
Effects of β-Carotene and Its Cleavage Products in Primary Pneumocyte Type II Cells


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of β-Carotene Cleavage Products (CPs)
2.3. Animals
2.4. Methods
2.4.1. Isolation and Culture of Pneumocyte Type II Cells
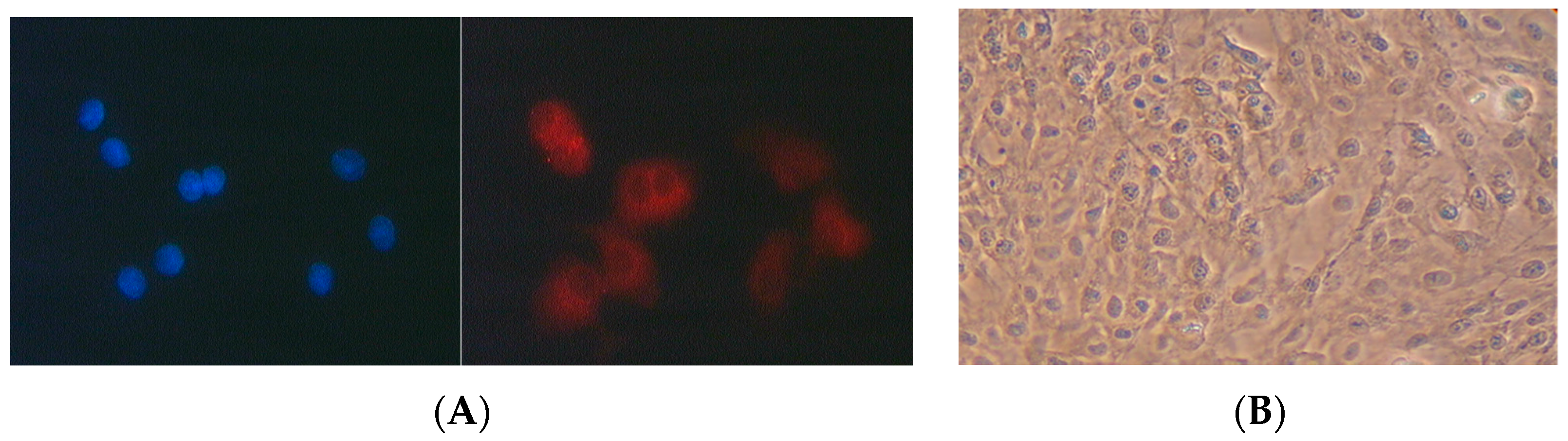
2.4.2. Characterisation of Pneumocyte Type II Cells
2.4.3. Treatment
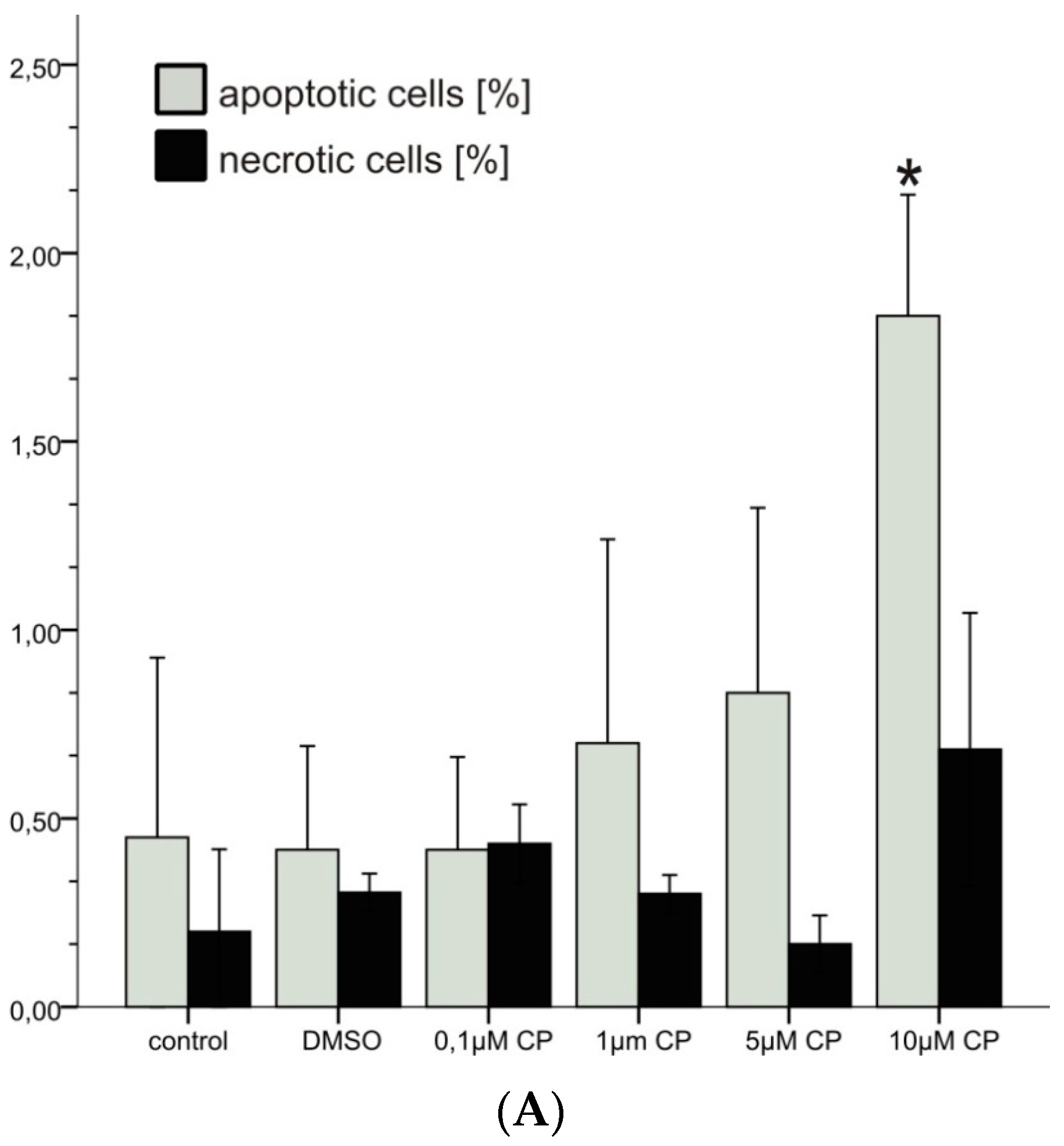
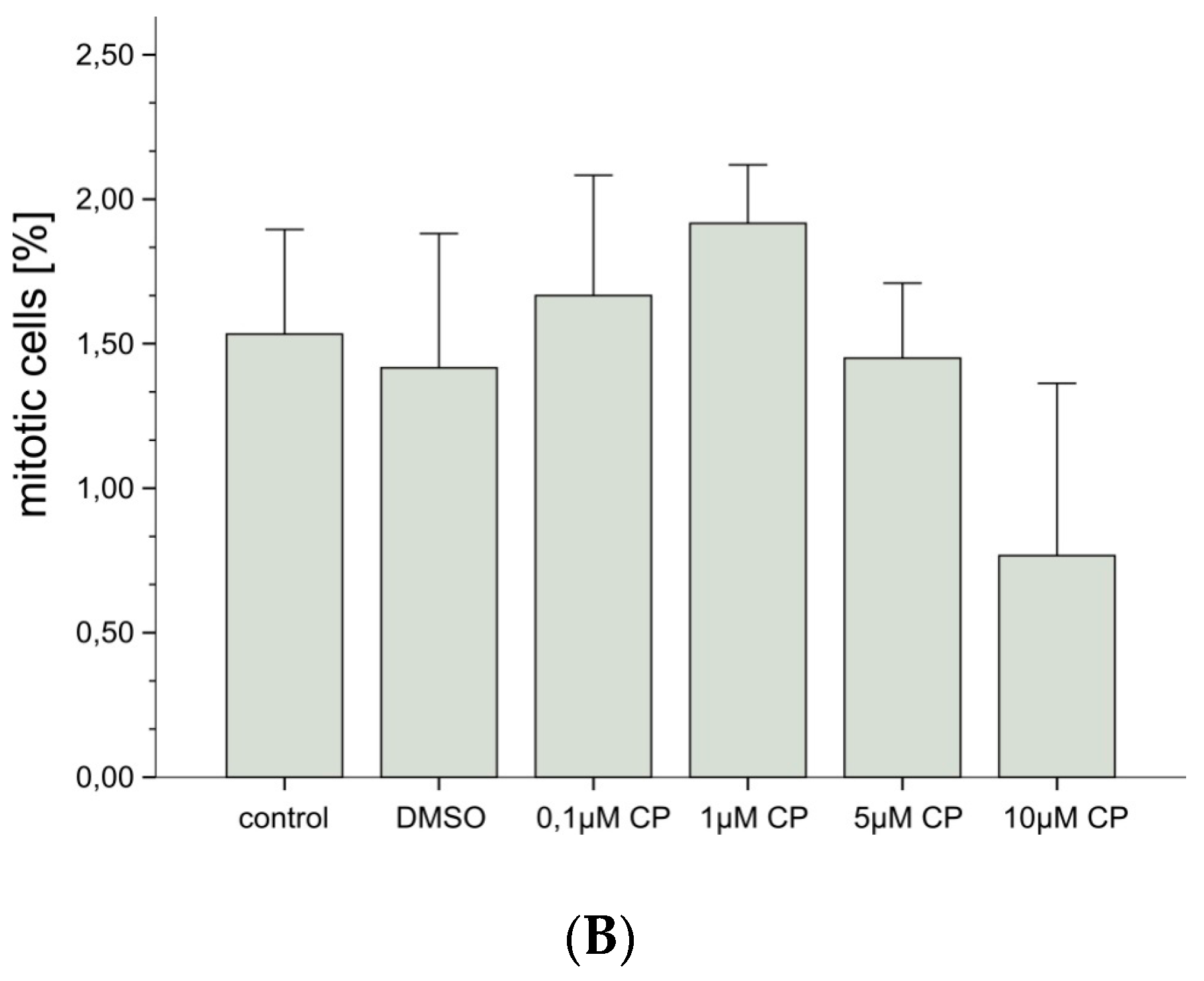
2.4.4. Fixation, Staining and Cytogenetic Analysis
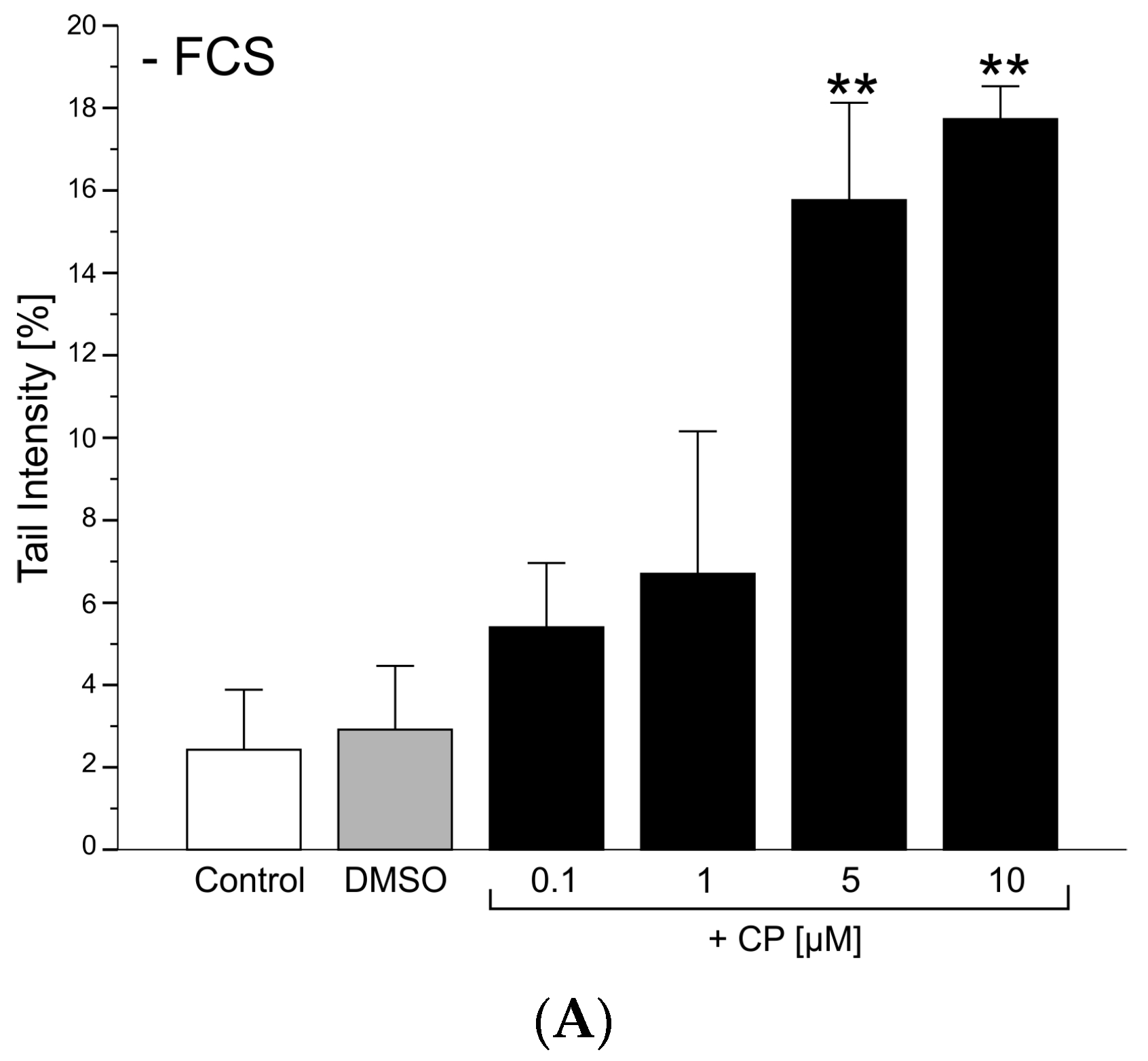
2.4.5. Comet Assay
2.4.6. Statistical Analysis
3. Results
3.1. Effects of CPs on Primary Pneumocytes
3.1.1. Cytotoxicity
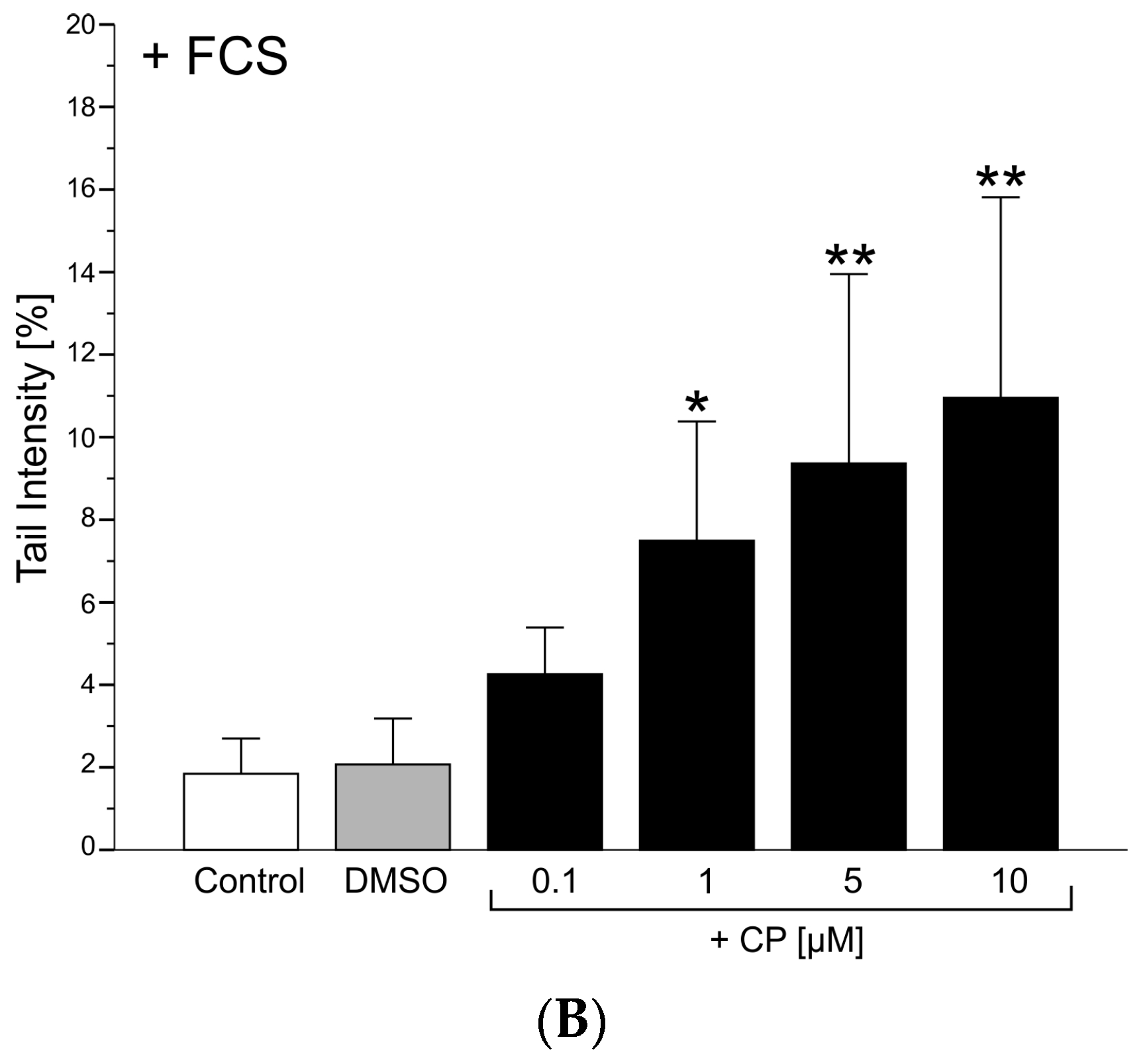
3.1.2. Genotoxicity
Comet Assay
3.2. Effects of β-Carotene in the Presence of Oxidative Stress
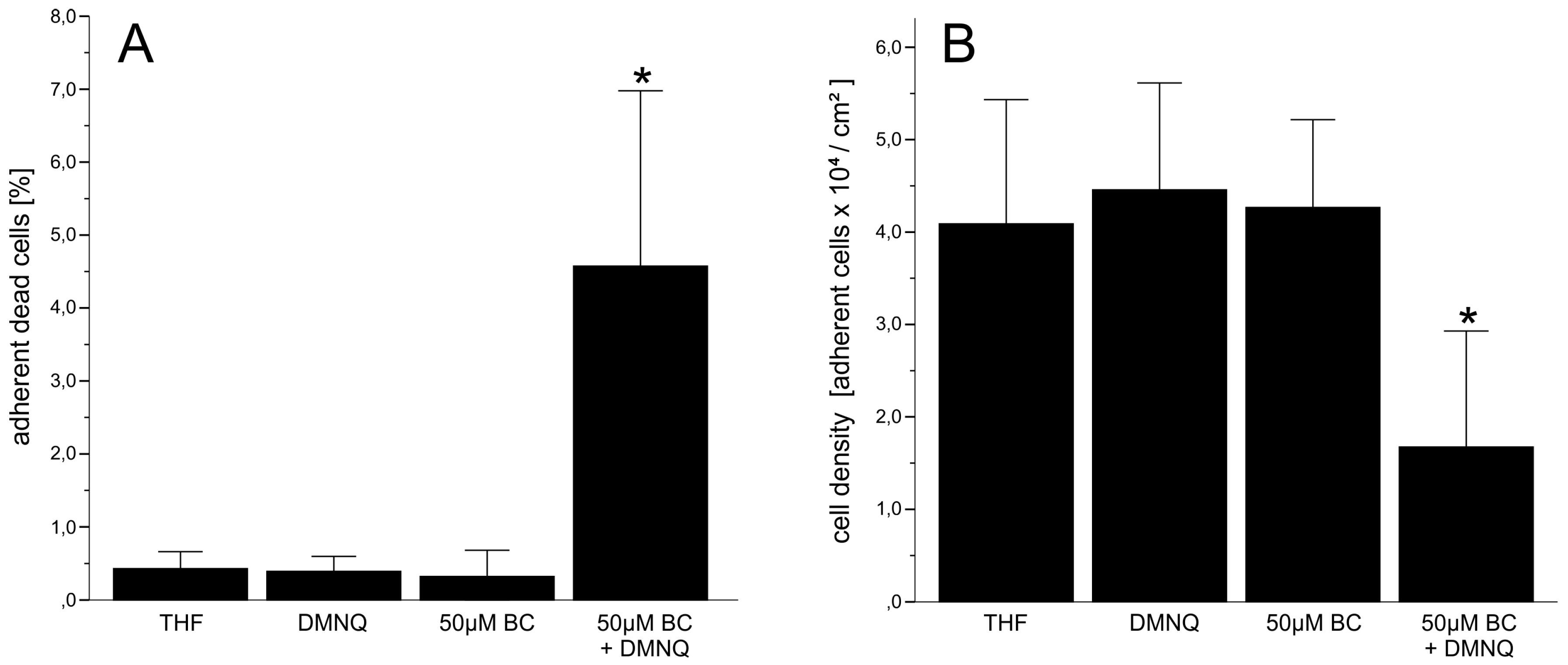
3.2.1. Cytotoxicity
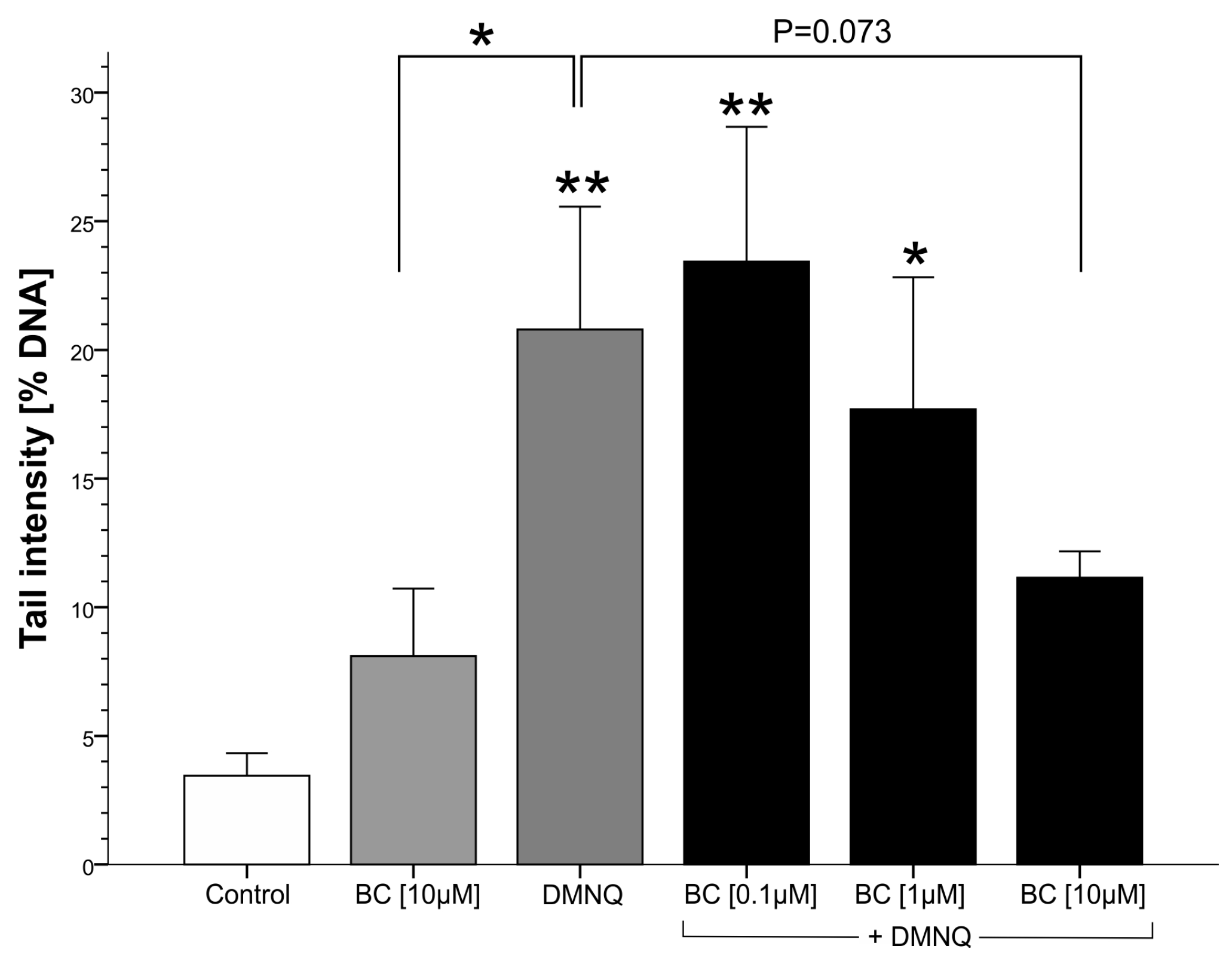
3.2.2. Genotoxicity
4. Discussion
4.1. β-Carotene Cleavage Products
4.2. β-Carotene under Oxidative Stress
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Burton, G.W. Antioxidant action of carotenoids. J. Nutr. 1989, 119, 109–111. [Google Scholar] [PubMed]
- Sies, H.; Stahl, W.; Sundquist, A.R. Antioxidant functions of vitamins. Ann. N. Y. Acad. Sci. 1992, 669, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, A.; Skibsted, L.H.; Sampson, J.; Rice-Evans, C.; Everett, S.A. Comparative mechanisms and rates of free radical scavenging by carotenoid antioxidants. FEBS Lett. 1997, 418, 91–97. [Google Scholar] [CrossRef]
- Foote, C.S.; Denny, R.W. Chemistry of singlet oxygen. VII. Quenching by. beta-carotene. J. Am. Chem. Soc. 1968, 90, 6233–6235. [Google Scholar] [CrossRef]
- Palozza, P.; Krinsky, N.I. The inhibition of radical-initiated peroxidation of microsomal lipids by both α-tocopherol and β-carotene. Free Radic. Biol. Med. 1991, 11, 407–414. [Google Scholar] [CrossRef]
- Tsuchihashi, H.; Kigoshi, M.; Iwatsuki, M.; Niki, E. Action of β-carotene as an antioxidant against lipid peroxidation. Arch. Biochem. Biophys. 1995, 323, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Gaziano, J.M.; Manson, J.E.; Branch, L.G.; Colditz, G.A.; Willett, W.C.; Buring, J.E. A prospective study of consumption of carotenoids in fruits and vegetables and decreased cardiovascular mortality in the elderly. Ann. Epidemiol. 1995, 5, 255–260. [Google Scholar] [CrossRef]
