Superoxide Dismutase Mimetic GC4419 Enhances the Oxidation of Pharmacological Ascorbate and Its Anticancer Effects in an H2O2-Dependent Manner

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Media
2.2. Drug Treatment
2.3. Electron Paramagnetic Resonance (EPR) Spectroscopy
2.4. Oxygen Consumption by Clark Electrode
2.5. H2O2 Quantification
2.6. SOD Activity Assay
2.7. Clonogenic Assay
2.8. Lentivirus Production and Transduction
2.9. Statistical Analysis
3. Results
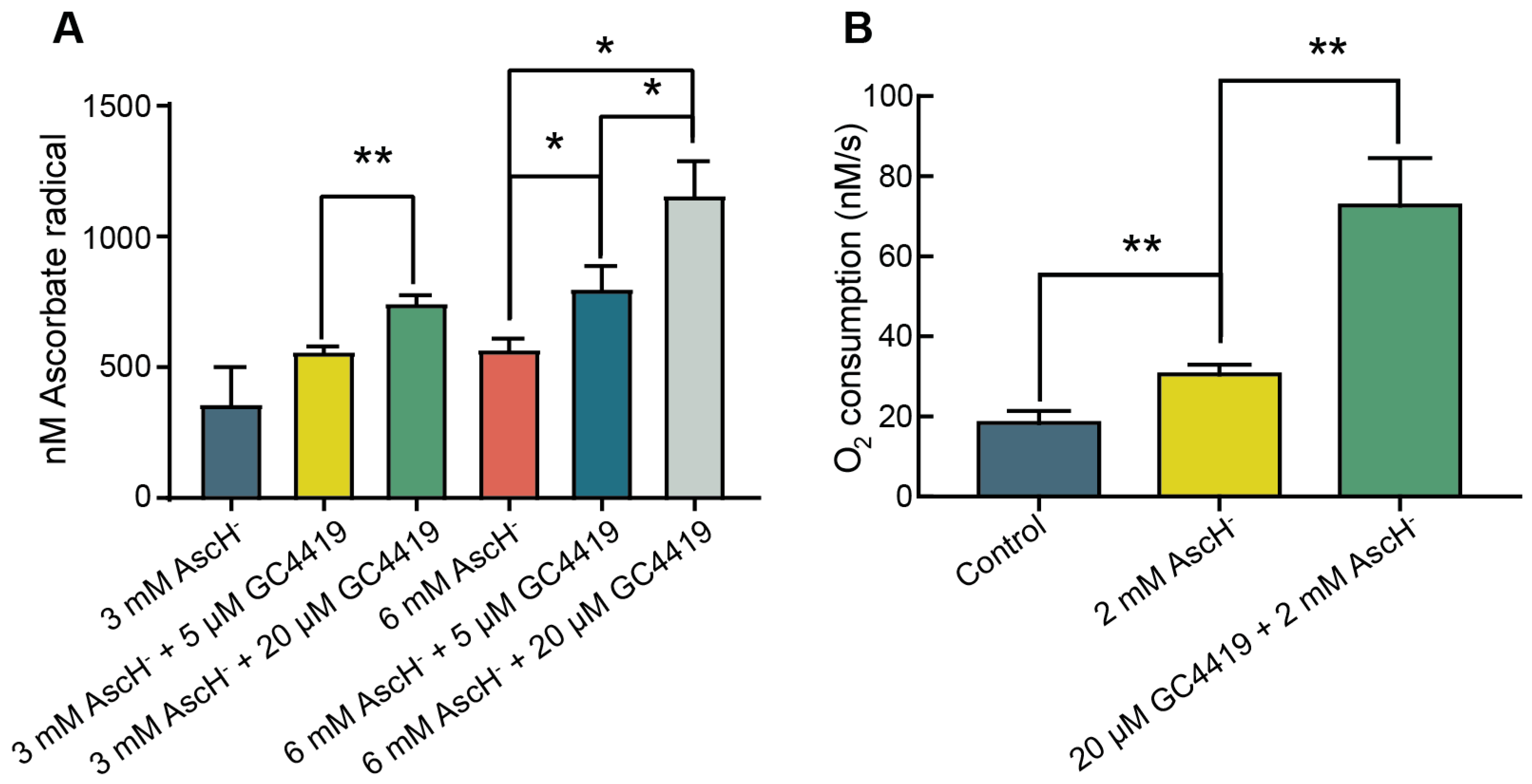
3.1. GC4419 Increases the Rate of Oxygen Consumption and Ascorbate Radical Steady-State Concentration in Systems Containing AscH−
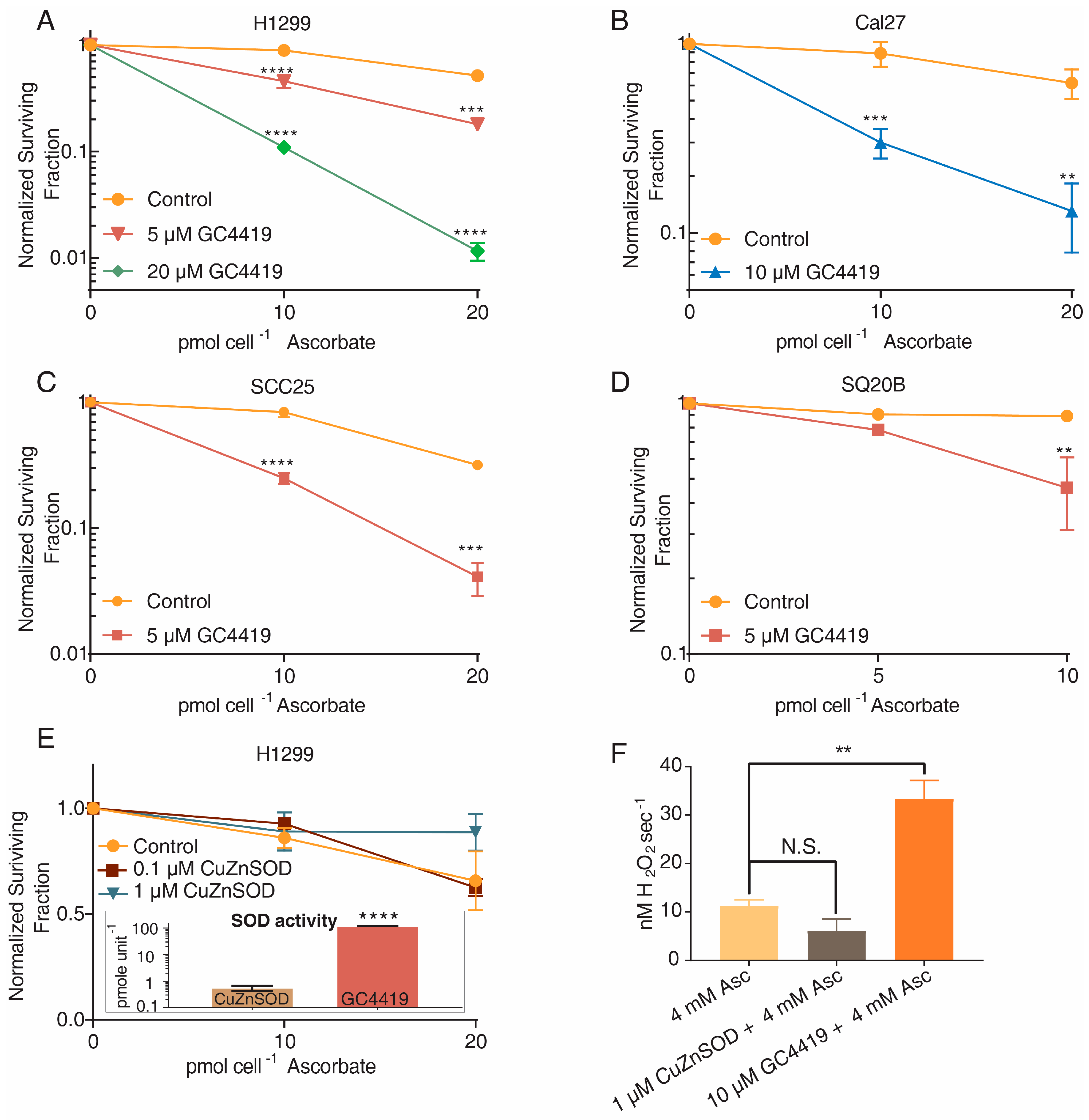
3.2. GC4419, but Not CuZnSOD, Increases H2O2 Production and Cancer Cell Toxicity when Combined with AscH−
3.3. GC/AscH− Toxicity is Dependent on H2O2
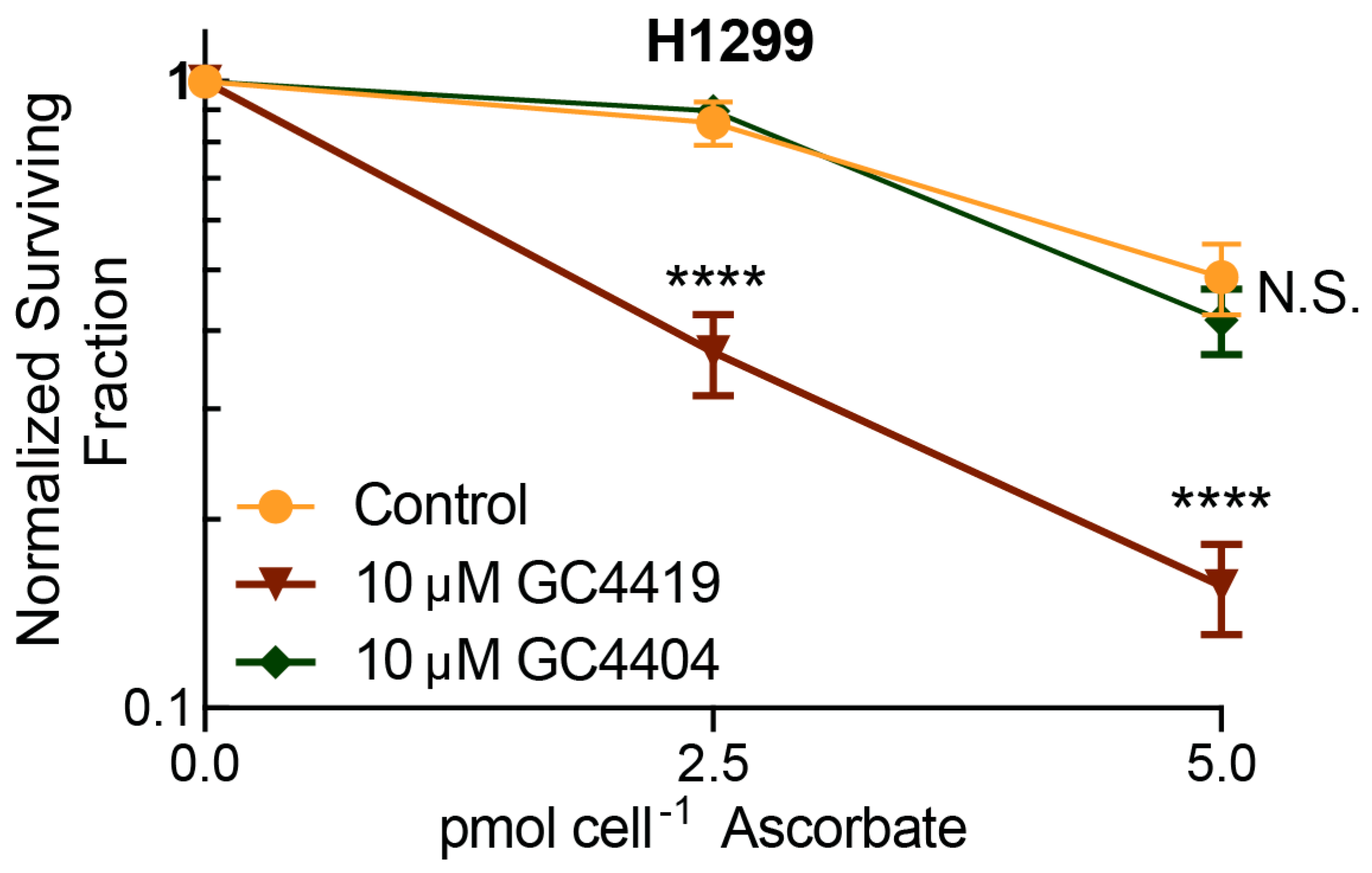
3.4. Catalytic Activity of Mn(II)-Pentaazamacrocylces is Required for Enhancement of AscH− Toxicity
3.5. GC/AscH− Treatment Increases Cancer Cell Response to Ionizing Radiation
4. Discussion
- (1)
- (2)
- Native CuZnSOD does not enhance cancer cell toxicity or H2O2 production when combined with AscH−, supporting the hypothesis that the SOD activity of GC4419 is not responsible for increased AscH− toxicity;
- (3)
- Addition of GC4419 to solutions containing AscH− results in a significant increase in [Asc•−]ss, supporting the hypothesis that addition of GC4419 increases the one-electron oxidation of AscH−;
- (4)
- The structurally-similar Mn(II)-pentaazamacrocycle complex, GC4404, which does not form the Mn(III)-intermediate which has been suggested to be reduced by AscH− [35], does not enhance AscH− cancer cell toxicity, strongly supporting the hypothesis that catalytic cycling of the pentaazamacrocycle is required for the enhancement of AscH− toxicity; and
- (5)
