Selenium Concentrations for Maximisation of Thioredoxin Reductase 2 Activity and Upregulation of Its Gene Transcripts in Senescent Human Fibroblasts

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation of Primary Fibroblast Cultures
2.2. Preparation and Characterization of Senescent Fibroblast Cultures
2.3. Preparation of Se-Supplemented Media
2.4. Experimental Design
2.5. Linearity of TR2 Activity with Respect to Time in Subcultures at Passages 5, 20, and 30 Incubated with MEM2 and MEM8
2.6. Separation of Fibroblast Mitochondria
2.7. TR2 Assay
2.8. H2O2 Generation Rates
2.9. Gene Expression Profiling of hsTR2 Using Real-Time Quantitative PCR (RT-qPCR)
2.10. Other Assays and Statistical Analysis
3. Results
3.1. Se Concentration in Culture Media
3.2. Identification of Pre-Senescent and Senescent Fibroblast Subcultures.
3.2.1. Growth and Replication of Subcultures.
3.2.2. Oxidative Stress Markers are Indicative of Senescence
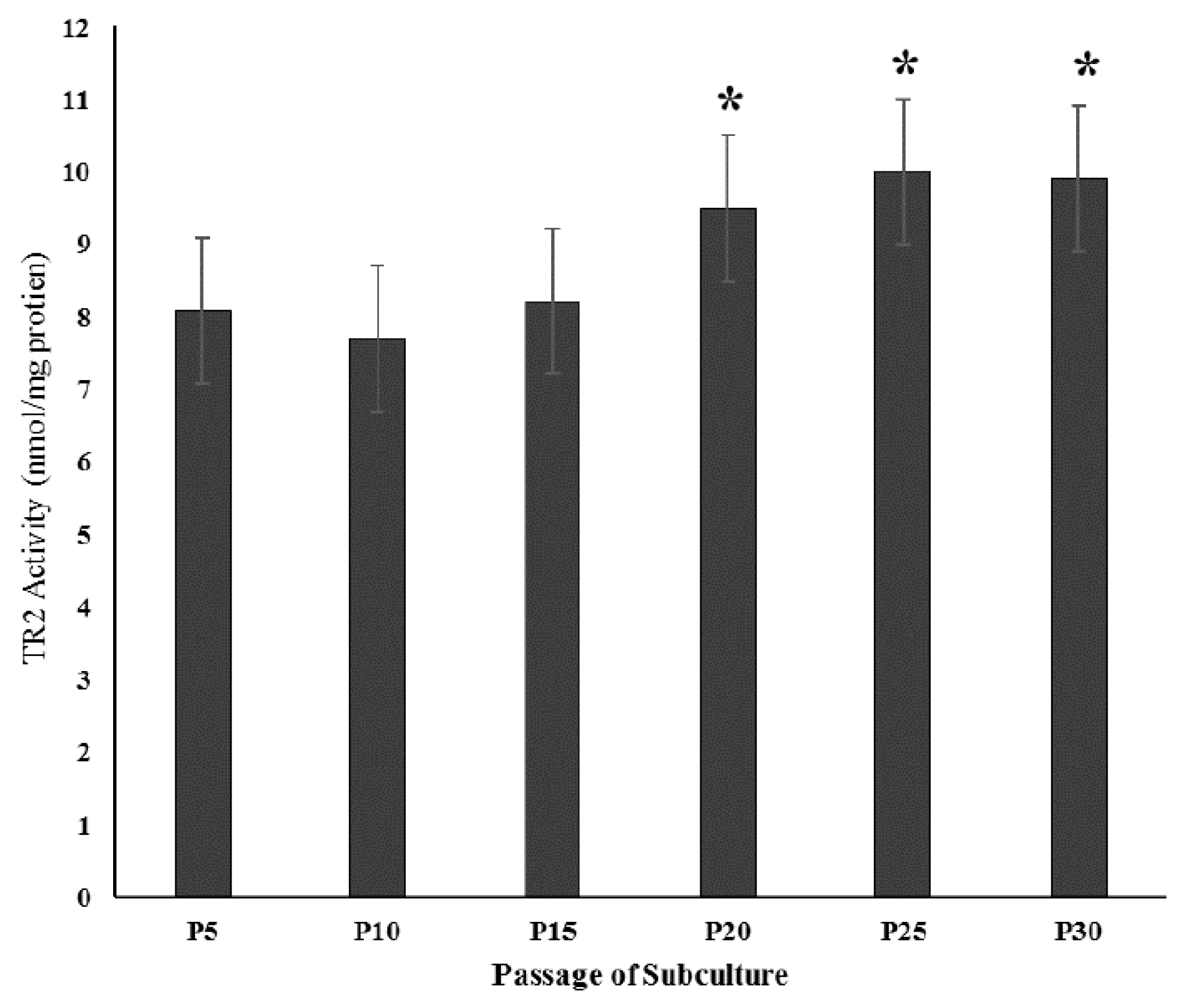
3.2.3. Effect of Senescence on Mitochondrial TR2 Activity in MEM1-Incubated Cultures
3.2.4. Effect of Increased Incubation Time of Pre-Senescent (P5) and Senescent (P20 and 30) Cultures with MEM2 and MEM8 on TR2 Activity
3.3. Effect of Increasing Se Concentrations on TR2 Activity and H2O2 Generation Rates in Pre-Senescent (P5) and Senescent (P20 and 30) Fibroblast Cultures
3.3.1. TR2 Activity
3.3.2. H2O2 Generation
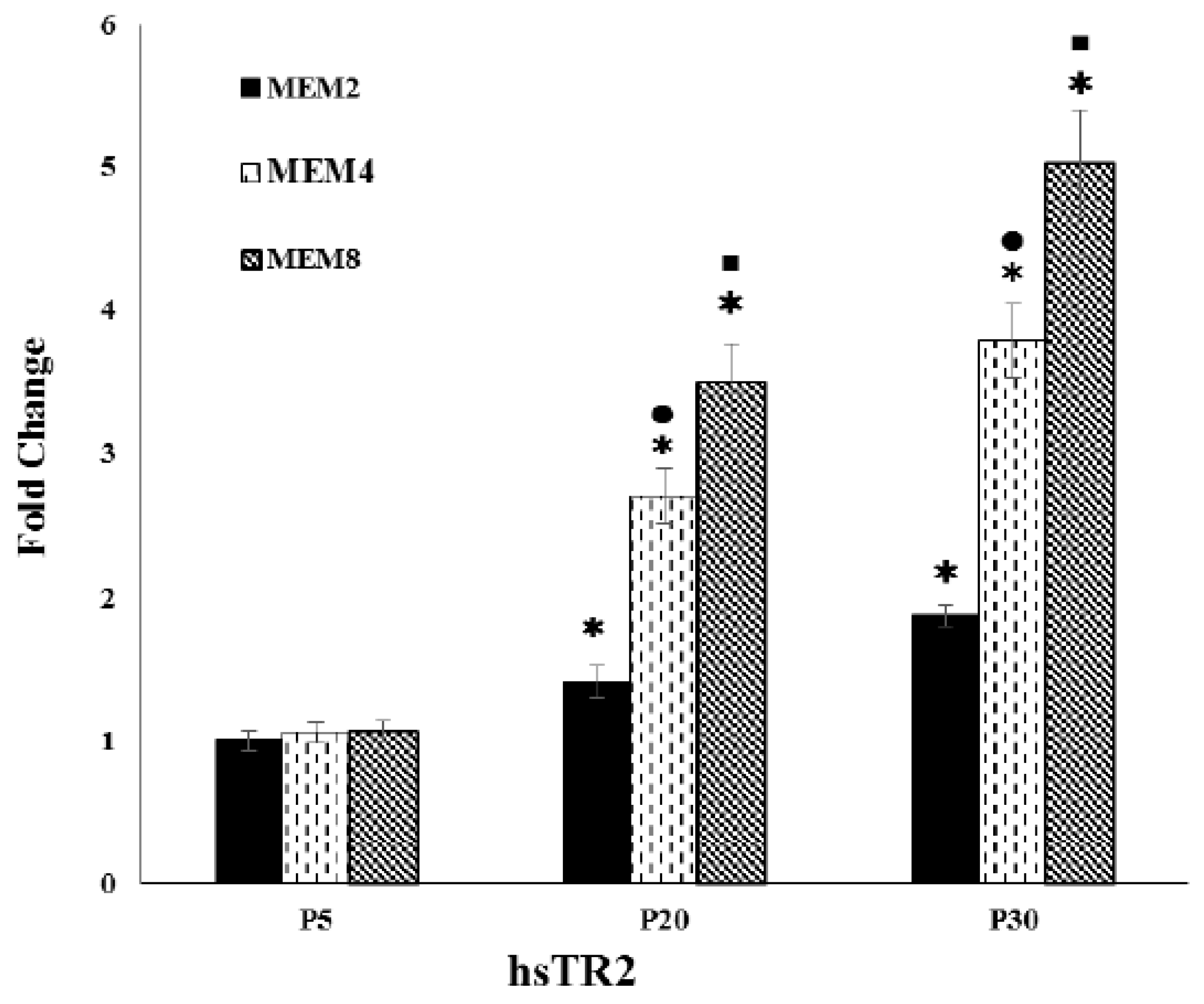
3.4. Effect of Increasing Se Concentrations on TR2 Gene Expression Levels in Pre-Senescent (P5), Early-Senescent (P20) and Senescent (P30) Subcultures
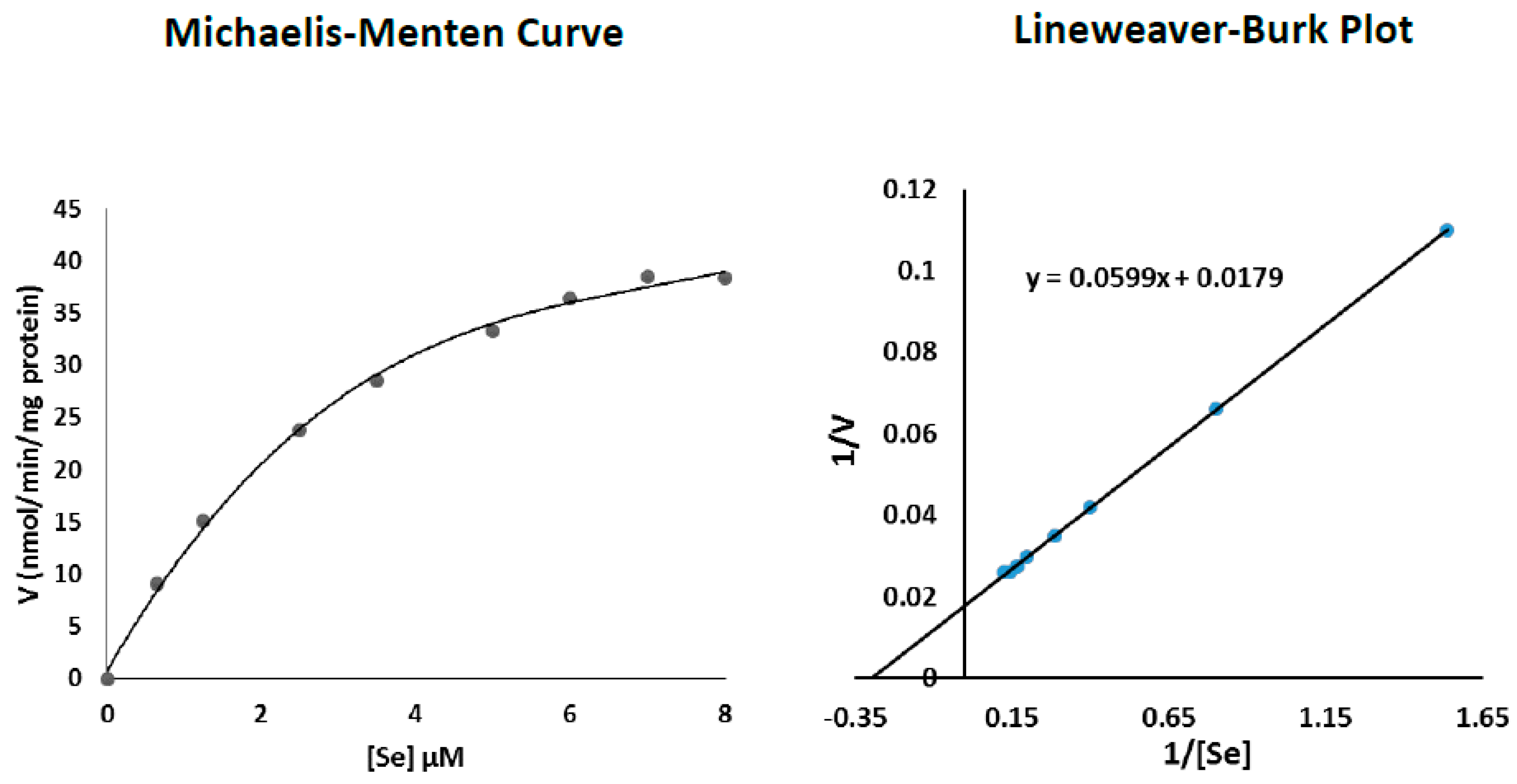
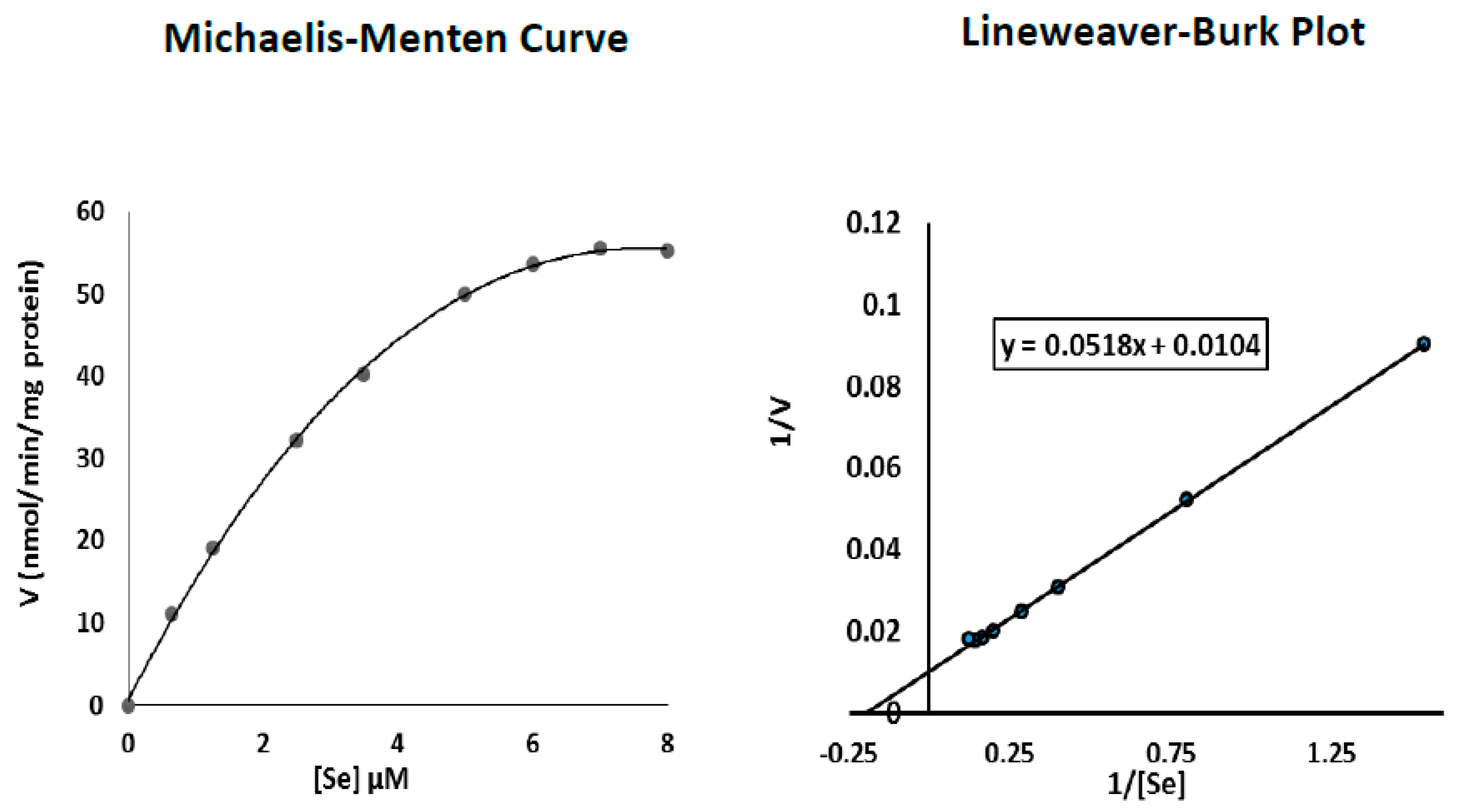
3.5. Determination of Se Km Values with Respect to TR2 Activity in Senescent Fibroblasts
4. Discussion
Acknowledgments
Conflicts of Interest
