Singlet Oxygen and Free Radical Reactions of Retinoids and Carotenoids—A Review



Abstract
:1. Introduction
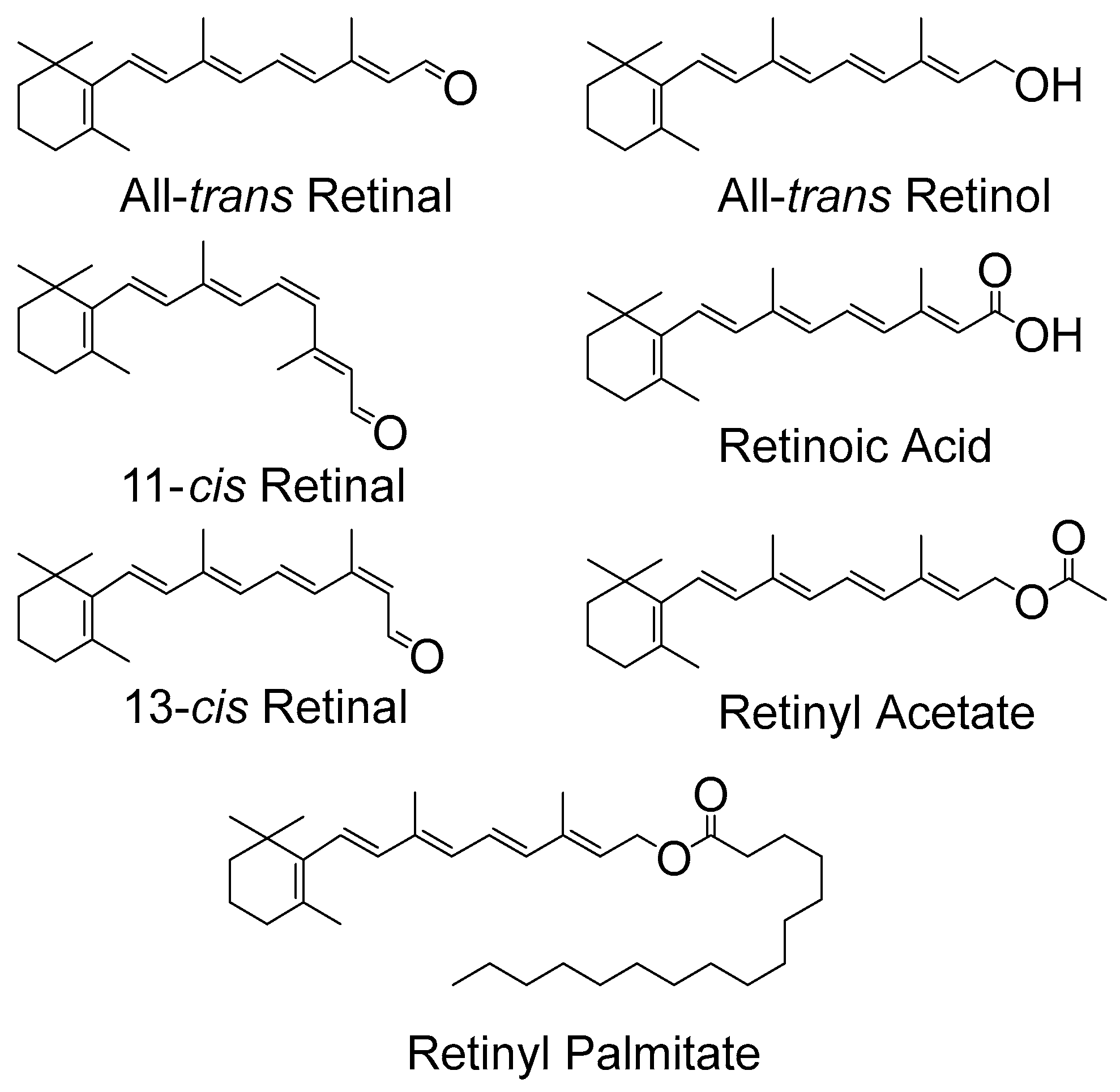
2. The Retinoids
2.1. Triplet States and Singlet Oxygen (1O2)
2.2. Retinoid Radicals
2.3. Radical Reactions
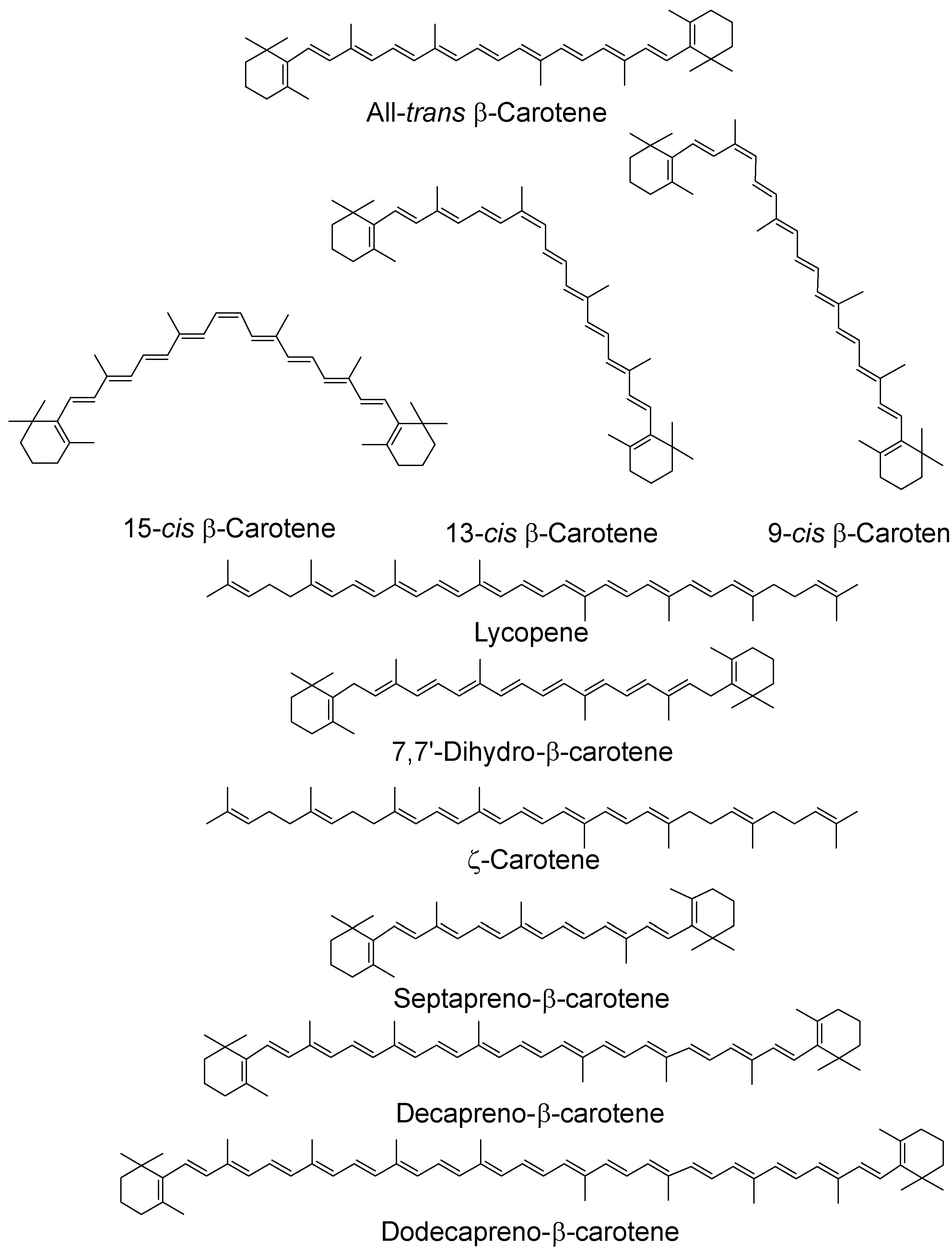
3. Carotenoids
3.1. Carotenoids and Singlet Oxygen
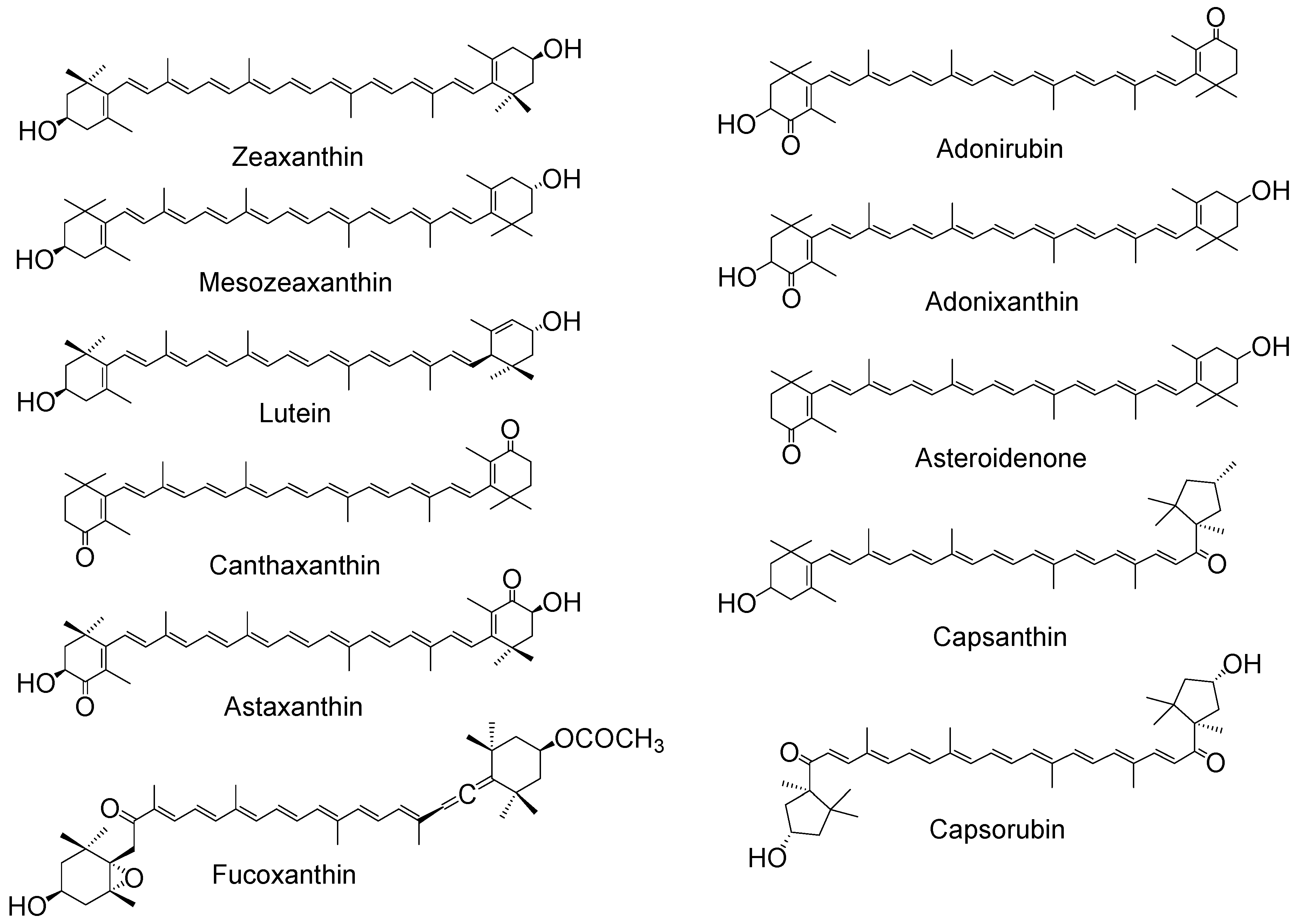
3.2. Carotenoid and Xanthophyll Radicals
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Bensasson, R.V.; Land, E.J.; Truscott, T.G. Excited States and Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 1993; pp. 201–227. [Google Scholar]
- Zhao, D.-Y.; Wintch, S.W.; Ermakov, I.V.; Gellermann, W.; Bernstein, P.S. Resonance Raman measurement of macular carotenoids in retinal, choroidal, and macular dystrophies. Arch. Opthalmol. 2003, 121, 967–972. [Google Scholar] [CrossRef] [PubMed]
- Frank, H.A.; Cogdell, R.J. Carotenoids in photosynthesis. Photochem. Photobiol. 1996, 63, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Mathews-Roth, M.M. Treatment of erythropoietic protoporphyria with beta-carotene. Photo-dermatology 1984, 1, 318–321. [Google Scholar] [CrossRef] [PubMed]
- Garone, M., Jr.; Howard, J.; Fabrikant, J. A review of common tanning methods. J. Clin. Aesthet. Dermatol. 2015, 8, 43–47. [Google Scholar] [PubMed]
- Mukai, K.; Ouchi, A.; Azuma, N.; Takahashi, S.; Aizawa, K.; Nagaoka, S. Development of a singlet oxygen absorption capacity (SOAC) assay method. Measurements of the SOAC values for carotenoids and α-tocopherol in an aqueous Triton X-100 micellar solution. J. Agric. Food Chem. 2017, 65, 784–792. [Google Scholar] [CrossRef] [PubMed]
- Bensasson, R.V.; Land, E.J. Intersystem crossing efficiencies of retinal isomers in different solvents measured by laser flash absorption spectrophotometry. Nouv. J. Chim. 1978, 2, 503–507. [Google Scholar]
- Bensasson, R.V.; Land, E.J.; Truscott, T.G. Flash Photolysis and Pulse Radiolysis; Pergamon Press: Oxford, UK, 1983; pp. 67–92. [Google Scholar]
- Dillon, J.; Gaillard, E.R.; Bilski, P.; Chignell, C.F.; Reszka, K.J. The photochemistry of the retinoids as studied by steady-state and pulsed methods. Photochem. Photobiol. 1996, 63, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Becker, R.S.; Freedman, K.; Lenoble, C. Photophysical and photochemical behavior of 11-cis-retinal and its Schiff base in a micelle. J. Phys. Chem. 1986, 90, 4334–4336. [Google Scholar] [CrossRef]
