
The Reactive Sulfur Species Concept: 15 Years On




Abstract
:1. Foreword
2. The Importance of Being Yellow
3. Do You Remember the Time?
3.1. A Historical Perspective
3.2. Inspiration from Natural Products
4. The Year 2003: Gestalt Switch in the Field of RSS Research
4.1. Sulfinic Acids in Proteins and Enzymes: Accidental or with (a) Purpose
4.2. New Paradigms: Floodgates, Signals and Omens of Omes to Come
5. What are RSS?
5.1. RSS: Reactive Sulfane Species as Organic Signalling Molecules and Secondary Metabolites
5.2. RSS: Regulatory Sulfur Species Occurring in Proteins and Enzymes
5.3. RSS: Reactive Sulfide Species from Inorganic Chemistry
5.4. RSS: Reducing Sulfur Species with Biological Activity
5.5. RSS: Radical Sulfur Species as Pinnacle of Sulfur Redox Chemistry
6. Food for Thought
6.1. RSS Formed Inside Human Cells
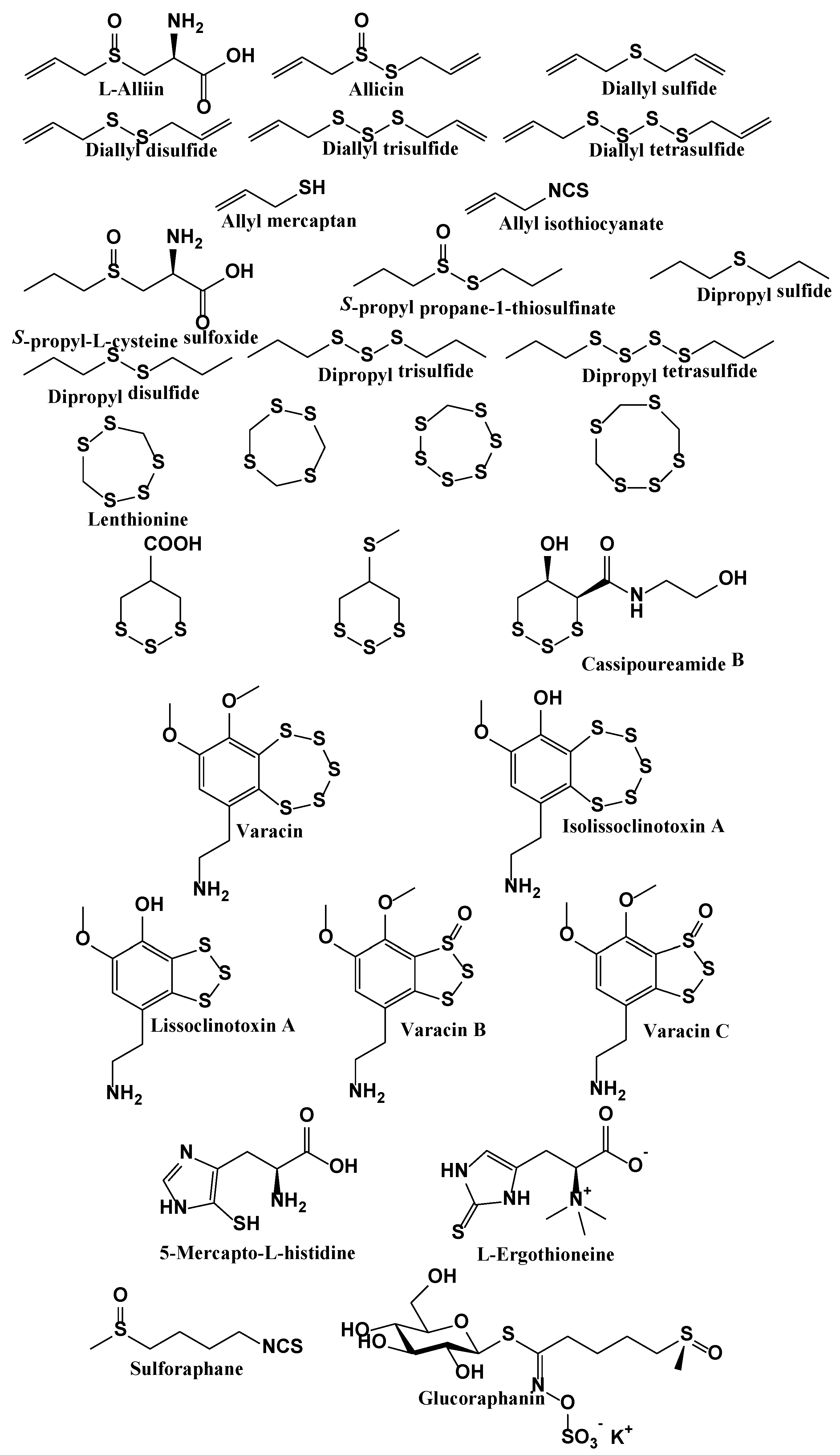
6.2. The Culinary Side of RSS: Organic Sulfur Compounds
6.3. Reprocessed Sulfur Species Produced by the Microbiota
6.4. RSS: Recreational Sulfur Species
7. Towards a Modern Definition of RSS
8. Conclusions and Outlook
8.1. Diversification and Fragmentation
8.2. New Players on the Block
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Giles, G.I.; Tasker, K.M.; Jacob, C. Hypothesis: The role of reactive sulfur species in oxidative stress. Free Radic. Biol. Med. 2001, 31, 1279–1283. [Google Scholar] [CrossRef]
- Kuhn, T.S. The Structure of Scientific Revolutions, 2nd ed.; University of Chicago Press: Chicago, IL, USA, 1970. [Google Scholar]
- Read, R.; Bellew, T.; Yang, J.G.; Hill, K.E.; Palmer, I.S.; Burk, R.F. Selenium and amino acid composition of selenoprotein P, the major selenoprotein in rat serum. J. Biol. Chem. 1990, 265, 17899–17905. [Google Scholar] [PubMed]
- Xu, D.; Rovira, I.I.; Finkel, T. Oxidants painting the cysteine chapel: Redox regulation of PTPs. Dev. Cell. 2002, 2, 251–252. [Google Scholar] [CrossRef]
- Gruhlke, M.C.; Slusarenko, A.J. The biology of reactive sulfur species (RSS). Plant. Physiol. Biochem. 2012, 59, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Giles, G.I.; Jacob, C. Reactive sulfur species: An emerging concept in oxidative stress. Biol. Chem. 2002, 383, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Lin, V.S.; Chen, W.; Xian, M.; Chang, C.J. Chemical probes for molecular imaging and detection of hydrogen sulfide and reactive sulfur species in biological systems. Chem. Soc. Rev. 2015, 44, 4596–4618. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, C.; Savvides, A.; Christou, A.; Fotopoulos, V. Unravelling chemical priming machinery in plants: The role of reactive oxygen-nitrogen-sulfur species in abiotic stress tolerance enhancement. Curr. Opin. Plant. Biol. 2016, 33, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Radi, R.; Trujillo, M. Special issue on “free radical and redox biochemistry of thiols”. Free Radic. Res. 2016, 50, 123–125. [Google Scholar] [CrossRef] [PubMed]
- Schoeneich, C.; Poole, L.B. Special section on redox biology of thiols in signaling pathways. Free Radic. Biol. Med. 2015, 80, 145–212. [Google Scholar]
- Leonard, S.E.; Reddie, K.G.; Carroll, K.S. Mining the thiol proteome for sulfenic acid modifications reveals new targets for oxidation in cells. ACS Chem. Biol. 2009, 4, 783–799. [Google Scholar] [CrossRef] [PubMed]
- Chiappetta, G.; Ndiaye, S.; Igbaria, A.; Kumar, C.; Vinh, J.; Toledano, M.B. Proteome screens for Cys residues oxidation: The redoxome. Methods Enzymol. 2010, 473, 199–216. [Google Scholar] [PubMed]
- Brandes, N.; Reichmann, D.; Tienson, H.; Leichere, L.I.; Jakob, U. Using quantitative redox proteomics to dissect the yeast redoxome. J. Biol. Chem. 2011, 286, 41893–41903. [Google Scholar] [CrossRef] [PubMed]
