
In Vitro Investigation of Six Antioxidants for Pig Diets

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Line and Culture Conditions
2.2. Induction of Oxidative Stress
2.3. Antioxidant Pre-Treatments
2.4. Viability (Neutral Red)
2.5. Intracellular Oxidative Stress
2.6. Permeability Assay
2.7. Transepithelial Electrical Resistance (TEER)
2.8. Statistical Analysis
3. Results
3.1. Effect of Antioxidants on the Viability of 1 mM H2O2 Stressed IPEC-J2 cells
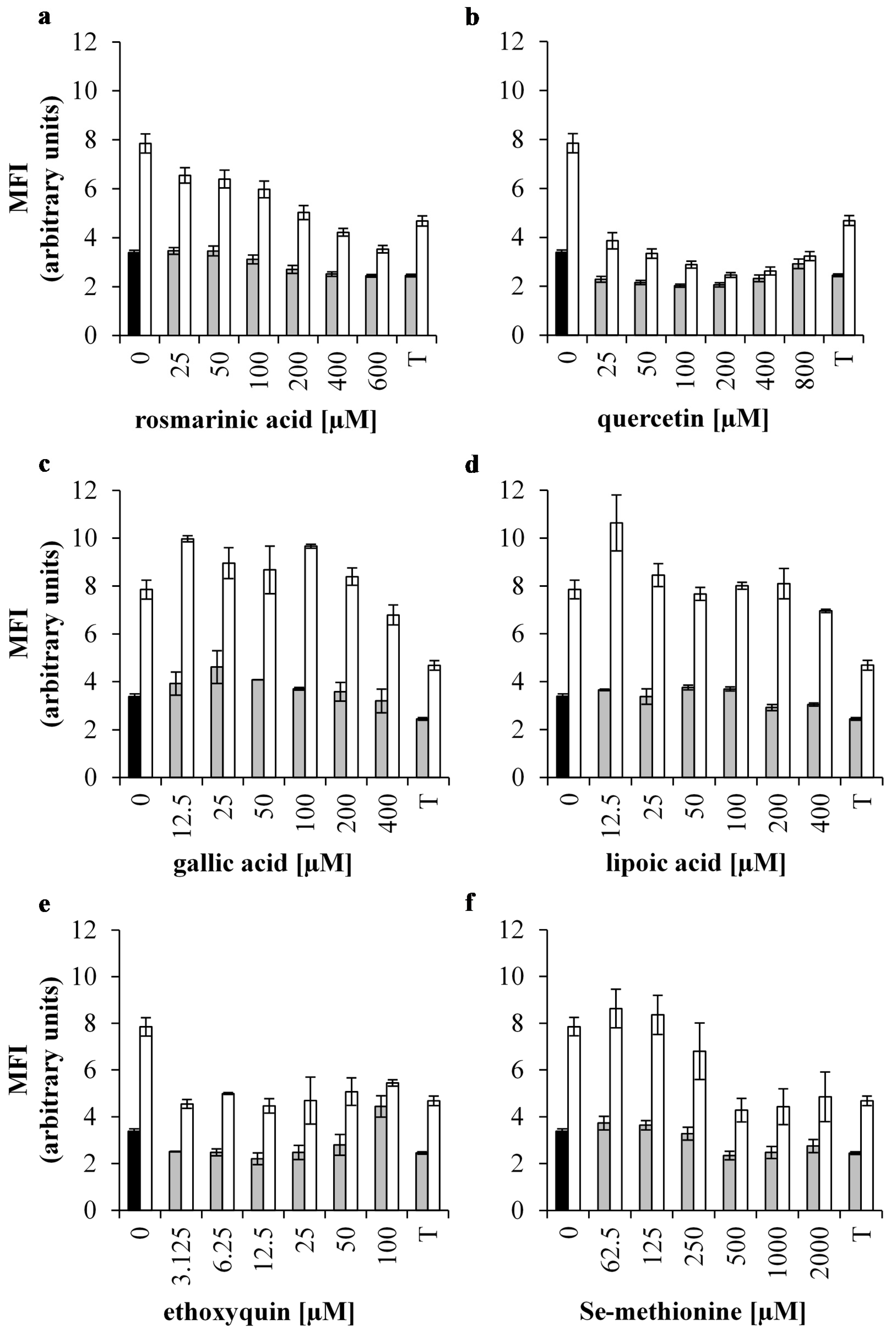
3.2. Effect of Antioxidants on Intracellular Oxidative Stress of 0.5 mM H2O2 Treated IPEC-J2 Cells
3.3. Protective Effect of Quercetin and Rosmarinic Acid on the Permeability and Integrity of A 1 mM H2O2 Treated IPEC-J2 Monolayer
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Halliwell, B. Antioxidants in human health and disease. Ann. Rev. Nutr. 1996, 16, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Rosero, D.S.; Odle, J.; Moeser, A.J.; Boyd, R.D.; van Heugten, E. Peroxidised dietary lipids impair intestinal function and morphology of the small intestine villi of nursery pigs in a dose-dependent manner. Brit. J. Nutr. 2015, 114, 1985–1992. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.C.; Yang, S.F.; Zhu, L.H.; Cai, X.; Sheng, Y.S.; Zhu, S.W.; Xu, J.X. Regulation of N-acetyl cysteine on gut redox status and major microbiota in weaned piglets1. J. Anim. Sci. 2014, 92, 1504–1511. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xu, C.; Chen, X.; Cai, X.; Yang, S.; Sheng, Y.; Wang, T. Regulation of an antioxidant blend on intestinal redox status and major microbiota in early weaned piglets. Nutrition 2014, 30, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Aw, T.Y. Molecular and cellular responses to oxidative stress and changes in oxidation-reduction imbalance in the intestine. Am. J. Clin. Nutr. 1999, 70, 557–565. [Google Scholar] [PubMed]
- Zhang, K.; Hornef, M.W.; Dupont, A. The intestinal epithelium as guardian of gut barrier integrity. Cell. Microbiol. 2015, 17, 1561–1569. [Google Scholar] [CrossRef] [PubMed]
- Aw, T.Y. Intestinal glutathione: Determinant of mucosal peroxide transport, metabolism, and oxidative susceptibility. Toxicol. Appl. Pharm. 2005, 204, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.; Choi, S.W. Antioxidants in food: Content, measurement, significance, action, cautions, caveats, and research needs. Adv. Food Nutr. Res. 2014, 71, 1–53. [Google Scholar] [PubMed]