- Gey, K.F.; Moser, U.K.; Jordan, P.; Stähelin, H.B.; Eichholzer, M.; Lüdin, E. Increased risk of cardiovascular disease at suboptimal plasma concentrations of essential antioxidants: An epidemiological update with special attention to carotene and vitamin C. Am. J. Clin. Nutr. 1993, 57, 787S–797S. [Google Scholar] [PubMed]
- Ito, Y.; Kurata, M.; Suzuki, K.; Hamajima, N.; Hishida, H.; Aoki, K. Cardiovascular disease mortality and serum carotenoid levels: A Japanese population-based follow-up study. J. Epidemiol. 2006, 16, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Seddon, J.M.; Ajani, U.A.; Sperduto, R.D.; Hiller, R.; Blair, N.; Burton, T.C.; Farber, M.D.; Gragoudas, E.S.; Haller, J.; Miller, D.T.; et al. Dietary carotenoids, vitamins A, C, and E, and advanced age-related macular degeneration. JAMA 1994, 272, 1413–1420. [Google Scholar] [CrossRef] [PubMed]
- Gale, C.R.; Hall, N.F.; Phillips, D.I.; Martyn, C.N. Plasma antioxidant vitamins and carotenoids and age-related cataract. Ophthalmology 2001, 108, 1992–1998. [Google Scholar] [CrossRef]
- Jacques, P.F.; Chylack, L.T. Epidemiologic evidence of a role for the antioxidant vitamins and carotenoids in cataract prevention. Am. J. Clin. Nutr. 1991, 53, 352S–355S. [Google Scholar] [PubMed]
- Ziegler, R.G. Vegetables, fruits, and carotenoids and the risk of cancer. Am. J. Clin. Nutr. 1991, 53, 251S–259S. [Google Scholar] [PubMed]
- Colditz, G.A.; Branch, L.G.; Lipnick, R.J.; Willett, W.C.; Rosner, B.; Posner, B.M.; Hennekens, C.H. Increased green and yellow vegetable intake and lowered cancer deaths in an elderly population. Am. J. Clin. Nutr. 1985, 41, 32–36. [Google Scholar] [PubMed]
- Stähelin, H.B.; Gey, K.F.; Eichholzer, M.; Lüdin, E. Beta-carotene and cancer prevention: The Basel Study. Am. J. Clin. Nutr. 1991, 53, 265S–269S. [Google Scholar] [PubMed]
- Giovannucci, E.; Ascherio, A.; Rimm, E.B.; Stampfer, M.J.; Colditz, G.A.; Willett, W.C. Intake of carotenoids and retino in relation to risk of prostate cancer. J. Natl. Cancer Inst. 1995, 87, 1767–1776. [Google Scholar] [CrossRef] [PubMed]
- Batieha, A.M.; Armenian, H.K.; Norkus, E.P.; Morris, J.S.; Spate, V.E.; Comstock, G.W. Serum micronutrients and the subsequent risk of cervical cancer in a population-based nested case-control study. Cancer Epidemiol. Biomark. Prev. 1993, 2, 335–339. [Google Scholar]