- GC4419 enhances H2O2 production when combined with AscH−, and the cancer cell toxicity of GC/AscH− is completely inhibited by catalase overexpression, demonstrating that H2O2 is responsible for the toxicity of GC/AscH−.
5. Conclusions
6. Patents
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.L.; Wagner, B.A.; van’t Erve, T.J.; Zehr, P.S.; Berg, D.J.; Halfdanarson, T.R.; Yee, N.S.; Bodeker, K.L.; Du, J.; Roberts, L.J.; et al. Pharmacological ascorbate with gemcitabine for the control of metastatic and node-positive pancreatic cancer (PACMAN): Results from a phase I clinical trial. Cancer Chemother. Pharmacol. 2013, 71, 765–775. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, C.M.; Levin, R.D.; Spector, T.; Lis, C.G. Phase I clinical trial to evaluate the safety, tolerability, and pharmacokinetics of high-dose intravenous ascorbic acid in patients with advanced cancer. Cancer Chemother. Pharmacol. 2013, 72, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Chapman, J.; Levine, M.; Polireddy, K.; Drisko, J.; Chen, Q. High-dose parenteral ascorbate enhanced chemosensitivity of ovarian cancer and reduced toxicity of chemotherapy. Sci. Transl. Med. 2014, 6, 222ra18. [Google Scholar] [CrossRef] [PubMed]
- Schoenfeld, J.D.; Sibenaller, Z.A.; Mapuskar, K.A.; Wagner, B.A.; Cramer-Morales, K.L.; Furqan, M.; Sandhu, S.; Carlisle, T.L.; Smith, M.C.; Abu Hejleh, T.; et al. O2•− and H2O2-Mediated Disruption of Fe Metabolism Causes the Differential Susceptibility of NSCLC and GBM Cancer Cells to Pharmacological Ascorbate. Cancer Cell 2017, 31, 487–500.e8. [Google Scholar] [CrossRef] [PubMed]
