. Antioxidants. 2024; 13(5):539. https://doi.org/10.3390/antiox13050539
Chicago/Turabian StyleMérida, Salvador, Amparo Návea, Carmen Desco, Bernardo Celda, Mercedes Pardo-Tendero, José Manuel Morales-Tatay, and Francisco Bosch-Morell. 2024. "Glutathione and a Pool of Metabolites Partly Related to Oxidative Stress Are Associated with Low and High Myopia in an Altered Bioenergetic Environment" Antioxidants 13, no. 5: 539. https://doi.org/10.3390/antiox13050539
APA StyleMérida, S., Návea, A., Desco, C., Celda, B., Pardo-Tendero, M., Morales-Tatay, J. M., & Bosch-Morell, F. (2024). Glutathione and a Pool of Metabolites Partly Related to Oxidative Stress Are Associated with Low and High Myopia in an Altered Bioenergetic Environment. Antioxidants, 13(5), 539. https://doi.org/10.3390/antiox13050539