- Blumberg, J. The alpha-tocopherol, beta-carotene cancer prevention study in Finland. Nutr. Rev. 1994, 52, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Omenn, G.S.; Goodman, G.E.; Thornquist, M.D.; Balmes, J.; Cullen, M.R.; Glass, A.; Keogh, J.P.; Meyskens, F.L., Jr.; Valanis, B.; Williams, J.H., Jr.; et al. Risk factors for lung cancer and for intervention effects in CARET, the Beta-Carotene and Retinol Efficacy Trial. J. Natl. Cancer Inst. 1996, 88, 1550–1559. [Google Scholar] [CrossRef] [PubMed]
- Omenn, G.S.; Goodman, G.E.; Thornquist, M.D.; Balmes, J.; Cullen, M.R.; Glass, A.; Keogh, J.P.; Meyskens, F.L.; Valanis, B.; Williams, J.H.; et al. Effects of a combination of beta carotene and vitamin A on lung cancer and cardiovascular disease. N. Engl. J. Med. 1996, 334, 1150–1155. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-D.; Russell, R.M. Procarcinogenic and anticarcinogenic effects of β-carotene. Nutr. Rev. 1999, 57, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Sommerburg, O.; Langhans, C.D.; Arnhold, J.; Leichsenring, M.; Salerno, C.; Crifò, C.; Hoffmann, G.F.; Debatin, K.M.; Siems, W.G. β-Carotene cleavage products after oxidation mediated by hypochlorous acid—A model for neutrophil-derived degradation. Free Radic. Biol. Med. 2003, 35, 1480–1490. [Google Scholar] [CrossRef] [PubMed]
- Siems, W.; Capuozzo, E.; Crifò, C.; Sommerburg, O.; Langhans, C.D.; Schlipalius, L.; Wiswedel, I.; Kraemer, K.; Salerno, C. Carotenoid cleavage products modify respiratory burst and induce apoptosis of human neutrophils. Biochim. Biophys. Acta Mol. Basis Dis. 2003, 1639, 27–33. [Google Scholar] [CrossRef]
- Siems, W.; Sommerburg, O.; Schild, L.; Augustin, W.; Langhans, C.D.; Wiswedel, I. β-Carotene cleavage products induce oxidative stress in vitro by impairing mitochondrial respiration. FASEB J. 2002, 16, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Siems, W.; Wiswedel, I.; Salerno, C.; Crifò, C.; Augustin, W.; Schild, L.; Langhans, C.D.; Sommerburg, O. β-Carotene breakdown products may impair mitochondrial functions—Potential side effects of high-dose β-carotene supplementation. J. Nutr. Biochem. 