- Boyum, A.; Forstrom, R.J.; Sefland, I.; Sand, K.L.; Benestad, H.B. Intricacies of redoxome function demonstrated with a simple in vitro chemiluminescence method, with special reference to vitamin B12 as antioxidant. Scand. J. Immunol. 2014, 80, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Jacob, C. Redox signalling via the cellular thiolstat. Biochem. Soc. T 2011, 39, 1247–1253. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Davies, M.J.; Kramer, A.C.; Miotto, G.; Zaccarin, M.; Zhang, H.; Ursini, F. Protein cysteine oxidation in redox signaling: Caveats on sulfenic acid detection and quantification. Arch. Biochem. Biophys. 2017, 617, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant. Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Ha, A.W.; Hong, K.H.; Kim, H.S.; Kim, W.K. Inorganic sulfur reduces cell proliferation by inhibiting of ErbB2 and ErbB3 protein and mRNA expression in MDA-MB-231 human breast cancer cells. Nutr. Res. Pract. 2013, 7, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Devasagayam, T.P.A.; Sundquist, A.R.; Di Mascio, P.; Kaiser, S.; Sies, H. Activity of thiols as singlet molecular-oxygen quenchers. J. Photoch. Photobio. B 1991, 9, 105–116. [Google Scholar] [CrossRef]
- Salmeen, A.; Andersen, J.N.; Myers, M.P.; Meng, T.C.; Hinks, J.A.; Tonks, N.K.; Barford, D. Redox regulation of protein tyrosine phosphatase 1B involves a sulphenyl-amide intermediate. Nature 2003, 423, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Jacob, C.; Giles, G.L.; Giles, N.M.; Sies, H. Sulfur and selenium: The role of oxidation state in protein structure and function. Angew. Chem. Int. Edit 2003, 42, 4742–4758. [Google Scholar] [CrossRef] [PubMed]
- Ghezzi, P. Regulation of protein function by glutathionylation. Free Radical Res. 2005, 39, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Pompella, A.; Visvikis, A.; Paolicchi, A.; De Tata, V.; Casini, A.F. The changing faces of glutathione, a cellular protagonist. Biochem. Pharmacol. 2003, 66, 1499–1503. [Google Scholar] [CrossRef]
- Ghezzi, P. Oxidoreduction of protein thiols in redox regulation. Biochem. Soc. T 2005, 33, 1378–1381. [Google Scholar] [CrossRef]
- Schoneich, C.; Asmus, K.D. Reaction of thiyl radicals with alcohols, ethers and polyunsaturated fatty-acids—A possible role of thiyl free-radicals in thiol mutagenesis. Radiat. Environ. Bioph. 1990, 29, 263–271. [Google Scholar] [CrossRef]
- Akhlaq, M.S.; Schuchmann, H.P.; Vonsonntag, C. The reverse of the repair reaction of thiols: H-abstraction at carbon by thiyl radicals. Int. J. Radiat. Biol. 1987, 51, 91–102. [Google Scholar] [CrossRef]
- Winterbourn, C.C. Are free radicals involved in thiol-based redox signaling? Free Radical Bio. Med. 2015, 80, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Jacob, C. A scent of therapy: Pharmacological implications of natural products containing redox-active sulfur atoms. Nat. Prod. Rep. 2006, 23, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Block, E. The organosulfur chemistry of the genus allium—Implications for the organic chemistry of sulfur. Angew. Chem.-Int. Edit. 1992, 31, 1135–1178. [Google Scholar] [CrossRef]
- Schroder, E.; Littlechild, J.A.; Lebedev, A.A.; Errington, N.; Vagin, A.A.; Isupov, M.N. Crystal structure of decameric 2-Cys peroxiredoxin from human erythrocytes at 1.7 angstrom resolution. Struct. Fold. Des. 2000, 8, 605–615. [Google Scholar] [CrossRef]
- Harris, J.R.; Schroder, E.; Isupov, M.N.; Scheffler, D.; Kristensen, P.; Littlechild, J.A.; Vagin, A.A.; Meissner, U. Comparison of the decameric structure of peroxiredoxin-ii by transmission electron microscopy and X-ray crystallography. Bba.-Protein. Struct. M 2001, 1547, 221–234. [Google Scholar] [CrossRef]
- Biteau, B.; Labarre, J.; Toledano, M.B. ATP-dependent reduction of cysteine-sulphinic acid by S. cerevisiae sulphiredoxin. Nature 2003, 425, 980–984. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.; Park, S.J.; Chang, T.S.; Lee, D.Y.; Rhee, S.G. Molecular mechanism of the reduction of cysteine sulfinic acid of peroxiredoxin to cysteine by mammalian sulfiredoxin. J. Biol. Chem. 2006, 281, 14400–14407. [Google Scholar] [CrossRef] [PubMed]
- Canet-Aviles, R.M.; Wilson, M.A.; Miller, D.W.; Ahmad, R.; McLendon, C.; Bandyopadhyay, S.; Baptista, M.J.; Ringe, D.; Petsko, G.A.; Cookson, M.R. The Parkinson's disease protein DJ-1 is neuroprotective due to cysteine-sulfinic acid-driven mitochondrial localization. Proc. Natl. Acad. Sci. USA 2004, 101, 9103–9108. [Google Scholar] [CrossRef] [PubMed]