- Buettner, G.R. The pecking order of free radicals and antioxidants: Lipid peroxidation, alpha-tocopherol, and ascorbate. Arch. Biochem. Biophys. 1993, 300, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Martin, S.M.; Levine, M.; Wagner, B.A.; Buettner, G.R.; Wang, S.; Taghiyev, A.F.; Du, C.; Knudson, C.M.; Cullen, J.J. Mechanisms of ascorbate-induced cytotoxicity in pancreatic cancer. Clin. Cancer Res. 2010, 16, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Buettner, G.R. In the absence of catalytic metals ascorbate does not autoxidize at pH 7: Ascorbate as a test for catalytic metals. J. Biochem. Biophys. Methods 1988, 16, 27–40. [Google Scholar] [CrossRef]
- Du, J.; Wagner, B.A.; Buettner, G.R.; Cullen, J.J. Role of labile iron in the toxicity of pharmacological ascorbate. Free Radic. Biol. Med. 2015, 84, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Rawal, M.; Schroeder, S.R.; Wagner, B.A.; Cushing, C.M.; Welsh, J.L.; Button, A.M.; Du, J.; Sibenaller, Z.A.; Buettner, G.R.; Cullen, J.J. Manganoporphyrins increase ascorbate-induced cytotoxicity by enhancing H2O2 generation. Cancer Res. 2013, 73, 5232–5241. [Google Scholar] [CrossRef] [PubMed]
- Tovmasyan, A.; Sampaio, R.S.; Boss, M.-K.; Bueno-Janice, J.C.; Bader, B.H.; Thomas, M.; Reboucas, J.S.; Orr, M.; Chandler, J.D.; Go, Y.-M.; et al. Anticancer therapeutic potential of Mn porphyrin/ascorbate system. Free Radic. Biol. Med. 2015, 89, 1231–1247. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.K.; Tovmasyan, A.; Batinic-Haberle, I.; Devi, G.R. Mn porphyrin in combination with ascorbate acts as a pro-oxidant and mediates caspase-independent cancer cell death. Free Radic. Biol. Med. 2014, 68, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Cieslak, J.A.; Strother, R.K.; Rawal, M.; Du, J.; Doskey, C.M.; Schroeder, S.R.; Button, A.; Wagner, B.A.; Buettner, G.R.; Cullen, J.J. Manganoporphyrins and ascorbate enhance gemcitabine cytotoxicity in pancreatic cancer. Free Radic. Biol. Med. 2015, 83, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Batinić-Haberle, I.; Rajić, Z.; Benov, L. A