2005, 16, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Alija, A.J.; Bresgen, N.; Sommerburg, O.; Siems, W.; Eckl, P.M. Cytotoxic and genotoxic effects of β-carotene breakdown products on primary rat hepatocytes. Carcinogenesis 2004, 25, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Alija, A.J.; Bresgen, N.; Sommerburg, O.; Langhans, C.D.; Siems, W.; Eckl, P.M. β-Carotene breakdown products enhance genotoxic effects of oxidative stress in primary rat hepatocytes. Carcinogenesis 2006, 27, 1128–1133. [Google Scholar] [CrossRef] [PubMed]
- Alija, A.J.; Bresgen, N.; Bojaxhi, E.; Vogl, C.; Siems, W.; Eckl, P.M. Cytotoxicity of beta-carotene cleavage products and its prevention by antioxidants. Acta Biochim. Pol. 2010, 57, 217–221. [Google Scholar] [PubMed]
- Yurtcu, E.; Iseri, O.D.; Sahin, F.I. Effects of ascorbic acid and β-carotene on HepG2 human hepatocellular carcinoma cell line. Mol. Boil. Rep. 2011, 38, 4265–4272. [Google Scholar] [CrossRef] [PubMed]
- Castranova, V.; Rabovsky, J.; Tucker, J.H.; Miles, P.R. The alveolar type II epithelial cell: A multifunctional pneumocyte. Toxicol. Appl. Pharmacol. 1988, 93, 472–483. [Google Scholar] [CrossRef]
- De Boeck, M.; Hoet, P.; Lombaert, N.; Nemery, B.; Kirsch-Volders, M.; Lison, D. In vivo genotoxicity of hard metal dust: Induction of micronuclei in rat type II epithelial lung cells. Carcinogenesis 2003, 24, 1793–1800. [Google Scholar] [CrossRef] [PubMed]
- Richards, R.J.; Davies, N.; Atkins, J.; Oreffo, V.I. Isolation, biochemical characterization, and culture of lung type II cells of the rat. Lung 1987, 165, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Kost, D.P.; Michalopoulos, G.K. Effect of 2% dimethyl sulfoxide on the mitogenic properties of epidermal growth factor and hepatocyte growth factor in primary hepatocyte culture. J. Cell. Physiol. 1991, 147, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Cooney, R.V.; Kappock, T.J., 4th; Pung, A.; Bertram, J.S. Solubilization, cellular uptake, and activity of β-carotene and other carotenoids as inhibitors of neoplastic transformation in cultured cells. Methods Enzymol. 1993, 214, 55–68. [Google Scholar] [PubMed]
- Da Violante, G.; Zerrouk, N.; Richard, I.; Provot, G.; Chaumeil, J.C.; Arnaud, P. Evaluation of the cytotoxicity effect of dimethyl sulfoxide (DMSO) on Caco2/TC7 colon tumor cell cultures. Biol. Pharm. Bull. 2002, 25, 1600–1603. [Google Scholar] [CrossRef] [PubMed]