- Ferguson, J.; Johnson, B.E. Photosensitivity due to retinoids: Clinical and laboratory. Br. J. Dermatol. 1986, 115, 275–283. [Google Scholar] [CrossRef]
- Lo, K.K.N.; Land, E.J.; Truscott, T.G. Primary intermediates in the pulsed irradiation of retinoids. Photochem. Photobiol. 1982, 36, 139–145. [Google Scholar] [CrossRef]
- Tolleson, W.H.; Cherng, S.-H.; Xia, Q.; Boudreau, M.; Yin, J.J.; Wamer, W.G.; Howard, P.C.; Yu, H.; Fu, P.P. Photodecomposition and phototoxicity of natural retinoids. Int. J. Environ. Res. Public Health 2005, 2, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, T.; Alchalal, A.; Ottolenghi, M. Primary photoprocesses in retinol. Chem. Phys. Lett. 1973, 20, 291–297. [Google Scholar] [CrossRef]
- Collins, C.M.; Leventis, N.; Sotiriou-Leventis, C. Relative reactivity of vitamin A versus a mixture of β-carotene geometric isomers with electrochemically generated superoxide and hydroperoxyl radicals. Electrochimica Acta 2001, 47, 567–576. [Google Scholar] [CrossRef]
- Różanowska, M.; Cantrell, A.; Edge, R.; Land, E.J.; Sarna, T.; Truscott, T.G. Pulse radiolysis study of the interaction of retinoids with peroxyl radicals. Free Radic. Biol. Med. 2005, 39, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Böhm, F.; Edge, R.; Land, E.J.; McGarvey, D.J.; Truscott, T.G. Carotenoids enhance vitamin E antioxidant efficiency. J. Am. Chem. Soc. 1997, 119, 621–622. [Google Scholar] [CrossRef]
- El-Agamey, A.; Fukuzumi, S.; Naqvi, K.R.; McGarvey, D.J. Kinetic studies of retinol addition radicals. Org. Biomol. Chem. 2011, 9, 1459–1465. [Google Scholar] [CrossRef] [PubMed]
- Aveline, B.M.; Kochevar, I.E.; Redmond, R.W. Photochemistry of the nonspecific hydroxyl radical generator, N-hydroxypyridine-2(1H)-thione. J. Am. Chem. Soc. 1996, 118, 10113–10123. [Google Scholar] [CrossRef]
- El-Agamey, A.; McGarvey, D.J. First direct observation of reversible oxygen addition to a carotenoid-derived carbon-centered neutral radical. Org. Lett. 2005, 7, 3957–3960. [Google Scholar] [CrossRef] [PubMed]
- El-Agamey, A.; McGarvey, D.J. The reactivity of carotenoid radicals with oxygen. Free Radic. Res. 2007, 41, 295–302. [Google Scholar] [CrossRef] [PubMed]
