Crystal Structure of Chloroplastic Thioredoxin f2 from Chlamydomonas reinhardtii Reveals Distinct Surface Properties

 , ,
, , 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cloning, Expression, and Purification of CrTRXf2 and CrTRXh1
2.2. Crystallization and Diffraction Data Collection
2.3. Structure Determination, Model Building, and Analysis
2.4. Circular Dichroism (CD) Spectroscopy
3. Results
3.1. Sequence Analysis of Chlamydomonas TRX f2
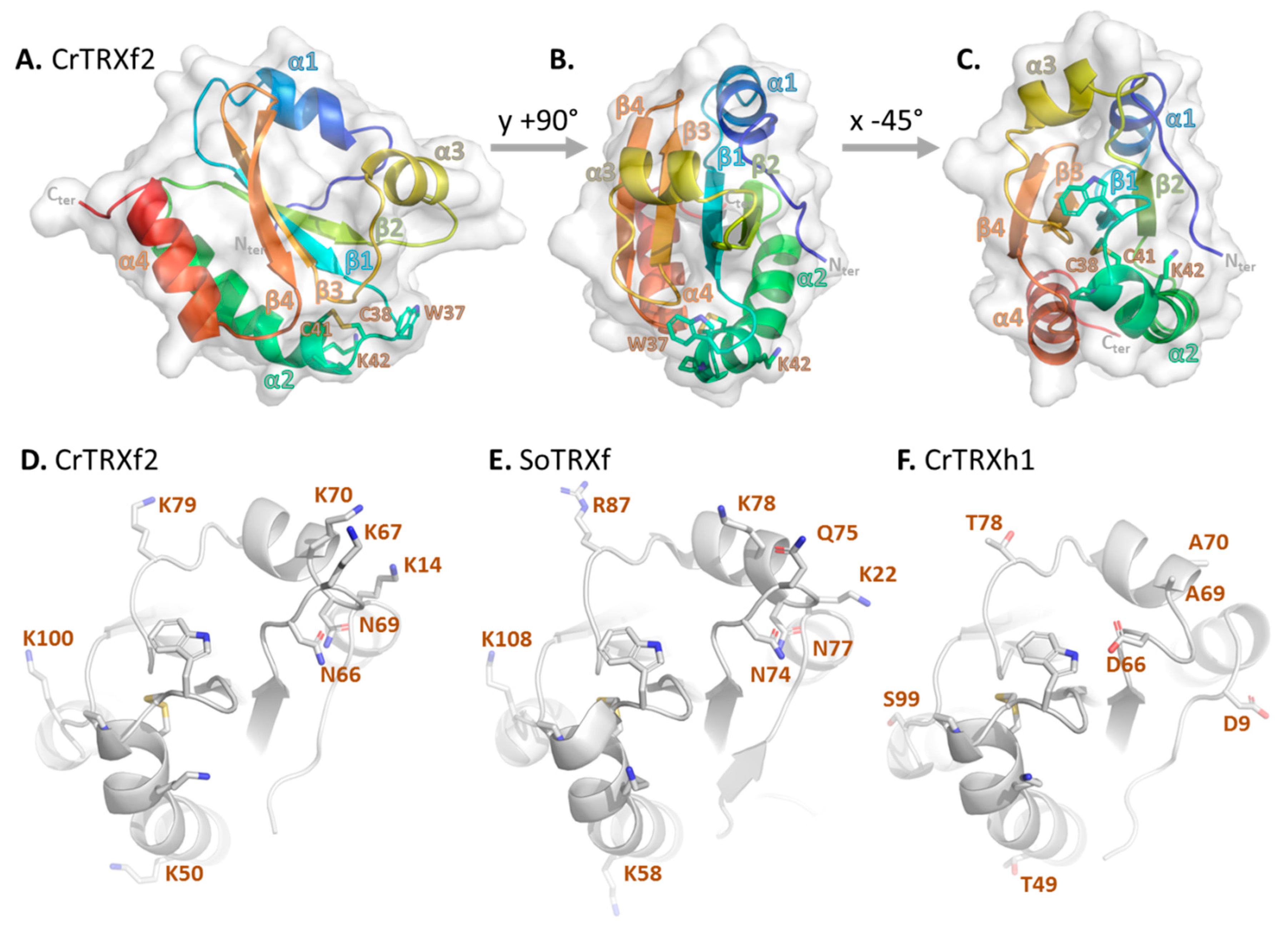
3.2. Chlamydomonas TRX f2 Folds as a Canonical TRX