- Eckl, P.; Raffelsberger, I. The primary rat hepatocyte micronucleus assay: General features. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 1997, 392, 117–124. [Google Scholar] [CrossRef]
- Bresgen, N.; Rolinek, R.; Hochleitner, E.; Lottspeich, F.; Eckl, P.M. Induction of apoptosis by a hepatocyte conditioned medium. J. Cell. Physiol. 2004, 198, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Lindl, T.; Bauer, J. Zeil-und Gewebekultur; Gustav Fischer Verlag: Stuttgart, Germany, 1994. [Google Scholar]
- Tice, R.R.; Agurell, E.; Anderson, D.; Burlinson, B.; Hartmann, A.; Kobayashi, H.; Miyamae, Y.; Rojas, E.; Ryu, J.C.; Sasaki, Y.F. Single cell gel/comet assay: Guidelines for in vitro and in vivo genetic toxicology testing. Environ. Mol. Mutagen. 2000, 35, 206–221. [Google Scholar] [CrossRef]
- Burlinson, B.; Tice, R.R.; Speit, G.; Agurell, E.; Brendler-Schwaab, S.Y.; Collins, A.R.; Escobar, P.; Honma, M.; Kumaravel, T.S.; Nakajima, M.; et al. Fourth International Workgroup on Genotoxicity testing: Results of the in vivo Comet assay workgroup. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2007, 627, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Xue, K.-X.; Wu, J.Z.; Ma, G.J.; Yuan, S.; Qin, H.L. Comparative studies on genotoxicity and antigenotoxicity of natural and synthetic β-carotene stereoisomers. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 1998, 418, 73–78. [Google Scholar] [CrossRef]
- Salvadori, D.M.; Ribeiro, L.R.; Oliveira, M.D.; Pereira, C.A.; Beçak, W. The protective effect of β-carotene on genotoxicity induced by cyclophosphamide. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 1992, 265, 237–244. [Google Scholar] [CrossRef]
- Rauscher, R.; Edenharder, R.; Platt, K. In vitro antimutagenic and in vivo anticlastogenic effects of carotenoids and solvent extracts from fruits and vegetables rich in carotenoids. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 1998, 413, 129–142. [Google Scholar] [CrossRef]
- Aidoo, A.; Lyn-Cook, L.E.; Lensing, S.; Bishop, M.E.; Wamer, W. In vivo antimutagenic activity of beta-carotene in rat spleen lymphocytes. Carcinogenesis 1995, 16, 2237–2241. [Google Scholar] [CrossRef] [PubMed]
- Morgan, W.A.; Hartley, J.A.; Cohen, G.M. Quinone-induced DNA single strand breaks in rat hepatocytes and human chronic myelogenous leukaemic K562 cells. Biochem. Pharmacol. 1992, 44, 215–221. [Google Scholar] [CrossRef]