- Qiang, Z.; Ye, Z.; Hauck, C.; Murphy, P.A.; McCoy, J.A.; Widrlechner, M.P.; Reddy, M.B.; Hendrich, S. Permeability of rosmarinic acid in prunella vulgaris and ursolic acid in salvia officinalis extracts across caco-2 cell monolayers. J. Ethnopharmacol. 2011, 137, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Liu, D.; Yu, X.; Sun, H.; Li, Y. A Caco-2 cell-based quantitative antioxidant activity assay for antioxidants. Food Chem. 2015, 175, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Vergauwen, H.; Tambuyzer, B.; Jennes, K.; Degroote, J.; Wang, W.; De Smet, S.; Michiels, J.; van Ginneken, C. Trolox and ascorbic acid reduce direct and indirect oxidative stress in the IPEC-J2 cells, an in vitro model for the porcine gastrointestinal tract. PLoS ONE 2015, 10, e0120485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farkas, O.; Palocz, O.; Paszti-Gere, E.; Galfi, P. Polymethoxyflavone apigenin-trimethylether suppresses LPS-induced inflammatory response in nontransformed porcine intestinal cell line IPEC-J2. Oxid. Med. Cell. Longev. 2015, 2015, 673847. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative stress: An essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Degroote, J.; van Ginneken, C.; van Poucke, M.; Vergauwen, H.; Dam, T.M.; Vanrompay, D.; Peelman, L.J.; De Smet, S.; Michiels, J. Intrauterine growth restriction in neonatal piglets affects small intestinal mucosal permeability and mrna expression of redox-sensitive genes. FASEB J. 2015, 30, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Degroote, J.; Vergauwen, H.; Wang, W.; van Ginneken, C.; de Smet, S.; Michiels, J. Oxidative status in piglets is affected by the weaning process. Commun. Agric. Appl. Biol. Sci. 2015, 80, 183–188. [Google Scholar] [PubMed]
- Machlin, L.J.; Bendich, A. Free radical tissue damage: Protective role of antioxidant nutrients. FASEB J. 1987, 1, 441–445. [Google Scholar] [PubMed]
- Russell, E.G.; Cotter, T.G. New Insight into the Role of Reactive Oxygen Species (ROS) in Cellular Signal-Transduction Processes. Int. Rev. Cell Mol. Biol. 2015, 319, 221–254. [Google Scholar] [PubMed]
- Sultatos, L.G.; Huang, G.J.; Jackson, O.; Reed, K.; Soranno, T.M. The effect of glutathione monoethyl ester on the potentiation of the acute toxicity of methyl parathion, methyl paraoxon or fenitrothion by diethyl maleate in the mouse. Toxicol. Lett. 1991, 55, 77–83. [Google Scholar] [CrossRef]
- Ivison, S.M.; Wang, C.; Himmel, M.E.; Sheridan, J.; Delano, J.; Mayer, M.L.; Yao, Y.; Kifayet, A.; Steiner, T.S. Oxidative stress enhances IL-8 and inhibits CCL20 production from intestinal epithelial cells in response to bacterial flagellin. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G733–G741. [Google Scholar] [CrossRef] [PubMed]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Fan, P.X.; Li, L.S.; Qiao, S.Y.; Zhang, G.L.; Li, D.F. Butyrate promotes the recovering of intestinal wound healing through its positive effect on the tight junctions. J. Ani. Sci. 2012, 90, 266–268. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.P.; Wei, H.L.; Zhao, H.S.; Xiao, S.Y.; Zheng, R.L. Antiapoptotic and antioxidant effects of rosmarinic acid in astrocytes. Pharmazie 2005, 60, 62–65. [Google Scholar] [PubMed]