3.3. The Redox Site of CrTRXf2
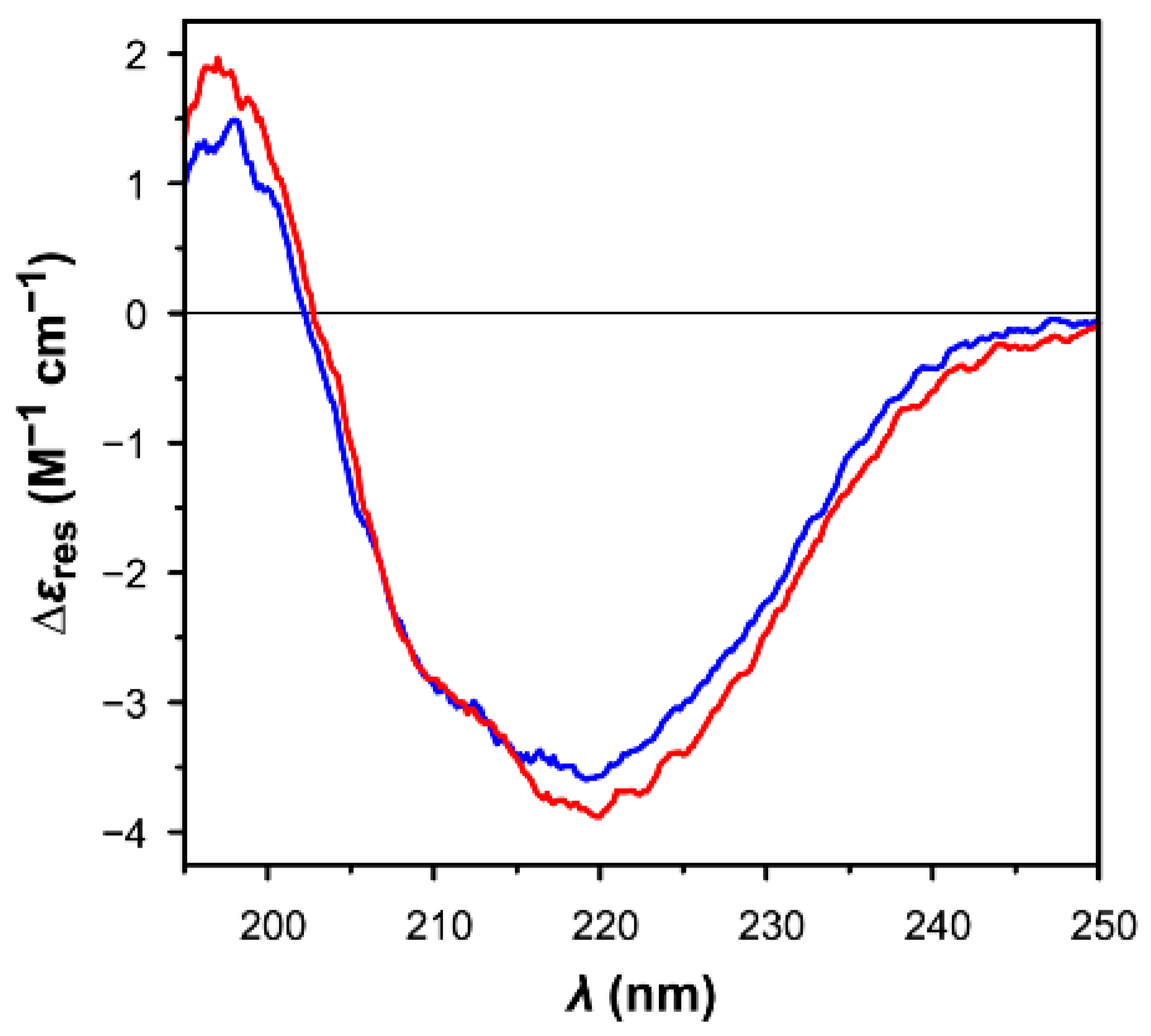
3.4. Comparison of the Secondary Structures of Oxidized and Reduced CrTRXf2
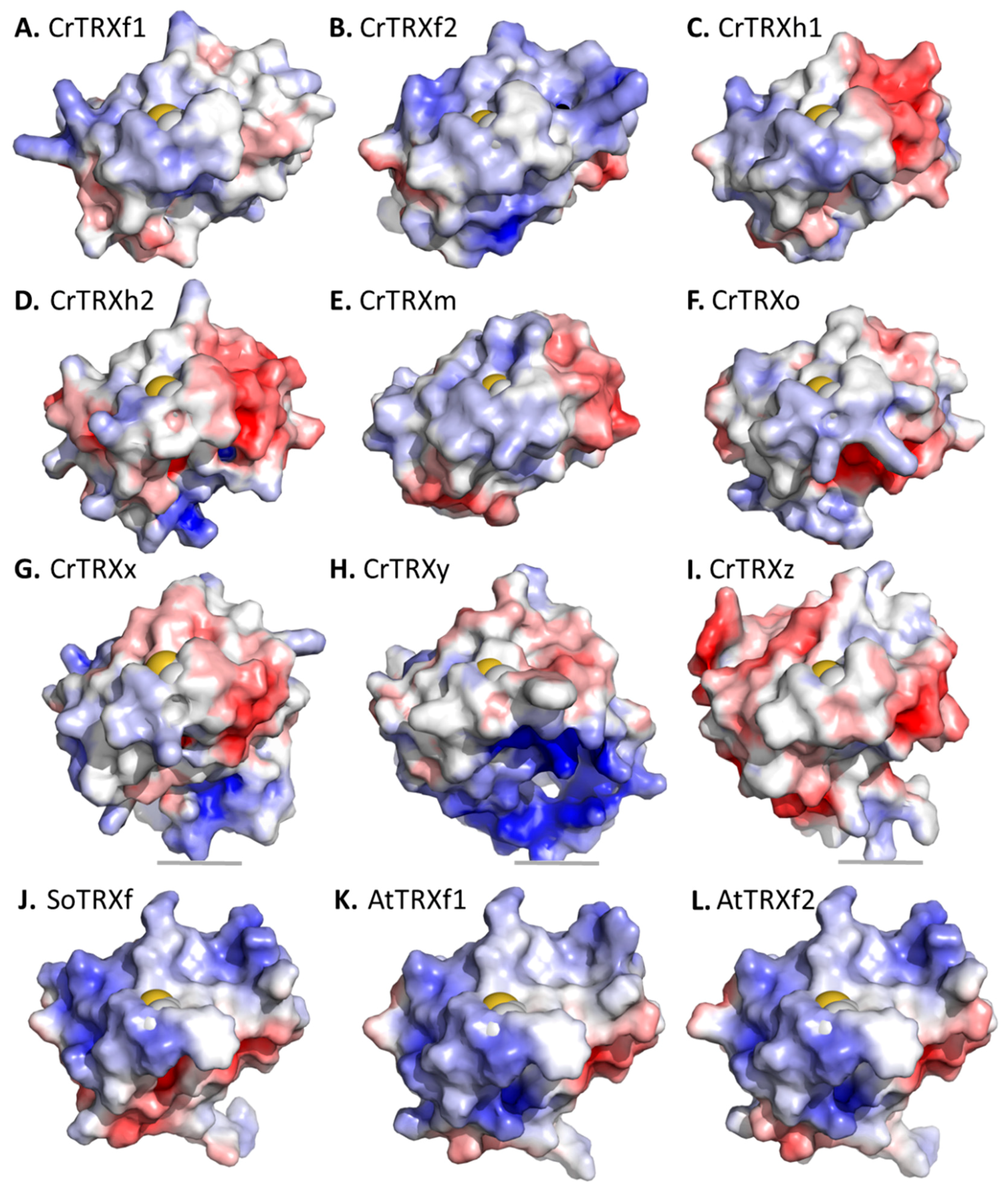
3.5. Surface Specificities of CrTRXf2
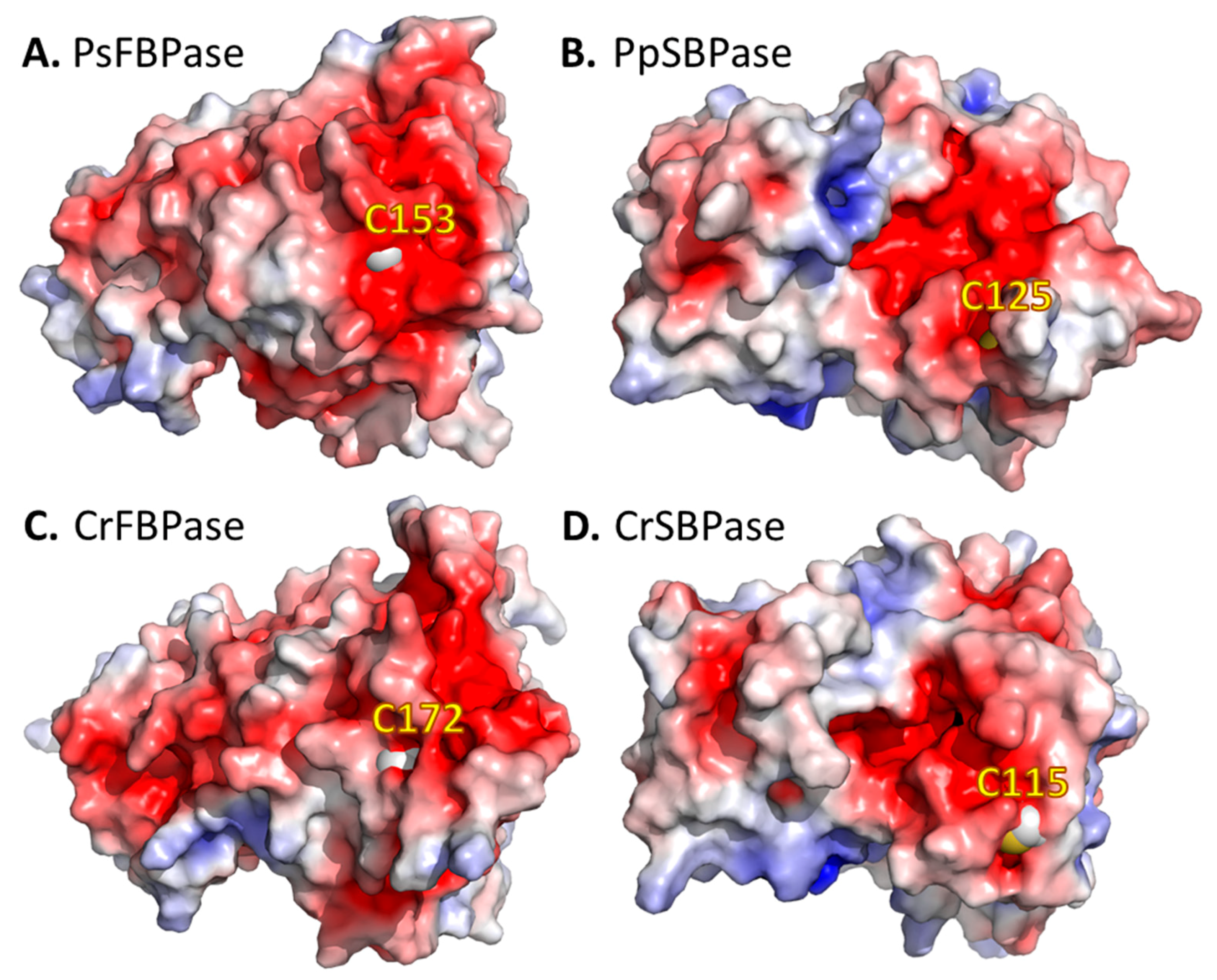
3.6. Surface Specificities of TRXf Targets
4. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Martin, J.L. Thioredoxin—A fold for all reasons. Structure 1995, 3, 245–250. [Google Scholar] [CrossRef]