References
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial ROS and ROS-incubated ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [PubMed]
- Holzereva, E.; Prokisch, H. Mitochondrial: Much ado about nothing? How dangerous is reactive oxygen species production? Int. J. Biochem. Cell. Biol. 2015, 63, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Sena, L.A.; Chandel, N.S. Physiological roles of mitochondrial reactive oxygen species. Mol. Cell 2012, 48, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Holmgren, A. Thioredoxin system in cell death progression. Antioxid. Redox Signal. 2012, 17, 1738–1747. [Google Scholar] [CrossRef] [PubMed]
- Amer, E.S.; Holmgren, A. Physiologic functions of Thioredoxin and Thioredoxin Reductase. Eur. J. Biochem. 2000, 267, 6102–6109. [Google Scholar]
- Amer, E.S.; Holmgren, A. The Thioredoxin system in cancer. Semin. Cancer Biol. 2006, 16, 420–426. [Google Scholar]
- Gladyshev, V.N.; Jeang, K.T.; Stadtman, T.C. Selenocysteine, identified as the penultimate C-terminal residue in human T-cell thioredoxin reductase, corresponds to TGA in the human placental gene. Proc. Natl. Acad. Sci. USA 1996, 93, 6146–6151. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, S.E.; Prast-Nielsen, S.; Flaberg, E.; Szekely, L.; Amer, S.E. High levels of thioredoxin reductase 1 modulate drug-specific cytotoxic efficacy. Free Radic. Biol. Med. 2009, 47, 1661–1667. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.M.; Mayers, C.R. The effects of hexavalent chromium on thioredoxin reductase and peroxiredoxins in human bronchial epithelial cells. Free Radic. Biol. Med. 2009, 47, 1477–1485. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Yang, J.; Deng, S.; Lan, L.; He, F.; Kuang, B.; Zeng, H. A thioredoxin reductase inhibitor induces growth inhibition and apoptosis in cultured human carcinoma cell lines. Cancer Lett. 2006, 236, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Marzano, C.; Gandin, V.; Folda, A.; Scutari, G.; Bindoli, A.; Rigobello, M.P. Inhibition of thioredoxin by auranofin induces apoptosis in cisplatin-resistant human ovarian cancer cells. Free Radic. Biol. Med. 2007, 42, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigo, R.; Gladyshev, V.N. Characterization of mammalian selenoproteomes. Science 2003, 300, 1439–1443. [Google Scholar] [CrossRef] [PubMed]
- Selenius, M.; Randolf, A.K.; Olm, E.; Fernandes, A.P.; Björnstedt, M. Selenium and the selenoprotein thioredoxin reductase in the prevention, treatment and diagnosis of cancer. Antioxid. Redox Signal. 2010, 12, 867–880. [Google Scholar] [CrossRef] [PubMed]
- Lewin, M.H.; Arthur, J.R.; Riemersma, R.A.; Nicol, F.; Walker, S.; Millar, E.M.; Howie, A.F.; Beckett, G.J. Se supplementation acting through induction of Thioredoxin reductase and Glutathione peroxidase protects human endothelial cells line EAhy 929 from damage by lipid hydroperoxidases. Biochim. Biophys. Acta-Mol. Cell. Res. 2002, 1593, 85–92. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhong, L.; Shen, X. Effect of Se supplementation on the calcium signaling in human endothelial cells. J. Cell. Physiol. 2005, 205, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Khera, A.; Vanderlelie, J.J.; Perkins, A.V. Selenium supplementation protects trophoblast cells from mitochondrial OS. Placenta 2013, 34, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Watson, M.; Van Leer, L.; Vanderlelie, J.J.; Perkins, A.V. Selenium supplementation protects trophoblast cells from OS. Placenta 2012, 33, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Ebert, R.; Ulmer, M.; Zeck, S.; Meissner-Weigl, J.; Schneider, D.; Stopper, H.; Schupp, N.; Kassem, M.; Jakob, F. Selenium supplementationrestores the antioxidant capacity and prevents cell damage in bone marrow-stromal cells in vitro. Stem Cells 2006, 24, 1226–1235. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, O.; Stanzial, A.M.; Girelli, D.; Trevisan, M.T.; Guarini, P.; Terz, M.; Caffi, S.; Fontana, F.; Casaril, M.; Ferrari, S. Selenium status, fatty acids, vitamin A and E in aging. The Nove study. Am. J. Clin. Nutr. 1994, 60, 510–517. [Google Scholar] [PubMed]
- Ray, A.L.; Semba, R.D.; Walston, J.; Ferrucci, L.; Cappola, A.R.; Ricks, M.O.; Xue, Q.L.; Fried, L.P. Low serum Selenium and total carotenoids predict mortality among older women living in the community. The women’s health and aging studies. J. Nutr. 2006, 136, 172–176. [Google Scholar] [PubMed]
- Akbaraly, N.J.; Arnaud, J.; Hininger-Favier, I.; Gourlet, V.; Roussel, A.M.; Berr, C. Selenium and mortality in the elderly. Results from the EVA study. Clin. Chem. 2005, 51, 2117–2123. [Google Scholar] [CrossRef] [PubMed]
- Hornsby, P.J.; Harris, S.E. Oxidative damage to DNA and replicative lifespan in cultured adrenocortical cells. Exp. Cell. Res. 1987, 168, 203–217. [Google Scholar] [CrossRef]
- Ghneim, H.K.; Al-Sheikh, Y.A. Effect of selenium supplementation on Glutathione Peroxidase and Catalase Activities in Senescent Cultured Human Fibroblasts. Ann. Nutr. Metab. 2011, 59, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Legarin, Y.; Touat-Hamici, Z.; Chavatte, L. Interplay between Se levels, selenoprotein expression and replicative senescence in WI-38 human fibroblasts. J. Biol. Chem. 2014, 289, 6299–6310. [Google Scholar] [CrossRef] [PubMed]
- Ghneim, H.K.; Al-Sheikh, Y.A. The effect of micronutrients on superoxide dismutase in senescent fibroblasts. Cell. Biochem. Funct. 2011, 29, 384–394. [Google Scholar]