- El-Agamey, A.; Fukuzumi, S. Laser flash photolysis study on the retinol radical cation in polar solvents. Org. Biomol. Chem. 2011, 9, 6437–6446. [Google Scholar] [CrossRef] [PubMed]
- El-Agamey, A.; Melø, T.B.; Sliwka, H.-R. Exploring the reactivity of retinol radical cation toward organic and biological molecules: A laser flash photolysis study. J. Photochem. Photobiol. B Biol. 2017, 170, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Gorman, A.A.; Rodgers, M.A.J. Singlet molecular oxygen. Chem. Soc. Rev. 1981, 10, 205–231. [Google Scholar] [CrossRef]
- Piette, J. Biological consequences associated with DNA oxidation mediated by singlet oxygen. J. Photochem. Photobiol. 1991, 11, 241–260. [Google Scholar] [CrossRef]
- Girotti, A.W. Photodynamic lipid peroxidation in biological systems. Photochem. Photobiol. 1990, 51, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Minami, Y.; Yokoyama, K.; Bando, N.; Kawai, Y.; Terao, J. Occurance of singlet oxygen oxygenation of oleic acid and linoleic acid in the skin of live mice. Free Radic. Res. 2008, 42, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J.; Truscott, R.J.W. Photo-oxidation of proteins and its role in cataractogenesis. J. Photochem. Photobiol. B Biol. 2001, 63, 114–125. [Google Scholar] [CrossRef]
- Badger, R.M.; Wright, A.C.; Whitlock, R.F. Absolute intensities of the discrete and continuous absorption bands of oxygen gas at 1.26 and 1.065 μ and the radiative lifetime of the 1Δg state of oxygen. J. Chem. Phys. 1965, 43, 4345–4350. [Google Scholar] [CrossRef]
- Foote, C.S.; Denny, R.W. Chemistry of singlet oxygen. VII. Quenching by β-carotene. J. Am. Chem. Soc. 1968, 90, 6233–6235. [Google Scholar] [CrossRef]
- Farmilo, A.; Wilkinson, F. On the mechanism of quenching of singlet oxygen in solution. Photochem. Photobiol. 1973, 18, 447–450. [Google Scholar] [CrossRef] [PubMed]
- Fiedor, J.; Fiedor, L.; Haeβner, R.; Scheer, H. Cyclic endoperoxides of β-carotene, potential pro-oxidants, as products of chemical quenching of singlet oxygen. Biochim. Biophys. Acta 2005, 1709, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Nishino, A.; Yasui, H.; Maoka, T. Reaction and scavenging mechanism of β-carotene and zeaxanthin with reactive oxygen species. J. Oleo Sci. 2017, 66, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Liebler, D.C. Antioxidant reactions of carotenoids. Ann. N. Y. Acad. Sci. 1993, 691, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Oliveros, E.; Braun, A.M.; Aminian-Saghafi, T.; Sliwka, H.R. Quenching of singlet oxygen (1Δg) by carotenoid derivatives: Kinetic analysis by near infra-red luminescence. New J. Chem. 1994, 18, 535–539. [Google Scholar]