- Aherne, S.A.; O’Brien, N.M. Protection by the flavonoids myricetin, quercetin, and rutin against hydrogen peroxide-induced DNA damage in Caco-2 and Hep G2 cells. Nutr. Cancer 1999, 34, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Serrano, J.; Cipak, A.; Boada, J.; Gonzalo, H.; Cacabelos, D.; Cassanye, A.; Pamplona, R.; Zarkovic, N.; Portero-Otin, M. Double-edged sword behaviour of gallic acid and its interaction with peroxidases in human microvascular endothelial cell culture (HMEC-1). Antioxidant and pro-oxidant effects. Acta Biochim. Pol. 2010, 57, 193–198. [Google Scholar] [PubMed]
- Blaszczyk, A.; Skolimowski, J.; Materac, A. Genotoxic and antioxidant activities of ethoxyquin salts evaluated by the comet assay. Chem. Biol. Interact. 2006, 162, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Miranda, S.G.; Purdie, N.G.; Osborne, V.R.; Coomber, B.L.; Cant, J.P. Selenomethionine increases proliferation and reduces apoptosis in bovine mammary epithelial cells under oxidative stress. J. Dairy Sci. 2011, 94, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Flora, S.J.S. Structural, chemical and biological aspects of antioxidants for strategies against metal and metalloid exposure. Oxid. Med. Cell. Longev. 2009, 2, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Rahmanto, A.S.; Davies, M.J. Selenium-containing amino acids as direct and indirect antioxidants. IUBMB Life 2012, 64, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Min, K.; Ebeler, S.E. Quercetin inhibits hydrogen peroxide-induced DNA damage and enhances DNA repair in Caco-2 cells. Food Chem. Toxicol. 2009, 47, 2716–2722. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Meng, H.; Zhao, Y.; Chen, F.; Yu, S. Antioxidant and in vitro anticancer activities of phenolics isolated from sugar beet molasses. BMC Complement. Altern. Med. 2015, 15, 313. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Chen, X.; Wang, X.; Xu, C.; Guo, Q.; Zhu, L.; Zhu, S.; Xu, J. Pre-protective effect of lipoic acid on injury induced by H2O2 in IPEC-J2 cells. Mol. Cell. Biochem. 2013, 238. [Google Scholar] [CrossRef] [PubMed]
- Blaszczyk, A.; Skolimowski, J. Evaluation of the genotoxic and antioxidant effects of two novel feed additives (ethoxyquin complexes with flavonoids) by comet assay and micronucleus test. Food Addit. Contam. 2007, 24, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Fuerst, F.; Hotchkiss, E.; Boland, C.R. Selenomethionine induces p53 mediated cell cycle arrest and apoptosis in human colon cancer cells. Cancer Biol. Ther. 2006, 5, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Fadel, O.; El Kirat, K.; Morandat, S. The natural antioxidant rosmarinic acid spontaneously penetrates membranes to inhibit lipid peroxidation in situ. Biochim. Biophys. Acta 2011, 1808, 2973–2980. [Google Scholar] [CrossRef] [PubMed]