Combination of Two Antioxidants (An SOD Mimic and Ascorbate) Produces a Pro-Oxidative Effect Forcing Escherichia coli to Adapt via Induction of oxyR Regulon. Anticancer Agents Med. Chem. 2011, 11, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Salvemini, D.; Mazzon, E.; Dugo, L.; Riley, D.P.; Serraino, I.; Caputi, A.P.; Cuzzocrea, S. Pharmacological manipulation of the inflammatory cascade by the superoxide dismutase mimetic, M40403. Br. J. Pharmacol. 2001, 132, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Salvemini, D.; Mazzon, E.; Dugo, L.; Serraino, I.; De Sarro, A.; Caputi, A.P.; Cuzzocrea, S. Amelioration of joint disease in a rat model of collagen-induced arthritis by M40403, a superoxide dismutase mimetic. Arthritis Rheum. 2001, 44, 2909–2921. [Google Scholar] [CrossRef]
- Masini, E.; Cuzzocrea, S.; Mazzon, E.; Marzocca, C.; Mannaioni, P.F.; Salvemini, D. Protective effects of M40403, a selective superoxide dismutase mimetic, in myocardial ischaemia and reperfusion injury in vivo. Br. J. Pharmacol. 2002, 136, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Aston, K.; Rath, N.; Naik, A.; Slomczynska, U.; Schall, O.F.; Riley, D.P. Computer-Aided Design (CAD) of Mn(II) Complexes: Superoxide Dismutase Mimetics with Catalytic Activity Exceeding the Native Enzyme. Inorg. Chem. 2001, 40, 1779–1789. [Google Scholar] [CrossRef] [PubMed]
- Mapuskar, K.A.; Flippo, K.H.; Schoenfeld, J.D.; Riley, D.P.; Strack, S.; Abu-Hejleh, T.; Furqan, M.; Monga, V.; Domann, F.E.; Buatti, J.M.; et al. Mitochondrial superoxide increases age-associated susceptibility of human dermal fibroblasts to radiation and chemotherapy. Cancer Res. 2017. [CrossRef] [PubMed]