- Jacob, C.; Holme, A.L.; Fry, F.H. The sulfinic acid switch in proteins. Org. Biomol. Chem. 2004, 2, 1953–1956. [Google Scholar] [CrossRef] [PubMed]
- Allah, D.R.; Schwind, L.; Asali, I.A.; Nasim, J.; Jacob, C.; Gotz, C.; Montenarh, M. A scent of therapy: Synthetic polysulfanes with improved physico-chemical properties induce apoptosis in human cancer cells. Int. J. Oncol. 2015, 47, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Saidu, N.E.B.; Intemann, J.; Jacob, C.; Montenarh, M. A new tellurium-containing amphiphilic molecule induces apoptosis in HCT116 colon cancer cells. Bba.-Gen. Subjects 2014, 1840, 1808–1816. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Viswanathan, U.M.; Khairan, K.; Buric, T.; Saidu, N.E.B.; Xu, Z.J.; Hanf, B.; Bazukyan, I.; Trchounian, A.; Hannemann, F.; et al. Synthesis of amphiphilic, chalcogen-based redox modulators with in vitro cytotoxic activity against cancer cells, macrophages and microbes. Medchemcomm 2014, 5, 25–31. [Google Scholar] [CrossRef]
- Saidu, N.E.B.; Asali, I.A.; Czepukojc, B.; Seitz, B.; Jacob, C.; Montenarh, M. Comparison between the effects of diallyl tetrasulfide on human retina pigment epithelial cells (ARPE-19) and HCT116 cells. Bba.-Gen. Subjects 2013, 1830, 5267–5276. [Google Scholar] [CrossRef] [PubMed]
- Saidu, N.E.B.; Touma, R.; Asali, I.A.; Jacob, C.; Montenarh, M. Diallyl tetrasulfane activates both the eIF2 alpha and Nrf2/HO-1 pathways. Bba.-Gen. Subjects 2013, 1830, 2214–2225. [Google Scholar] [CrossRef] [PubMed]
- Wood, Z.A.; Poole, L.B.; Karplus, P.A. Peroxiredoxin evolution and the regulation of hydrogen peroxide signaling. Science 2003, 300, 650–653. [Google Scholar] [CrossRef] [PubMed]
- Poole, L.B.; Karplus, P.A.; Claiborne, A. Protein sulfenic acids in redox signaling. Annu. Rev. Pharmacol. 2004, 44, 325–347. [Google Scholar] [CrossRef] [PubMed]
- Roberts, B.R.; Wood, Z.A.; Jonsson, T.J.; Poole, L.B.; Karplus, P.A. Oxidized and synchrotron cleaved structures of the disulfide redox center in the N-terminal domain of salmonella typhimurium AhpF. Protein Sci. 2005, 14, 2414–2420. [Google Scholar] [CrossRef] [PubMed]
- Cording, J.; Gunther, R.; Vigolo, E.; Tscheik, C.; Winkler, L.; Schlattner, I.; Lorenz, D.; Haseloff, R.F.; Schmidt-Ott, K.M.; Wolburg, H.; et al. Redox regulation of cell contacts by tricellulin and occludin: Redox-sensitive cysteine sites in tricellulin regulate both tri- and bicellular junctions in tissue barriers as shown in hypoxia and ischemia. Antioxid. Redox Sign 2015, 23, 1035–1049. [Google Scholar] [CrossRef] [PubMed]
- Grek, C.L.; Zhang, J.; Manevich, Y.; Townsend, D.M.; Tew, K.D. Causes and consequences of cysteine S-glutathionylation. J. Bio. Chem. 2013, 288, 26497–26504. [Google Scholar] [CrossRef] [PubMed]
- Ashby, M.T.; Aneetha, H. Reactive sulfur species: Aqueous chemistry of sulfenyl thiocyanates. J. Am. Chem. Soc. 2004, 126, 10216–10217. [Google Scholar] [CrossRef] [PubMed]
- Mishanina, A.V.; Libiad, M.; Banerjee, R. Biogenesis of reactive sulfur species for signaling by hydrogen sulfide oxidation pathways. Nat. Chem. Biol. 2015, 11, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Toohey, J.I.; Cooper, A.J.L. Thiosulfoxide (sulfane) sulfur: New chemistry and new regulatory roles in biology. Molecules 2014, 19, 12789–12813. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, L.R.; Francom, D.; Dieken, F.P.; Taylor, J.D.; Warenycia, M.W.; Reiffenstein, R.J.; Dowling, G. Determination of sulfide in brain-tissue by gas dialysis ion chromatography: Postmortem studies and 2 case-reports. J. Anal. Toxicol. 1989, 13, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, R.O.; Bus, J.S.; Popp, J.A.; Boreiko, C.J.; Andjelkovich, D.A. A critical review of the literature on hydrogen sulfide toxicity. Crit. Rev. Toxicol. 1984, 13, 25–97. [Google Scholar] [CrossRef] [PubMed]
- Abe, K.; Kimura, H. The possible role of hydrogen sulfide as an endogenous neuromodulator. J. Neurosci. 1996, 16, 1066–1071. [Google Scholar] [PubMed]