- Edge, R.; McGarvey, D.J.; Truscott, T.G. The carotenoids as antioxidants—A review. J. Photochem. Photobiol. B Biol. 1997, 41, 189–200. [Google Scholar] [CrossRef]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef]
- Weedon, B.C.L.; Moss, C.P. Structure and nomenclature. In Carotenoids: Isolation and Analysis; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser: Basel, Switzerland, 1995; Volume 1A, pp. 27–70. [Google Scholar]
- Cantrell, A.; McGarvey, D.J.; Truscott, T.G. Singlet oxygen quenching by dietary carotenoids in a model membrane environment. Arch. Biochem. Biophys. 2003, 412, 47–54. [Google Scholar] [CrossRef]
- Nishino, A.; Yasui, H.; Maoka, T. Reaction of paprika carotenoids, capsanthin and capsrubin, with reactive oxygen species. J. Agric. Food Chem. 2016, 64, 4786–4792. [Google Scholar] [CrossRef] [PubMed]
- Burke, M. Pulsed Radiation Studies of Carotenoid Radicals and Excited States. Ph.D Thesis, Keele University, Staffordshire, UK, April 2001. [Google Scholar]
- Edge, R.; Truscott, T.G. Properties of carotenoid radicals and excited states and their potential role in biological systems. In Carotenoids. Physical, Chemical, and Biological Functions and Properties; Landrum, J.T., Ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 283–307. [Google Scholar]
- Teraoka, J.; Hashimoto, H.; Matsudaira, S.; Koyama, Y. Resonance Raman spectra of excited triplet states of β-carotene isomers. Chem. Lett. 1985, 14, 311–314. [Google Scholar] [CrossRef]
- Heymann, T.; Heinz, P.; Glomb, M.A. Lycopene inhibits the isomerization of β-carotene during quenching of singlet oxygen and free radicals. J. Agric. Food. Chem. 2015, 63, 3279–3287. [Google Scholar] [CrossRef] [PubMed]
- Fukuzawa, K. Singlet oxygen scavenging in phospholipid membranes. Methods Enzymol. 2000, 319, 101–110. [Google Scholar] [PubMed]