- Ghneim, H.K.; Al-Sheikh, Y.A. The effect of aging and increasing ascorbate concentrations on respiratory chain activity in cultured human fibroblast. Cell. Biochem. Funct. 2010, 28, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Ghneim, H.K. The effect of Echis coloratus venom on biochemical and molecular markers of the antioxidant capacity in human fibroblasts. Libyan J. Med. 2017, 12, 1304515. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Diwu, Z.; Panchuk-Voloshina, N.; Haugland, R.P. A stable nonfluorescent derivative of resorufin for the fluorometric determination of trace hydrogen peroxide: Applications in detecting the activity of phagocyte NADH oxidase and other oxidases. Anal. Biochem. 1997, 253, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Ghneim, H.K.; Alshebly, M.M. Biochemical markers of oxidative stress in Saudi women with recurrent miscarriage. J. Korean Med. Sci. 2016, 31, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Ghneim, H.K.; Al-Sheikh, Y.A.; Alshebly, M.M.; Aboul-Soud, M.A.M. Superoxide dismutase activity and Gene expression levels in Saudi women with recurrent miscarriage. Mol. Med. Rep. 2016, 13, 2606–2612. [Google Scholar] [CrossRef] [PubMed]
- Saquib, Q.; Attia, S.M.; Siddiqui, M.A.; Aboul-Soud, M.A.M.; Al-Khedairy, A.A.; Giesy, J.P.; Musarrat, J. Phorate-induced oxidative stress, DNA damage and transcriptional activation of P53 and caspase genes in male Wistar rats. Toxicol. Appl. Pharmacol. 2012, 259, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, B.E.; Lockitch, G. Direct determination of Se in serum by graphite-furnace atomiv absorption spectrophotometry with deuterium background correction and reduced palladium modifier: Age-specific reference ranges. Clin. Chem. 1988, 34, 709–714. [Google Scholar] [PubMed]