- Doeller, J.E.; Isbell, T.S.; Benavides, G.; Koenitzer, J.; Patel, H.; Patel, R.P.; Lancaster, J.R.; Darley-Usmar, V.M.; Kraus, D.W. Polarographic measurement of hydrogen sulfide production and consumption by mammalian tissues. Anal. Biochem. 2005, 341, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Dello, R.C.; Tringali, G.; Ragazzoni, E.; Maggiano, N.; Menini, E.; Vairano, M.; Preziosi, P.; Navarra, P. Evidence that hydrogen sulphide can modulate hypothalamo-pituitary-adrenal axis function: In vitro and in vivo studies in the rat. J. Neuroendocrinol. 2000, 12, 225–233. [Google Scholar] [CrossRef]
- Eto, K.; Ogasawara, M.; Umemura, K.; Nagai, Y.; Kimura, H. Hydrogen sulfide is produced in response to neuronal excitation. J. Neurosci. 2002, 22, 3386–3391. [Google Scholar] [PubMed]
- Hosoki, R.; Matsuki, N.; Kimura, H. The possible role of hydrogen sulfide as an endogenous smooth muscle relaxant in synergy with nitric oxide. Biochem. Biophys. Res. Commun. 1997, 237, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhang, J.; Lu, Y.; Wang, R. The vasorelaxant effect of H2S as a novel endogenous gaseous KATP channel opener. EMBO J. 2001, 20, 6008–6016. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Bhatia, M.; Zhu, Y.Z.; Zhu, Y.C.; Ramnath, R.D.; Wang, Z.J.; Anuar, F.B.; Whiteman, M.; Salto-Tellez, M.; Moore, P.K. Hydrogen sulfide is a novel mediator of lipopolysaccharide-induced inflammation in the mouse. Faseb. J. 2005, 19, 1196–1198. [Google Scholar] [CrossRef] [PubMed]
- Ang, A.D.; Konigstorfer, A.; Giles, G.I.; Bhatia, M. Measuring free tissue sulfide. Adv. Biol. Chem. 2012, 2, 360–365. [Google Scholar] [CrossRef]
- Finley, J.W.; Wheeler, E.L.; Witt, S.C. Oxidation of glutathione by hydrogen peroxide and other oxidizing agents. J. Agric. Food Chem. 1981, 29, 404–407. [Google Scholar] [CrossRef] [PubMed]
- Saurin, A.T.; Neubert, H.; Brennan, J.P.; Eaton, P. Widespread sulfenic acid formation in tissues in response to hydrogen peroxide. Proc. Natl. Acad. Sci. USA 2004, 101, 17982–17987. [Google Scholar] [CrossRef] [PubMed]
- Waszczak, C.; Akter, S.; Eeckhout, D.; Persiau, G.; Wahni, K.; Bodra, N.; Van Molle, I.; De Smet, B.; Vertommen, D.; Gevaert, K.; et al. Sulfenome mining in arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2014, 111, 11545–11550. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Carroll, K.S. Sulfenic acid chemistry, detection and cellular lifetime. Biochim. Biophys. Acta 2014, 1840, 847–875. [Google Scholar] [CrossRef] [PubMed]
- Aroca, A.; Serna, A.; Gotor, C.; Romero, L.C. S-sulfhydration: A cysteine posttranslational modification in plant systems. Plant. Physiol. 2015, 168, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Doka, E.; Pader, I.; Biro, A.; Johansson, K.; Cheng, Q.; Ballago, K.; Prigge, J.R.; Pastor-Flores, D.; Dick, T.P.; Schmidt, E.E.; et al. A novel persulfide detection method reveals protein persulfide- and polysulfide-reducing functions of thioredoxin and glutathione systems. Sci. Adv. 2016, 2, e1500968. [Google Scholar] [CrossRef] [PubMed]
- Drazic, A.; Winter, J. The physiological role of reversible methionine oxidation. Biochim. Biophys. Acta 2014, 1844, 1367–1382. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Weiss, S.J.; Levine, R.L. Methionine oxidation and reduction in proteins. Biochim. Biophys. Acta 2014, 1840, 901–905. [Google Scholar] [CrossRef] [PubMed]
- Jacques, S.; Ghesquiere, B.; De Bock, P.J.; Demol, H.; Wahni, K.; Willems, P.; Messens, J.; Van Breusegem, F.; Gevaert, K. Protein methionine sulfoxide dynamics in arabidopsis thaliana under oxidative stress. Mol. Cell. Proteom. 2015, 14, 1217–1229. [Google Scholar] [CrossRef] [PubMed]
- Schwarzlander, M.; Wagner, S.; Ermakova, Y.G.; Belousov, V.V.; Radi, R.; Beckman, J.S.; Buettner, G.R.; Demaurex, N.; Duchen, M.R.; Forman, H.J.; et al. The “mitoflash” probe cpYFP does not respond to superoxide. Nature 2014, 514, E12–E14. [Google Scholar] [CrossRef] [PubMed]
- Shen, E.Z.; Song, C.Q.; Lin, Y.; Zhang, W.H.; Su, P.F.; Liu, W.Y.; Zhang, P.; Xu, J.; Lin, N.; Zhan, C.; et al. Mitoflash frequency in early adulthood predicts lifespan in caenorhabditis elegans. Nature 2014, 508, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Wang, W.; Wang, X.; Sheu, S.S.; Dirksen, R.T.; Dong, M.Q. Cheng et al. Reply. Nature 2014, 514, E14–E15. [Google Scholar] [CrossRef] [PubMed]