- Stahl, W.; Schwarz, W.; von Laar, J.; Sies, H. All-trans beta-carotene preferentially accumulates in human chylomicrons and very low density lipoproteins compared with the 9-cis geometrical isomer. J. Nutr. 1995, 125, 2128–22133. [Google Scholar] [PubMed]
- Goodman, D.S.; Blomstrand, R.; Werner, B.; Huang, H.S.; Shiratori, T. The intestinal absorption and metabolism of vitamin A and beta-carotene in man. J. Clin. Investig. 1966, 45, 1615–1623. [Google Scholar] [CrossRef] [PubMed]
- Ellsworth, J.; Kraemer, F.; Cooper, A. Transport of beta-very low density lipoproteins and chylomicron remnants by macrophages is mediated by the low density lipoprotein receptor pathway. J. Biol. Chem. 1987, 262, 2316–2325. [Google Scholar] [PubMed]
- Pryor, W.A.; Hales, B.J.; Premovic, P.I.; Church, D.F. The radicals in cigarette tar: Their nature and suggested physiological implications. Science 1983, 220, 425–427. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; MacNee, W. Lung glutathione and oxidative stress: Implications in cigarette smoke-induced airway disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 1999, 277, L1067–L1088. [Google Scholar]
- Elsayed, N.M.; Bendich, A. Dietary antioxidants: Potential effects on oxidative products in cigarette smoke. Nutr. Res. 2001, 21, 551–567. [Google Scholar] [CrossRef]
- Myrvik, Q.N.; Gristina, A.G.; Giridhar, G.; Hayakawa, H. Particle-induced in vivo priming of alveolar macrophages for enhanced oxidative responses: A novel system of cellular immune augmentation. J. Leukoc. Boil. 1993, 54, 439–443. [Google Scholar]
- Kirkham, P.A.; Spooner, G.; Ffoulkes-Jones, C.; Calvez, R. Cigarette smoke triggers macrophage adhesion and activation: Role of lipid peroxidation products and scavenger receptor. Free Radic. Biol. Med. 2003, 35, 697–710. [Google Scholar] [CrossRef]
- Van der Vaart, H.; Postma, D.S.; Timens, W.; ten Hacken, N.H. Acute effects of cigarette smoke on inflammation and oxidative stress: A review. Thorax 2004, 59, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.; Theron, A. Physiological Potential of Ascorbate, β-Carotene and α-Tocopherol Individually and in Combination in the Prevention of Tissue Damage, Carcinogenesis and Immune Dysfunction Mediated by Phagocyte-Derived Reactive Oxidants, in Aspects of Some Vitamins, Minerals and Enzymes in Health and Disease; Karger Publishers: London, UK, 1990; pp. 27–58. [Google Scholar]