- Anderson, C.M.; Sonis, S.T.; Lee, C.M.; Adkins, D.; Allen, B.G.; Sun, W.; Agarwala, S.S.; Venigalla, M.L.; Chen, Y.; Zhen, W.; et al. Phase 1b/2a Trial of The Superoxide Dismutase Mimetic GC4419 to Reduce Chemoradiotherapy-induced Oral Mucositis in Patients with Oral Cavity or Oropharyngeal Carcinoma. Int. J. Radiat. Oncol. Biol. Phys. 2017. [CrossRef] [PubMed]
- Kelso, G.F.; Maroz, A.; Cochemé, H.M.; Logan, A.; Prime, T.A.; Peskin, A.V.; Winterbourn, C.C.; James, A.M.; Ross, M.F.; Brooker, S.; et al. A Mitochondria-Targeted Macrocyclic Mn(II) Superoxide Dismutase Mimetic. Chem. Biol. 2012, 19, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Doskey, C.M.; van’t Erve, T.J.; Wagner, B.A.; Buettner, G.R. Moles of a Substance per Cell Is a Highly Informative Dosing Metric in Cell Culture. PLoS ONE 2015, 10, e0132572. [Google Scholar] [CrossRef] [PubMed]
- Spitz, D.R.; Dewey, W.C.; Li, G.C. Hydrogen peroxide or heat shock induces resistance to hydrogen peroxide in Chinese hamster fibroblasts. J. Cell. Physiol. 1987, 131, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Doskey, C.M.; Buranasudja, V.; Wagner, B.A.; Wilkes, J.G.; Du, J.; Cullen, J.J.; Buettner, G.R. Tumor cells have decreased ability to metabolize H2O2: Implications for pharmacological ascorbate in cancer therapy. Redox Biol. 2016, 10, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Buettner, G.R.; Kiminyo, K.P. Optimal EPR detection of weak nitroxide spin adduct and ascorbyl free radical signals. J. Biochem. Biophys. Methods 1992, 24, 147–151. [Google Scholar] [CrossRef]
- Spitz, D.R.; Oberley, L.W. An assay for superoxide dismutase activity in mammalian tissue homogenates. Anal. Biochem. 1989, 179, 8–18. [Google Scholar] [CrossRef]
- Brandt, K.E.; Falls, K.C.; Schoenfeld, J.D.; Rodman, S.N.; Gu, Z.; Zhan, F.; Cullen, J.J.; Wagner, B.A.; Buettner, G.R.; Allen, B.G.; et al. Augmentation of intracellular iron using iron sucrose enhances the toxicity of pharmacological ascorbate in colon cancer cells. Redox Biol. 2018, 14, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Spitz, D.R.; Elwell, J.H.; Sun, Y.; Oberley, L.W.; Oberley, T.D.; Sullivan, S.J.; Roberts, R.J. Oxygen toxicity in control and H2O2-resistant Chinese hamster fibroblast cell lines. Arch. Biochem. Biophys. 