- Schwarzlander, M.; Dick, T.P.; Meyer, A.J.; Morgan, B. Dissecting redox biology using fluorescent protein sensors. Antioxid. Redox Signal. 2016, 24, 680–712. [Google Scholar] [CrossRef] [PubMed]
- DeLeon, E.R.; Gao, Y.; Huang, E.; Arif, M.; Arora, N.; Divietro, A.; Patel, S.; Olson, K.R. A case of mistaken identity: Are reactive oxygen species actually reactive sulfide species? Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R549–R560. [Google Scholar] [CrossRef] [PubMed]
- Lemma, K.; Ashby, M.T. Reactive sulfur species: Kinetics and mechanism of the reaction of hypothiocyanous acid with cyanide to give dicyanosulfide in aqueous solution. Chem. Res. Toxicol. 2009, 22, 1622–1628. [Google Scholar] [CrossRef] [PubMed]
- Lemma, K.; Ashby, M.T. Reactive sulfur species: Kinetics and mechanism of the equilibrium between cysteine sulfenyl thiocyanate and cysteine thiosulfinate ester in acidic aqueous solution. J. Org. Chem. 2008, 73, 3017–3023. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.; Wang, X.; Lemma, K.; Ashby, M.T. Reactive sulfur species: Hydrolysis of hypothiocyanite to give thiocarbamate-S-oxide. J. Am. Chem. Soc. 2007, 129, 15756–15757. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.; Lemma, K.; Ashby, M.T. Reactive sulfur species: Kinetics and mechanisms of the reaction of cysteine thiosulfinate ester with cysteine to give cysteine sulfenic acid. J. Org. Chem. 2007, 72, 8838–8846. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.; Ashby, M.T. Reactive sulfur species: Kinetics and mechanisms of the oxidation of cysteine by hypohalous acid to give cysteine sulfenic acid. J. Am. Chem. Soc. 2007, 129, 14082–14091. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.; Ashby, M.T. Reactive sulfur species: Kinetics and mechanism of the hydrolysis of cysteine thiosulfinate ester. Chem. Res. Toxicol. 2007, 20, 1364–1372. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.; Ashby, M.T. Reactive sulfur species: Kinetics and mechanism of the oxidation of cystine by hypochlorous acid to give N,N’-dichlorocystine. Chem. Res. Toxicol. 2005, 18, 919–923. [Google Scholar] [CrossRef] [PubMed]
- Ueki, H.; Chapman, G.; Ashby, M.T. Reactive sulfur species: Kinetics and mechanism of the oxidation of aryl sulfinates with hypochlorous acid. J. Phys. Chem. A 2010, 114, 1670–1676. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ashby, M.T. Reactive sulfur species: Kinetics and mechanism of the reaction of thiocarbamate-S-oxide with cysteine. Chem. Res. Toxicol. 2008, 21, 2120–2126. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, M.; Li, L.; Kostetski, I.; Chu, S.H.; Siau, J.L.; Bhatia, M.; Moore, P.K. Evidence for the formation of a novel nitrosothiol from the gaseous mediators nitric oxide and hydrogen sulphide. Biochem. Biophys. Res. Commun. 2006, 343, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Oliva, S.; Herrera, B.; Toro-Labbe, A.; Chermette, H. On the mechanism of hydrogen transfer in the HSCH(O) ⇌ (S)CHOH and HSNO ⇌ SNOH reactions. J. Phys. Chem. A 2005, 109, 1748–1751. [Google Scholar] [CrossRef] [PubMed]
- Filipovic, M.R.; Miljkovic, J.; Nauser, T.; Royzen, M.; Klos, K.; Shubina, T.; Koppenol, W.H.; Lippard, S.J.; Ivanovic-Burmazovic, I. Chemical characterization of the smallest S-nitrosothiol, HSNO; cellular cross-talk of H2S and S-nitrosothiols. J. Am. Chem. Soc. 2012, 134, 12016–12027. [Google Scholar] [CrossRef] [PubMed]
- Miljkovic, J.; Kenkel, I.; Ivanovic-Burmazovic, I.; Filipovic, M.R. Generation of HNO and HSNO from nitrite by heme-iron-catalyzed metabolism with H2S. Angew. Chem. Int. Ed. Engl. 2013, 52, 12061–12064. [Google Scholar] [CrossRef] [PubMed]
- Cortese-Krott, M.M.; Butler, A.R.; Woollins, J.D.; Feelisch, M. Inorganic sulfur-nitrogen compounds: From gunpowder chemistry to the forefront of biological signaling. Dalton Trans. 2016, 45, 5908–5919. [Google Scholar] [CrossRef] [PubMed]
- Wedmann, R.; Zahl, A.; Shubina, T.E.; Durr, M.; Heinemann, F.W.; Bugenhagen, B.E.; Burger, P.; Ivanovic-Burmazovic, I.; Filipovic, M.R. Does perthionitrite (SSNO−) account for sustained bioactivity of NO? A (bio)chemical characterization. Inorg. Chem. 2015, 54, 9367–9380. [Google Scholar] [CrossRef] [PubMed]