- Huie, R.E.; Padmaja, S. The reaction of NO with superoxide. Free Radic. Res. Commun. 1993, 18, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Radi, R.; Peluffo, G.; Alvarez, M.N.; Naviliat, M.; Cayota, A. Unraveling peroxynitrite formation in biological systems. Free Radic. Biol. Med. 2001, 30, 463–488. [Google Scholar] [CrossRef]
- Nierenberg, D.W.; Stukel, T.A.; Baron, J.A.; Dain, B.J.; Greenberg, E.R. Determinants of increase in plasma concentration of beta-carotene after chronic oral supplementation. The Skin Cancer Prevention Study Group. Am. J. Clin. Nutr. 1991, 53, 1443–1449. [Google Scholar] [PubMed]
- Van Helden, Y.G.; Keijer, J.; Knaapen, A.M.; Heil, S.G.; Briedé, J.J.; van Schooten, F.J.; Godschalk, R.W. β-Carotene metabolites enhance inflammation-induced oxidative DNA damage in lung epithelial cells. Free Radic. Biol. Med. 2009, 46, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.M.; Kugelman, A.; Iwamoto, T.; Tian, L.; Forman, H.J. Quinone-induced oxidative stress elevates glutathione and induces gamma-glutamylcysteine synthetase activity in rat lung epithelial L2 cells. J. Biol. Chem. 1994, 269, 26512–26517. [Google Scholar] [PubMed]
- Govindaraju, K.; Govindaraju, V.; Eidelman, D. Analysis of glutathione in rat airway surface liquid by capillary zone electrophoresis with conductivity detection. J. Chromatogr. B 2003, 788, 369–376. [Google Scholar] [CrossRef]
- Iwasaki, Y.; Saito, Y.; Nakano, Y.; Mochizuki, K.; Sakata, O.; Ito, R.; Saito, K.; Nakazawa, H. Chromatographic and mass spectrometric analysis of glutathione in biological samples. J. Chromatogr. B 2009, 877, 3309–3317. [Google Scholar] [CrossRef] [PubMed]
- Van Der Vliet, A.; O’Neill, C.A.; Cross, C.E.; Koostra, J.M.; Volz, W.G.; Halliwell, B.; Louie, S. Determination of low-molecular-mass antioxidant concentrations in human respiratory tract lining fluids. Am. J. Physiol. Lung Cell. Mol. Physiol. 1999, 276, L289–L296. [Google Scholar]
- Esteban-Pretel, G.; López-García, M.P. An experimental design for the controlled modulation of intracellular GSH levels in cultured hepatocytes. Free Radic. Biol. Med. 2006, 41, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Monostori, P.; Wittmann, G.; Karg, E.; Túri, S. Determination of glutathione and glutathione disulfide in biological samples: An in-depth review. J. Chromatogr. B 2009, 877, 3331–3346. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.L.; Morgan, P.E.; Davies, M.J. Quantification of protein modification by oxidants. Free Radic. Biol. Med. 2009, 46, 965–988. [Google Scholar] [CrossRef] [PubMed]