1990, 279, 249–260. [Google Scholar] [CrossRef]
- Buettner, G.R.; Jurkiewicz, B.A. Ascorbate free radical as a marker of oxidative stress: An EPR study. Free Radic. Biol. Med. 1993, 14, 49–55. [Google Scholar] [CrossRef]
- Goldstein, S.; Fridovich, I.; Czapski, G. Kinetic properties of Cu,Zn-superoxide dismutase as a function of metal content—order restored. Free Radic. Biol. Med. 2006, 41, 937–941. [Google Scholar] [CrossRef] [PubMed]
- Spitz, D.R.; Azzam, E.I.; Li, J.J.; Gius, D. Metabolic oxidation/reduction reactions and cellular responses to ionizing radiation: A unifying concept in stress response biology. Cancer Metastasis Rev. 2004, 23, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Cullen, J.J.; Buettner, G.R. Ascorbic acid: Chemistry, biology and the treatment of cancer. Biochim. Biophys. Acta 2012, 1826, 443–457. [Google Scholar] [CrossRef] [PubMed]
- Buettner, G.R.; Jurkiewicz, B.A. Catalytic metals, ascorbate and free radicals: Combinations to avoid. Radiat. Res. 1996, 145, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Riley, D.P.; Lennon, P.J.; Neumann, W.L.; Weiss, R.H. Toward the rational design of superoxide dismutase mimics: Mechanistic studies for the elucidation of substituent effects on the catalytic activity of macrocyclic manganese(ii) complexes. J. Am. Chem. Soc. 1997, 119, 6522–6528. [Google Scholar] [CrossRef]
- Riley, D.P.; Schall, O.F. Structure—Activity Studies and the Design of Synthetic Superoxide Dismutase (SOD) Mimetics as Therapeutics. In Advances in Inorganic Chemistry: Template Effects and Molecular Organization; van Eldik, R., Bowman-James, K., Eds.; Academic Press: Cambridge, MA, USA, 2006; Volume 59, pp. 233–263. [Google Scholar]