- Berenyiova, A.; Grman, M.; Mijuskovic, A.; Stasko, A.; Misak, A.; Nagy, P.; Ondriasova, E.; Cacanyiova, S.; Brezova, V.; Feelisch, M.; et al. The reaction products of sulfide and S-nitrosoglutathione are potent vasorelaxants. Nitric Oxide 2015, 46, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, L.V.; Anton, B.J.; Timerghazin, Q.K. On the possible biological relevance of HSNO isomers: A computational investigation. Phys. Chem. Chem. Phys. 2014, 16, 8476–8486. [Google Scholar] [CrossRef] [PubMed]
- Cortese-Krott, M.M.; Fernandez, B.O.; Santos, J.L.; Mergia, E.; Grman, M.; Nagy, P.; Kelm, M.; Butler, A.; Feelisch, M. Nitrosopersulfide (SSNO−) accounts for sustained NO bioactivity of S-nitrosothiols following reaction with sulfide. Redox Biol. 2014, 2, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Grman, M.; Misak, A.; Jacob, C.; Tomaskova, Z.; Bertova, A.; Burkholz, T.; Docolomansky, P.; Habala, L.; Ondrias, K. Low molecular thiols, pH and O2 modulate H2S-induced S-nitrosoglutathione decomposition—•NO release. Gen. Physiol. Biophys. 2013, 32, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Hydrogen sulfide and polysulfides as signaling molecules. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2015, 91, 131–159. [Google Scholar] [CrossRef] [PubMed]
- Ida, T.; Sawa, T.; Ihara, H.; Tsuchiya, Y.; Watanabe, Y.; Kumagai, Y.; Suematsu, M.; Motohashi, H.; Fujii, S.; Matsunaga, T.; et al. Reactive cysteine persulfides and S-polythiolation regulate oxidative stress and redox signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 7606–7611. [Google Scholar] [CrossRef] [PubMed]
- Greiner, R.; Palinkas, Z.; Basell, K.; Becher, D.; Antelmann, H.; Nagy, P.; Dick, T.P. Polysulfides link H2S to protein thiol oxidation. Antioxid. Redox Signal. 2013, 19, 1749–1765. [Google Scholar] [CrossRef] [PubMed]
- Munchberg, U.; Anwar, A.; Mecklenburg, S.; Jacob, C. Polysulfides as biologically active ingredients of garlic. Org. Biomol. Chem. 2007, 5, 1505–1518. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Physiological roles of hydrogen sulfide and polysulfides. Handb. Exp. Pharmacol. 2015, 230, 61–81. [Google Scholar] [PubMed]
- Beltowski, J. Hydrogen sulfide in pharmacology and medicine—An update. Pharmacol. Rep. 2015, 67, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Mueller, E.G. Trafficking in persulfides: Delivering sulfur in biosynthetic pathways. Nat. Chem. Biol. 2006, 2, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Mitsuyama, H.; May, J.M. Uptake and antioxidant effects of ergothioneine in human erythrocytes. Clin. Sci. (Lond.) 1999, 97, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Grundemann, D.; Harlfinger, S.; Golz, S.; Geerts, A.; Lazar, A.; Berkels, R.; Jung, N.; Rubbert, A.; Schomig, E. Discovery of the ergothioneine transporter. Proc. Natl. Acad. Sci. USA 2005, 102, 5256–5261. [Google Scholar] [CrossRef] [PubMed]
- Borlinghaus, J.; Albrecht, F.; Gruhlke, M.C.; Nwachukwu, I.D.; Slusarenko, A.J. Allicin: Chemistry and biological properties. Molecules 2014, 19, 12591–12618. [Google Scholar] [CrossRef] [PubMed]
- Fry, F.H.; Okarter, N.; Baynton-Smith, C.; Kershaw, M.J.; Talbot, N.J.; Jacob, C. Use of a substrate/alliinase combination to generate antifungal activity in situ. J. Agric. Food Chem. 2005, 53, 574–580. [Google Scholar] [CrossRef] [PubMed]
- Estevam, E.C.; Griffin, S.; Nasim, M.J.; Zielinski, D.; Aszyk, J.; Osowicka, M.; Dawidowska, N.; Idroes, R.; Bartoszek, A.; Jacob, C. Inspired by nature: The use of plant-derived substrate/enzyme combinations to generate antimicrobial activity in situ. Nat. Prod. Commun. 2015, 10, 1733–1738. [Google Scholar] [PubMed]
- Gruhlke, M.C.; Hemmis, B.; Noll, U.; Wagner, R.; Luhring, H.; Slusarenko, A.J. The defense substance allicin from garlic permeabilizes membranes of beta vulgaris, rhoeo discolor, chara corallina and artificial lipid bilayers. Biochim. Biophys. Acta 2015, 1850, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, C.G.; Rother, D.; Bardischewsky, F.; Quentmeier, A.; Fischer, J. Oxidation of reduced inorganic sulfur compounds by bacteria: Emergence of a common mechanism? Appl. Environ. Microbiol. 2001, 67, 2873–2882. [Google Scholar] [CrossRef] [PubMed]