- Fukuzawa, K.; Inokami, Y.; Tokumura, A.; Terao, J.; Suzuki, A. Rate constants for quenching singlet oxygen and activities for inhibiting lipid peroxidation of carotenoids and α-tocopherol in liposomes. Lipids 1998, 33, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Naito, Y.; Yoshikawa, T.; Niki, E. Rapid assessment of singlet oxygen-induced plasma lipid oxidation and its inhibition by antioxidants with diphenyl-1-pyrenylphosphine (DPPP). Anal. Bioanal. Chem. 2016, 408, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Sachindra, N.M.; Sato, E.; Maeda, H.; Hosokawa, M.; Niwano, Y.; Kohno, M.; Miyashita, K. Radical scavenging and singlet oxygen quenching activity of marine carotenoid fucoxanthin and its metabolites. J. Agric. Food Chem. 2007, 55, 8516–8522. [Google Scholar] [CrossRef] [PubMed]
- Telfer, A.; Dhami, S.; Bishop, S.M.; Phillips, D.; Barber, J. β-Carotene quenches singlet oxygen formed in isolated photosystem II reaction centers. Biochemistry 1994, 33, 14469–14474. [Google Scholar] [CrossRef] [PubMed]
- Böhm, F.; Haley, J.; Truscott, T.G.; Schalch, W. Cellular bound β-carotene quenches singlet oxygen in man. J. Photochem. Photobiol. B Biol. 1993, 21, 219–221. [Google Scholar] [CrossRef]
- Tinkler, J.H.; Böhm, F.; Schalch, W.; Truscott, T.G. Dietary carotenoids protect human cells from damage. J. Photochem. Photobiol. B Biol. 1994, 26, 283–285. [Google Scholar] [CrossRef]
- Boehm, F.; Edge, R.; Burke, M.; Truscott, T.G. Dietary uptake of lycopene protects human cells from singlet oxygen and nitrogen dioxide—ROS components from cigarette smoke. J. Photochem. Photobiol. B Biol. 2001, 64, 176–178. [Google Scholar] [CrossRef]
- Bosio, G.N.; Breitenbach, T.; Parisi, J.; Reigosa, M.; Blaikie, F.H.; Pedersen, B.W.; Silva, E.F.F.; Martire, D.O.; Ogilvy, P.R. Antioxidant β-carotene does not quench singlet oxygen in mammalian cells. J. Am. Chem. Soc. 2013, 135, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, E.F.F.; Pedersen, B.W.; Breitenbach, T.; Toftegaard, R.; Kuimova, M.K.; Arnaut, L.G.; Ogilvy, P.R. Irradiation- and sensitizer-dependent changes in the lifetime of intracellular singlet oxygen produced in a photosensitized process. J. Phys. Chem. B 2012, 116, 445–461. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, F.; Helman, W.P.; Ross, A.B. Rate constants for the decay and reactions of the lowest electronically excited singlet state of molecular oxygen in solution. An expanded and revised compilation. J. Phys. Chem. Ref. Data 1995, 24, 663–1021. [Google Scholar] [CrossRef]