- Bradford, M.M. A rapid sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ghneim, H.K. Characterization of the aging of serially subcultured human skin fibroblasts. Med. Sci. Res. 1993, 21, 249–250. [Google Scholar]
- Campisi, J.; D’Adda di Fagagna, F. Cellular senescence: When bad things happen to good cells. Nat. Rev. Mol. Cell. Biol. 2007, 8, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Rodier, F.; Campisi, J. Four faces of cellular senescence. J. Cell. Biol. 2011, 192, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Ghneim, H.K. The kinetics of the effect of manganese supplementation on SOD2 activity in senescent human fibroblasts. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 1866–1880. [Google Scholar] [PubMed]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Haratake, M.; Hongoh, M.; Ono, M.; Nakayama, M. Thiol-dependent membrane transport of selenium through an integral protein of the red blood cell membrane. Inorg. Chem. 2009, 48, 7805–7811. [Google Scholar] [CrossRef] [PubMed]
- McDermott, J.R.; Rosen, B.P.; Liu, Z.J. Jen1P: A high affinity selenite transporter in yeast. Mol. Biol. Cell 2010, 21, 3934–3941. [Google Scholar] [CrossRef] [PubMed]
- Tarze, A.; Dauplais, M.; Grigoras, I.; Lazard, M.; Ha-Duong, N.T.; Barbier, F.; Blanquet, S.; Plateau, P. Extracellular production of hydrogen selenide accounts for thiol-assisted toxicity of selenite against Saccharomyces cerevisiae. J. Biol. Chem. 2007, 282, 8759–8767. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fibroblast Cultures (n = 6)/Passage of Subcultures | Oxidative Stress Markers | ||||
|---|---|---|---|---|---|
| H2O2 | SOA | LPO | PCC | GSH/GSSG | |
| pmol/min/mg protein | µmol/min/mg protein | pmol/min/mg protein | nmol/mg | ||