- Kabil, O.; Vitvitsky, V.; Banerjee, R. Sulfur as a signaling nutrient through hydrogen sulfide. Annu. Rev. Nutr. 2014, 34, 171–205. [Google Scholar] [CrossRef] [PubMed]
- Ankri, S.; Mirelman, D. Antimicrobial properties of allicin from garlic. Microbes Infect. 1999, 1, 125–129. [Google Scholar] [CrossRef]
- Czepukojc, B.; Leroch, M.; Salm, F.; Viswanathan, U.M.; Burkholz, T.; Hahn, M.; Jacob, C. Antifungal activity of tetrasulfanes against botrytis cinerea. Nat. Prod. Commun. 2013, 8, 1599–1603. [Google Scholar] [PubMed]
- Czepukojc, B.; Viswanathan, U.M.; Raza, A.; Ali, S.; Burkholz, T.; Jacob, C. Tetrasulfanes as selective modulators of the cellular thiolstat. Phosphorus Sulfur Silicon Relat. Elem. 2013, 188, 446–453. [Google Scholar] [CrossRef]
- Stroot, P.G. The primary cause of oxidative stress is ultra-exogenous sulfide formation (USF). Med. Hypotheses 2014, 83, 766–768. [Google Scholar] [CrossRef] [PubMed]
- Genuine Haarlem Oil. Available online: https://www.haarlem-oil.com/ (accessed on 17 April 2017).
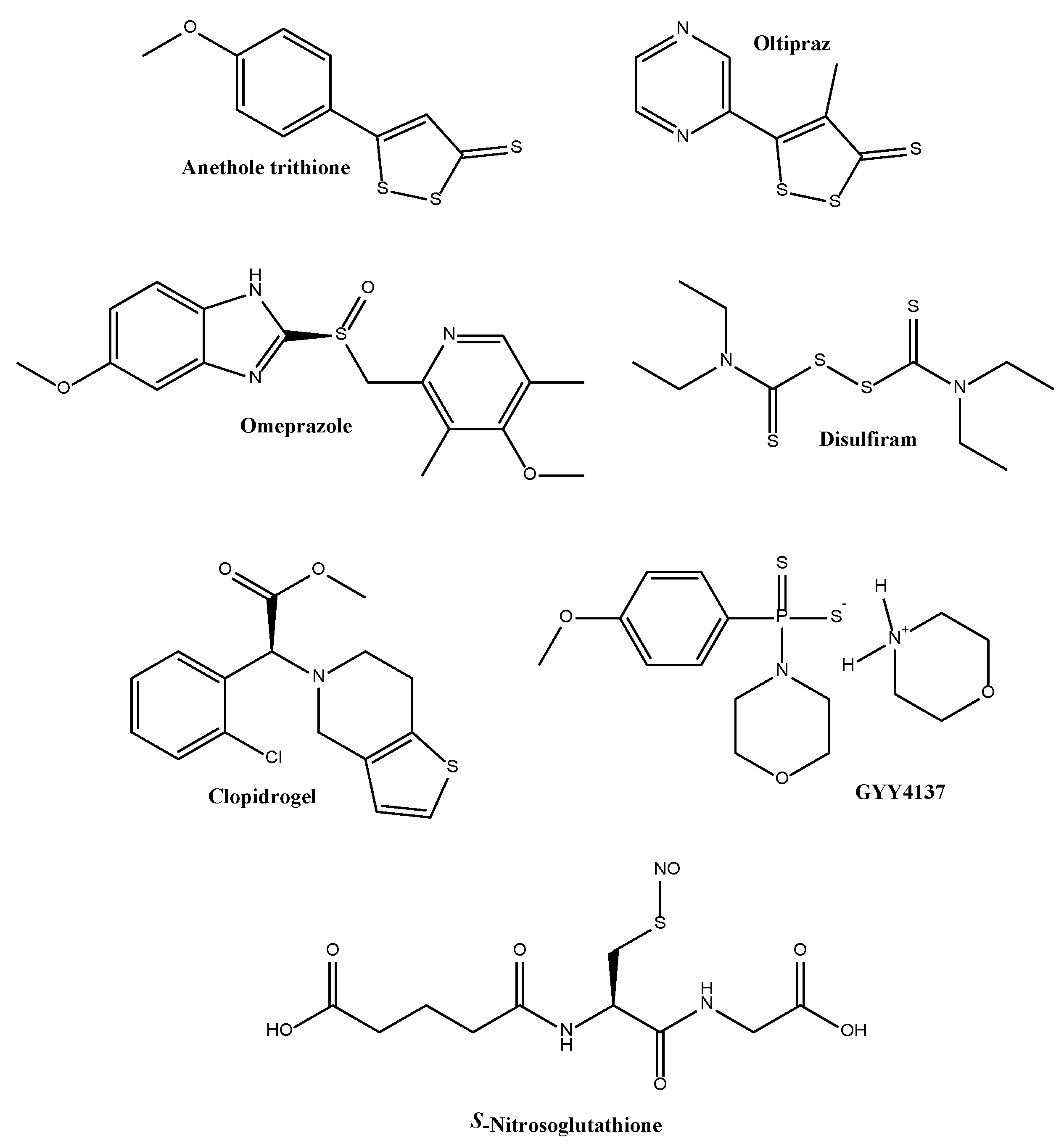
- Zhang, S.; Patel, A.; Moorthy, B.; Shivanna, B. Omeprazole induces NAD(P)H quinone oxidoreductase 1 via aryl hydrocarbon receptor-independent mechanisms: Role of the transcription factor nuclear factor erythroid 2-related factor 2. Biochem. Biophys. Res. Commun. 2015, 467, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Shinkai, Y.; Abiko, Y.; Ida, T.; Miura, T.; Kakehashi, H.; Ishii, I.; Nishida, M.; Sawa, T.; Akaike, T.; Kumagai, Y. Reactive sulfur species-mediated activation of the keap1-Nrf2 pathway by 1,2-naphthoquinone through sulfenic acids formation under oxidative stress. Chem. Res. Toxicol. 2015, 28, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Qi, H.; Shen, Q.; Lu, G.; Li, M.; Bao, K.; Wu, Y.; Zhang, W. 4,5-Diaryl-3H-1,2-dithiole-3-thiones and related compounds as combretastatin A-4/oltipraz hybrids: Synthesis, molecular modelling and evaluation as antiproliferative agents and inhibitors of tubulin. Eur. J. Med. Chem. 2016, 122, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Gruhlke, M.C.; Nicco, C.; Batteux, F.; Slusarenko, A.J. The effects of allicin, a reactive sulfur species from garlic, on a selection of mammalian cell lines. Antioxidants 2016, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Zhang, Y.; Paonessa, J.D.; Munday, C.M.; Wilkins, A.L.; Babu, J. Synthesis, biological evaluation, and structure-activity relationships of dithiolethiones as inducers of cytoprotective phase 2 enzymes. J. Med. Chem. 2010, 53, 4761–4767. [Google Scholar] [CrossRef] [PubMed]
- Tazzari, V.; Cappelletti, G.; Casagrande, M.; Perrino, E.; Renzi, L.; Del Soldato, P.; Sparatore, A. New aryldithiolethione derivatives as potent histone deacetylase inhibitors. Bioorg. Med. Chem. 2010, 18, 4187–4194. [Google Scholar] [CrossRef] [PubMed]
- Marut, W.; Jamier, V.; Kavian, N.; Servettaz, A.; Winyard, P.G.; Eggleton, P.; Anwar, A.; Nicco, C.; Jacob, C.; Chereau, C.; et al. The natural organosulfur compound dipropyltetrasulfide prevents HOCl-induced systemic sclerosis in the mouse. Arthritis Res. Ther. 2013, 15, R167. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Whiteman, M.; Guan, Y.Y.; Neo, K.L.; Cheng, Y.; Lee, S.W.; Zhao, Y.; Baskar, R.; Tan, C.H.; Moore, P.K. Characterization of a novel, water-soluble hydrogen sulfide-releasing molecule (GYY4137): New insights into the biology of hydrogen sulfide. Circulation 2008, 117, 2351–2360. [Google Scholar] [CrossRef] [PubMed]
- Caliendo, G.; Cirino, G.; Santagada, V.; Wallace, J.L. Synthesis and biological effects of hydrogen sulfide (H2S): Development of H2S-releasing drugs as pharmaceuticals. J. Med. Chem. 2010, 53, 6275–6286. [Google Scholar] [CrossRef] [PubMed]
- Brannan, R.G. Reactive sulfur species act as prooxidants in liposomal and skeletal muscle model systems. J. Agric. Food Chem. 2010, 58, 3767–3771. [Google Scholar] [CrossRef] [PubMed]
- Reactive Sulfur Species. Available online: https://en.wikipedia.org/wiki/Reactive_sulfur_species (Accessed on 17 April 2017).