- Maroz, A.; Kelso, G.F.; Smith, R.A.J.; Ware, D.C.; Anderson, R.F. Pulse radiolysis investigation on the mechanism of the catalytic action of Mn(II)−pentaazamacrocycle compounds as superoxide dismutase mimetics. J. Phys. Chem. A 2008, 112, 4929–4935. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heer, C.D.; Davis, A.B.; Riffe, D.B.; Wagner, B.A.; Falls, K.C.; Allen, B.G.; Buettner, G.R.; Beardsley, R.A.; Riley, D.P.; Spitz, D.R. Superoxide Dismutase Mimetic GC4419 Enhances the Oxidation of Pharmacological Ascorbate and Its Anticancer Effects in an H2O2-Dependent Manner. Antioxidants 2018, 7, 18. https://doi.org/10.3390/antiox7010018
Heer CD, Davis AB, Riffe DB, Wagner BA, Falls KC, Allen BG, Buettner GR, Beardsley RA, Riley DP, Spitz DR. Superoxide Dismutase Mimetic GC4419 Enhances the Oxidation of Pharmacological Ascorbate and Its Anticancer Effects in an H2O2-Dependent Manner. Antioxidants. 2018; 7(1):18. https://doi.org/10.3390/antiox7010018
Chicago/Turabian StyleHeer, Collin D., Andrew B. Davis, David B. Riffe, Brett A. Wagner, Kelly C. Falls, Bryan G. Allen, Garry R. Buettner, Robert A. Beardsley, Dennis P. Riley, and Douglas R. Spitz. 2018. "Superoxide Dismutase Mimetic GC4419 Enhances the Oxidation of Pharmacological Ascorbate and Its Anticancer Effects in an H2O2-Dependent Manner" Antioxidants 7, no. 1: 18. https://doi.org/10.3390/antiox7010018
APA StyleHeer, C. D., Davis, A. B., Riffe, D. B., Wagner, B. A., Falls, K. C., Allen, B. G., Buettner, G. R., Beardsley, R. A., Riley, D. P., & Spitz, D. R. (2018). Superoxide Dismutase Mimetic GC4419 Enhances the Oxidation of Pharmacological Ascorbate and Its Anticancer Effects in an H2O2-Dependent Manner. Antioxidants, 7(1), 18. https://doi.org/10.3390/antiox7010018