| P5 | 3.41 ± 0.29 | 1.03 ± 0.06 | 71.6 ± 5.63 | 4.88 ± 0.40 | 61.3 ± 3.98 |
| P10 | 3.28 ± 0.26 | 0.97 ± 0.06 | 73.5 ± 5.68 | 4.91 ± 0.41 | 59.6 ± 3.75 |
| P15 | 3.85 ± 0.33 * | 1.12 ± 0.07 * | 81.7 ± 6.29 * | 5.56 ± 0.46 * | 55.1 ± 3.65 * |
| P20 | 7.80 ± 0.63 ** | 1.27 ± 0.09 ** | 87.4 ± 6.69 ** | 6.01 ± 0.47 ** | 51.2 ± 3.24 ** |
| P25 | 12.6 ± 1.10 *** | 3.95 ± 0.24 *** | 272.3 ± 20.1 *** | 10.8 ± 0.85 *** | 28.3 ± 1.70 *** |
| P30 | 13.8 ± 1.14 *** | 4.14 ± 0.25 *** | 281.7 ± 21.4 *** | 11.3 ± 0.86 *** | 29.5 ± 1.71 *** |
| Incubation Time (Hours) | Subculture Passages TR2 Activity (nmol/min/mg protein) | |||||
|---|---|---|---|---|---|---|
| P5 | P20 | P30 | ||||
| MEM2 | MEM8 | MEM2 | MEM8 | MEM2 | MEM8 | |
| 1 | 6.01 ± 0.39 | 6.10 ± 0.41 | 9.22 ± 0.55 | 22.4 ± 1.30 | 9.60 ± 0.57 | 23.3 ± 1.37 |
| 2 | 6.94 ± 0.428 * | 6.91 ± 0.43 * | 10.6 ± 0.67 * | 28.2 ± 1.72 ** | 12.1 ± 0.74 ** | 27.1 ± 1.66 * |
| 4 | 8.09 ± 0.48 ** | 8.12 ± 0.53 ** | 12.4 ± 0.75 ** | 32.1 ± 1.93 ** | 14.3 ± 0.87 ** | 36.8 ± 2.28 ** |
| 6 | 8.03 ± 0.49 | 8.10 ± 0.49 | 15.6 ± 0.98 ** | 37.9 ± 2.35 **,■ | 16.6 ± 0.98 ** | 45.6 ± 2.87 ** |
| 8 | 7.98 ± 0.49 | 7.94 ± 0.48 | 15.4 ± 0.93 | 38.1 ± 2.29 | 19.4 ± 1.18 ** | 54.7 ± 3.34 **,■ |
| 10 | 8.12 ± 0.52 | 7.99 ± 0.49 | 15.1 ± 0.91 | 38.4 ± 2.36 | 19.2 ± 1.13 | 55.3 ± 3.31 |
| 12 | 8.02 ± 0.49 | 8.08 ± 0.52 | 15.0 ± 0.92 | 38.3 ± 2.31 | 19.5 ± 1.19 | 56.0 ± 3.34 |
| Passage of Subculture (n = 6) | TR2 Activity (nmol/min/mg protein) | |||||||
|---|---|---|---|---|---|---|---|---|
| MEM1 | MEM2 | MEM3 | MEM4 | MEM5 | MEM6 | MEM7 | MEM8 | |
| P5 | 8.08 ± 0.78 | 8.31 ± 0.80 | 8.19 ± 0.79 | 7.92 ± 0.75 | 7.81 ± 0.74 | 8.11 ± 0.79 | 8.36 ± 0.81 | 8.22 ± 0.80 |
| P20 | 9.50 ± 0.90 | 15.1 ± 1.38 * | 23.8 ± 2.25 *,■ | 28.6 ± 2.74 *,■■ | 33.3 ± 3.11 *,■■■ | 36.4 ± 3.20 * | 38.5 ± 3.71 * | 38.4 ± 3.68 * |