- Böhm, F.; Edge, R.; Truscott, T.G. Interactions of dietary carotenoids with activated (singlet) oxygen and free radicals: Potential effects for human health. Mol. Nutr. Food Res. 2012, 56, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, R.; Vaz, B.; Gronemayer, H.; de Lera, A.R. Functions, therapeutic applications, and synthesis of retinoids and carotenoids. Chem. Rev. 2014, 114, 1–125. [Google Scholar] [CrossRef] [PubMed]
- Boehm, F.; Edge, R.; Truscott, T.G.; Witt, C. Photoprotection and radiation protection by dietary carotenoids. In Carotenoids—Nutrition, Analysis and Technology; Kaczor, A., Baranska, M., Eds.; John Wiley & Sons: Chichester, UK, 2016; pp. 43–58. [Google Scholar]
- Land, E.J.; Lafferty, J.; Roach, A.C.; Sinclair, R.S.; Truscott, T.G. Absorption spectra of radical ions of polyenes of biological interest. J. Chem. Soc. Faraday Trans. 1977, 73, 416–429. [Google Scholar]
- Edge, R.; Land, E.J.; McGarvey, D.J.; Burke, M.; Truscott, T.G. The reduction potential of the β-carotene•+/β-carotene couple in an aqueous micro-heterogeneous environment. FEBS Lett. 2000, 471, 125–127. [Google Scholar] [CrossRef]
- Burke, M.; Edge, R.; Land, E.J.; McGarvey, D.J.; Truscott, T.G. One-electron reduction potentials of dietary carotenoid radical cations in aqueous micellar environments. FEBS Lett. 2001, 500, 132–136. [Google Scholar] [CrossRef]
- Tinkler, J.H.; Tavender, S.M.; Parker, A.W.; McGarvey, D.J.; Mulroy, L.; Truscott, T.G. An investigation of carotenoid radical cations and triplet states by laser flash photolysis and time-resolved resonance Raman spectroscopy: Observation of competitive energy and electron transfer. J. Am. Chem. Soc. 1996, 118, 1756–1761. [Google Scholar] [CrossRef]
- Cheng, H.; Han, R.-M.; Lyu, M.-K.; Zhang, J.-P.; Skibsted, L.H. Regeneration of β-carotene from the radical cation by tyrosine and tryptophan. J. Phys. Chem. B 2015, 119, 6603–6610. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-T.; Cheng, H.; Han, R.-M.; Wang, P.; Zhang, J.-P.; Skibsted, L.H. Regeneration of β-carotene from radical cation by eugenol, isoeugenol, and clove oil in the Marcus theory inverted region for electron transfer. J. Agric. Food Chem. 2017, 65, 908–912. [Google Scholar] [CrossRef] [PubMed]