- Siems, W.; Grune, T. Intracellular metabolism of 4-hydroxynonenal. Mol. Asp. Med. 2003, 24, 167–175. [Google Scholar] [CrossRef]
- Bilimoria, M.; Ecobichon, D. Protective antioxidant mechanisms in rat and guinea pig tissues challenged by acute exposure to cigarette smoke. Toxicology 1992, 72, 131–144. [Google Scholar] [CrossRef]
- Park, E.-M.; Park, Y.-M.; Gwak, Y.-S. Oxidative damage in tissues of rats exposed to cigarette smoke. Free Radic. Biol. Med. 1998, 25, 79–86. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Mitotic Cells (%) + | Apoptotic Cells (%) ++ | Necrotic Cells (%) + |
|---|---|---|---|
| Control THF | 2.95 ± 0.43 * | 0.31 ± 0.24 (p = 0.077) 1 | 0.13 ± 0.15 ** |
| Control DMNQ | 2.49 ± 1.16 ** | 0.38 ± 0.19 (p = 0.084) 1 | 0.02 ± 0.04 ** |
| Control 50 µM BC | 2.51 ± 0.89 * | 0.27 ± 0.27 (p = 0.074) 1 | 0.06 ± 0.09 ** |
| 50 µM BC + DMNQ | 0.56 ± 0.12 | 3.91 ± 2.37 | 0.66 ± 0.19 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haider, C.; Ferk, F.; Bojaxhi, E.; Martano, G.; Stutz, H.; Bresgen, N.; Knasmüller, S.; Alija, A.; Eckl, P.M. Effects of β-Carotene and Its Cleavage Products in Primary Pneumocyte Type II Cells. Antioxidants 2017, 6, 37. https://doi.org/10.3390/antiox6020037
Haider C, Ferk F, Bojaxhi E, Martano G, Stutz H, Bresgen N, Knasmüller S, Alija A, Eckl PM. Effects of β-Carotene and Its Cleavage Products in Primary Pneumocyte Type II Cells. Antioxidants. 2017; 6(2):37. https://doi.org/10.3390/antiox6020037
Chicago/Turabian StyleHaider, Cornelia, Franziska Ferk, Ekramije Bojaxhi, Giuseppe Martano, Hanno Stutz, Nikolaus Bresgen, Siegfried Knasmüller, Avdulla Alija, and Peter M. Eckl. 2017. "Effects of β-Carotene and Its Cleavage Products in Primary Pneumocyte Type II Cells" Antioxidants 6, no. 2: 37. https://doi.org/10.3390/antiox6020037
APA StyleHaider, C., Ferk, F., Bojaxhi, E., Martano, G., Stutz, H., Bresgen, N., Knasmüller, S., Alija, A., & Eckl, P. M. (2017). Effects of β-Carotene and Its Cleavage Products in Primary Pneumocyte Type II Cells. Antioxidants, 6(2), 37. https://doi.org/10.3390/antiox6020037