| P30 | 9.90 ± 1.21 | 19.2 ± 1.73 * | 32.3 ± 2.90 *,● | 40.2 ± 3.10 *,●● | 50.0 ± 4.69 *,●●● | 53.7 ± 4.78 *,▲ | 55.6 ± 5.22 *,▲ | 55.3 ± 5.09 *,▲ |
| Passage of Subculture (n = 6) | H2O2 Generation (pmol/min/mg Protein) | |||||||
|---|---|---|---|---|---|---|---|---|
| MEM1 | MEM2 | MEM3 | MEM4 | MEM5 | MEM6 | MEM7 | MEM8 | |
| P5 | 3.41 ± 0.29 | 3.20 ± 0.28 | 3.52 ± 0.30 | 3.58 ± 0.31 | 3.35 ± 0.30 | 3.40 ± 0.28 | 3.61 ± 0.32 | 3.33 ± 0.28 |
| P20 | 7.80 ± 0.63 | 7.12 ± 0.58 | 6.54 ± 0.53 * | 6.03 ± 0.50 ** | 5.62 ± 0.47 ** | 5.15 ± 0.42 ** | 4.64 ± 0.38 ** | 4.28 ± 0.34 ** |
| P30 | 13.8 ± 1.14 | 10.3 ± 0.83 ** | 9.10 ± 0.74 **,■ | 8.10 ± 0.65 **,■ | 6.99 ± 0.58 **,■ | 5.69 ± 0.49 **,■■ | 4.78 ± 0.40 **,■■ | 4.22 ± 0.35 **,■ |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghneim, H.K. Selenium Concentrations for Maximisation of Thioredoxin Reductase 2 Activity and Upregulation of Its Gene Transcripts in Senescent Human Fibroblasts. Antioxidants 2017, 6, 83. https://doi.org/10.3390/antiox6040083
Ghneim HK. Selenium Concentrations for Maximisation of Thioredoxin Reductase 2 Activity and Upregulation of Its Gene Transcripts in Senescent Human Fibroblasts. Antioxidants. 2017; 6(4):83. https://doi.org/10.3390/antiox6040083
Chicago/Turabian StyleGhneim, Hazem K. 2017. "Selenium Concentrations for Maximisation of Thioredoxin Reductase 2 Activity and Upregulation of Its Gene Transcripts in Senescent Human Fibroblasts" Antioxidants 6, no. 4: 83. https://doi.org/10.3390/antiox6040083
APA StyleGhneim, H. K. (2017). Selenium Concentrations for Maximisation of Thioredoxin Reductase 2 Activity and Upregulation of Its Gene Transcripts in Senescent Human Fibroblasts. Antioxidants, 6(4), 83. https://doi.org/10.3390/antiox6040083