- Mairanovsky, V.G.; Engovatov, A.A.; Ioffe, N.T.; Samokhvalov, G.I. Electron-donor and electron-acceptor properties of carotenoids: Electrochemical study of carotenes. J. Electroanal. Chem. 1975, 66, 122–137. [Google Scholar] [CrossRef]
- Focsan, A.L.; Pan, S.; Kispert, L.D. Electrochemical study of astaxanthin and astaxanthin n-octanoic monoester and diester: Tendency to form radicals. J. Phys. Chem. B 2014, 118, 2331–3229. [Google Scholar] [CrossRef] [PubMed]
- Focsan, A.L.; Magyar, A.; Kispert, L.D. Chemistry of carotenoid neutral radicals. Arch. Biochem. Biophys. 2015, 572, 167–174. [Google Scholar] [CrossRef] [PubMed]
- El-Agamey, A.; El-Hagrasy, M.A.; Suenobu, T.; Fukuzumi, S. Influence of pH on the decay of β-carotene radical cation in aqueous Triton X-100: A laser flash photolysis study. J. Photochem. Photobiol. B Biol. 2015, 146, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Focsan, A.L.; Kispert, L.D. Radicals formed from proton loss of carotenoid radical cations: A special form of carotenoid neutral radical occurring in photoprotection. J. Photochem. Photobiol. B Biol. 2017, 166, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Shinopoulos, K.E.; Tracewell, C.A.; Focsan, A.L.; Brudvig, G.W.; Kispert, L.D. Formation of carotenoid neutral radicals in Photosystem II. J. Phys. Chem. B 2009, 113, 9901–9908. [Google Scholar] [CrossRef] [PubMed]
- Magyar, A.; Bowman, M.K.; Molnar, P.; Kiapert, L. Neutral carotenoid radicals in photoprotection of wild-type Arabidopsis thaliana. J. Phys. Chem. B 2013, 117, 2239–2246. [Google Scholar] [CrossRef] [PubMed]
- El-Agamey, A.; McGarvey, D.J. Peroxyl radical reactions with carotenoids in microemulsions: Influence of microemulsion composition and the nature of peroxyl radical precursor. Free Radic. Biol. Med. 2016, 90, 75–84. [Google Scholar] [CrossRef] [PubMed]
- El-Agamey, A.; Edge, R.; Navaratnam, S.; Land, E.J.; Truscott, T.G. Carotenoid radical anions and their protonated derivatives. Org. Lett. 2006, 8, 4255–4258. [Google Scholar] [CrossRef] [PubMed]
- Everett, S.A.; Patel, K.B.; Maddix, S.; Kundu, S.C.; Willson, R.L. Scavenging of nitrogen dioxide, thiyl, and sulfonyl free radicals by the nutritional antioxidant β-carotene. J. Biol. Chem. 1996, 271, 3988–3994. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, A.; Skibsted, L.H. Kinetics of photobleaching of β-carotene in chloroform and formation of transient carotenoid species absorbing in the near infrared. Free Radic. Res. 1996, 25, 355–368. [Google Scholar] [CrossRef]