- Yokomizo, A.; Moriwaki, M. Effects of uptake of flavonoids on oxidative stress induced by hydrogen peroxide in human intestinal Caco-2 cells. Biosci. Biotech. Biochem. 2006, 70, 1317–1324. [Google Scholar] [CrossRef]
- Verma, S.; Singh, A.; Mishra, A. Gallic acid: Molecular rival of cancer. Environ. Toxicol. Phar. 2013, 35, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Erol-Dayi, O.; Arda, N.; Erdem, G. Protective effects of olive oil phenolics and gallic acid on hydrogen peroxide-induced apoptosis. Eur. J. Nutr. 2012, 51, 955–960. [Google Scholar] [CrossRef] [PubMed]
- Decker, E.A. Phenolics: Prooxidants or antioxidants? Nutr. Rev. 1997, 55, 396–398. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.K.; Baker, R.D.; Baker, S.S.; Gupta, A.; Holycross, M. Oxidant-induced disruption of intestinal epithelial barrier function: Role of protein tyrosine phosphorylation. Am. J. Physiol. 1997, 273, G812–G823. [Google Scholar] [PubMed]
- Welsh, M.J.; Shasby, D.M.; Husted, R.M. Oxidants increase paracellular permeability in a cultured epithelial cell line. J. Clin. Investig. 1985, 76, 1155–1168. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Hara, H. Quercetin enhances intestinal barrier function through the assembly of zonula [corrected] occludens-2, occludin, and claudin-1 and the expression of claudin-4 in Caco-2 cells. J. Nutr. 2009, 139, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Chuenkitiyanon, S.; Pengsuparp, T.; Jianmongkol, S. Protective effect of quercetin on hydrogen peroxide-induced tight junction disruption. Int. J. Toxicol. 2010, 29, 418–424. [Google Scholar] [CrossRef] [PubMed]




© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vergauwen, H.; Prims, S.; Degroote, J.; Wang, W.; Casteleyn, C.; Van Cruchten, S.; De Smet, S.; Michiels, J.; Van Ginneken, C. In Vitro Investigation of Six Antioxidants for Pig Diets. Antioxidants 2016, 5, 41. https://doi.org/10.3390/antiox5040041
Vergauwen H, Prims S, Degroote J, Wang W, Casteleyn C, Van Cruchten S, De Smet S, Michiels J, Van Ginneken C. In Vitro Investigation of Six Antioxidants for Pig Diets. Antioxidants. 2016; 5(4):41. https://doi.org/10.3390/antiox5040041
Chicago/Turabian StyleVergauwen, Hans, Sara Prims, Jeroen Degroote, Wei Wang, Christophe Casteleyn, Steven Van Cruchten, Stefaan De Smet, Joris Michiels, and Chris Van Ginneken. 2016. "In Vitro Investigation of Six Antioxidants for Pig Diets" Antioxidants 5, no. 4: 41. https://doi.org/10.3390/antiox5040041
APA StyleVergauwen, H., Prims, S., Degroote, J., Wang, W., Casteleyn, C., Van Cruchten, S., De Smet, S., Michiels, J., & Van Ginneken, C. (2016). In Vitro Investigation of Six Antioxidants for Pig Diets. Antioxidants, 5(4), 41. https://doi.org/10.3390/antiox5040041