- Jones, D.P.; Sies, H. The redox code. Antioxid. Redox Signal. 2015, 23, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.; Dymock, B.W.; Moore, P.K. Gyy4137, a novel water-soluble, H2S-releasing molecule. Methods Enzymol. 2015, 554, 143–167. [Google Scholar] [PubMed]
- Hellmich, M.R.; Coletta, C.; Chao, C.; Szabo, C. The therapeutic potential of cystathionine beta-synthetase/hydrogen sulfide inhibition in cancer. Antioxid. Redox Signal. 2015, 22, 424–448. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Yamashita, Y.; Suzuki, T.; Kani, Y.; Mizusawa, N.; Imamura, S.; Takemoto, K.; Hara, T.; Hossain, M.A.; Yabu, T.; et al. Selenoneine, a novel selenium-containing compound, mediates detoxification mechanisms against methylmercury accumulation and toxicity in zebrafish embryo. Mar. Biotechnol. 2013, 15, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.; Yabu, T.; Yamashita, M. Discovery of the strong antioxidant selenoneine in tuna and selenium redox metabolism. World J. Biol. Chem. 2010, 1, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.; Yamashita, M. Identification of a novel selenium-containing compound, selenoneine, as the predominant chemical form of organic selenium in the blood of bluefin tuna. J. Biol. Chem. 2010, 285, 18134–18138. [Google Scholar] [CrossRef] [PubMed]
- Estevam, E.C.; Witek, K.; Faulstich, L.; Nasim, M.J.; Latacz, G.; Dominguez-Alvarez, E.; Kiec-Kononowicz, K.; Demasi, M.; Handzlik, J.; Jacob, C. Aspects of a distinct cytotoxicity of selenium salts and organic selenides in living cells with possible implications for drug design. Molecules 2015, 20, 13894–13912. [Google Scholar] [CrossRef] [PubMed]
- Estevam, E.C.; Griffin, S.; Nasim, M.J.; Denezhkin, P.; Schneider, R.; Lilischkis, R.; Dominguez-Alvarez, E.; Witek, K.; Latacz, G.; Keck, C.; et al. Natural selenium particles from staphylococcus carnosus: Hazards or particles with particular promise? J. Hazard. Mater. 2017, 324, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Viswanathan, U.M.; Xu, Z.; Ebrahimnejad, H.; Hanf, B.; Burkholz, T.; Schneider, M.; Bernhardt, I.; Kirsch, G.; Jacob, C. Synthesis of amphiphilic seleninic acid derivatives with considerable activity against cellular membranes and certain pathogenic microbes. J. Hazard. Mater. 2014, 269, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Bhabak, K.P.; Satheeshkumar, K.; Jayavelu, S.; Mugesh, G. Inhibition of peroxynitrite- and peroxidase-mediated protein tyrosine nitration by imidazole-based thiourea and selenourea derivatives. Org. Biomol. Chem. 2011, 9, 7343–7350. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.K.; Koketsu, M.; Lee, K.; Choi, S.Y.; Park, J.H.; Ishihara, H.; Kim, S.Y. Inhibition of tyrosinase activity by N,N-unsubstituted selenourea derivatives. Biol. Pharm Bull. 2005, 28, 838–840. [Google Scholar] [CrossRef] [PubMed]
- Arsenyan, P.; Vasiljeva, J.; Belyakov, S.; Liepinsh, E.; Petrova, M. Fused selenazolinium salt derivatives with a Se–N+ bond: Preparation and properties. Eur. J. Org. Chem. 2015, 2015, 5842–5855. [Google Scholar] [CrossRef]
- Sivapriya, K.; Suguna, P.; Banerjee, A.; Saravanan, V.; Rao, D.N.; Chandrasekaran, S. Facile one-pot synthesis of thio and selenourea derivatives: A new class of potent urease inhibitors. Bioorg. Med. Chem. Lett. 2007, 17, 6387–6391. [Google Scholar] [CrossRef] [PubMed]
- Azad, G.K.; Tomar, R.S. Ebselen, a promising antioxidant drug: Mechanisms of action and targets of biological pathways. Mol. Biol. Rep. 2014, 41, 4865–4879. [Google Scholar] [CrossRef] [PubMed]
- Jackson-Rosario, S.E.; Self, W.T. Targeting selenium metabolism and selenoproteins: Novel avenues for drug discovery. Metallomics. 2010, 2, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Mihara, H. Discovery and molecular function analysis of sulfur- and selenium-eliminating enzymes crucial for biosynthesis of iron-sulfur clusters and selenoproteins. Seikagaku. 2011, 83, 1003–1015. [Google Scholar] [PubMed]
- Crich, D.; Krishnamurthy, V.; Brebion, F.; Karatholuvhu, M.; Subramanian, V.; Hutton, T.K. Dechalcogenative allylic selenosulfide and disulfide rearrangements: Complementary methods for the formation of allylic sulfides in the absence of electrophiles. Scope, limitations, and application to the functionalization of unprotected peptides in aqueous media. J. Am. Chem. Soc. 2007, 129, 10282–10294. [Google Scholar] [PubMed]
- Crich, D.; Krishnamurthy, V.; Hutton, T.K. Allylic selenosulfide rearrangement: A method for chemical ligation to cysteine and other thiols. J. Am. Chem. Soc. 2006, 128, 2544–2545. [Google Scholar] [CrossRef] [PubMed]
- Besse, D.; Moroder, L. Synthesis of selenocysteine peptides and their oxidation to diselenide-bridged compounds. J. Pept. Sci. 1997, 3, 442–453. [Google Scholar] [CrossRef]
- Nascimento, V.; Alberto, E.E.; Tondo, D.W.; Dambrowski, D.; Detty, M.R.; Nome, F.; Braga, A.L. Gpx-like activity of selenides and selenoxides: Experimental evidence for the involvement of hydroxy perhydroxy selenane as the active species. J. Am. Chem. Soc. 2012, 134, 138–141. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reactive Species | Chemical Formula | Oxidation State(s) of Sulfur |
|---|---|---|
| Thiyl radical | RS• | −1 |
| Disulfide | RSSR | −1 |
| Disulfide-S-monoxide (thiosulfinate) | RS(O)SR’ | +1, −1 |
| Disulfide-S-dioxide (thiosulfonate) | RS(O)2SR’ | +3, −1 |
| Sulfenic acid | RSOH | 0 |
| Sulfinic acid | RSO2H | +2 |
| Interpretation of RSS | Examples |
|---|---|
| Reactive sulfane species | RSxR’, RSxH (R ≠ H, x ≥ 2) |
| Reducing sulfur species | H2S, GSH, protein bound thiols, under specific circumstances possibly Sx2− |
| Radical sulfur species | Primarily RS•, GSSG•−, R’SR•+, HS• |
| Reactive sulfide species | H2Sx and its deprotonated forms |
| Reprocessed sulfur species | Biologically active sulfur metabolites formed by microbiota |
| Reactive sulfur substances | SO2, SO3, S2O32−, H2SO5 (often with limited biological relevance) |
| Reactive sulfur secondary metabolites | Allicin, DADS, DATS, DATTS |
| Recreational sulfur substances | Haarlem oil, 1,2-dithiol-3-thione |
| Regulatory sulfur species | Disulfides, sulfenic and sulfinic acids in proteins and enzymes as part of signalling and control |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giles, G.I.; Nasim, M.J.; Ali, W.; Jacob, C. The Reactive Sulfur Species Concept: 15 Years On. Antioxidants 2017, 6, 38. https://doi.org/10.3390/antiox6020038
Giles GI, Nasim MJ, Ali W, Jacob C. The Reactive Sulfur Species Concept: 15 Years On. Antioxidants. 2017; 6(2):38. https://doi.org/10.3390/antiox6020038
Chicago/Turabian StyleGiles, Gregory I., Muhammad Jawad Nasim, Wesam Ali, and Claus Jacob. 2017. "The Reactive Sulfur Species Concept: 15 Years On" Antioxidants 6, no. 2: 38. https://doi.org/10.3390/antiox6020038
APA StyleGiles, G. I., Nasim, M. J., Ali, W., & Jacob, C. (2017). The Reactive Sulfur Species Concept: 15 Years On. Antioxidants, 6(2), 38. https://doi.org/10.3390/antiox6020038