- Koppenol, W.H. Generation and thermodynamic properties of oxyradicals. In CRC Critical Reviews in Membrane Lipid Oxidation, 2nd ed.; Vigo-Pelfrety, C., Ed.; CRC Press: Boca Raton, FL, USA, 1989; Volume 1, pp. 1–13. [Google Scholar]
- Chen, C.-H.; Han, R.-M.; Liang, R.; Fu, L.-M.; Wang, P.; Ai, X.-C.; Zhang, J.-P.; Skibsted, L.H. Direct observation of the β-carotene reaction with hydroxyl radical. J. Phys. Chem. B 2011, 115, 2082–2089. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.W.; Ingold, K.U. beta-Carotene: An unusual type of lipid antioxidant. Science 1984, 224, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Boehm, F.; Edge, R.; Truscott, T.G.; Witt, C. A dramatic effect of oxygen on protection of human cells against γ-radiation by lycopene. FEBS Lett. 2016, 590, 1086–1093. [Google Scholar] [CrossRef] [PubMed]
- Mukai, K.; Yoshimoto, M.; Ishikura, M.; Nagaoka, S. Kinetic study of the aroxyl-radical-scavenging activity of five fatty acid esters and six carotenoids in toluene solution: Structure-activity relationship for the hydrogen abstraction reaction. J. Phys. Chem. B 2017, 121, 7593–7601. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| kq/109 dm3 mol−1 s−1 | |||||
|---|---|---|---|---|---|
| Carotenoid | DPPC Liposomes [39] | Micelles [36] | Benzene [36] | Ethanol:Chloroform:Water 50:50:1 [37] | |
| * | † | ||||
| Lycopene | 2.4 | 2.3 | 2.0 | 17.0 | 31.0 |
| β-Carotene | 2.3 | 2.5 | 2.4 | 13.0 | 14.0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Edge, R.; Truscott, T.G. Singlet Oxygen and Free Radical Reactions of Retinoids and Carotenoids—A Review. Antioxidants 2018, 7, 5. https://doi.org/10.3390/antiox7010005
Edge R, Truscott TG. Singlet Oxygen and Free Radical Reactions of Retinoids and Carotenoids—A Review. Antioxidants. 2018; 7(1):5. https://doi.org/10.3390/antiox7010005
Chicago/Turabian StyleEdge, Ruth, and T. George Truscott. 2018. "Singlet Oxygen and Free Radical Reactions of Retinoids and Carotenoids—A Review" Antioxidants 7, no. 1: 5. https://doi.org/10.3390/antiox7010005
APA StyleEdge, R., & Truscott, T. G. (2018). Singlet Oxygen and Free Radical Reactions of Retinoids and Carotenoids—A Review. Antioxidants, 7(1), 5. https://doi.org/10.3390/antiox7010005