- Katti, S.K.; LeMaster, D.M.; Eklund, H. Crystal structure of thioredoxin from Escherichia coli at 1.68 A resolution. J. Mol. Biol. 1990, 212, 167–184. [Google Scholar] [CrossRef]
- Dyson, H.J.; Gippert, G.P.; Case, D.A.; Holmgren, A.; Wright, P.E. Three-dimensional solution structure of the reduced form of Escherichia coli thioredoxin determined by nuclear magnetic resonance spectroscopy. Biochemistry 1990, 29, 4129–4136. [Google Scholar] [CrossRef] [PubMed]
- Forman-Kay, J.D.; Clore, G.M.; Wingfield, P.T.; Gronenborn, A.M. High-resolution three-dimensional structure of reduced recombinant human thioredoxin in solution. Biochemistry 1991, 30, 2685–2698. [Google Scholar] [CrossRef] [PubMed]
- Collet, J.F.; Messens, J. Structure, function, and mechanism of thioredoxin proteins. Antioxid. Redox Signal. 2010, 13, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, B.B.; Holmgren, A.; Jacquot, J.P.; Scheibe, R. Fifty years in the thioredoxin field and a bountiful harvest. Biochim. Biophys. Acta 2012, 1820, 1822–1829. [Google Scholar] [CrossRef] [PubMed]
- Hanschmann, E.M.; Godoy, J.R.; Berndt, C.; Hudemann, C.; Lillig, C.H. Thioredoxins, glutaredoxins, and peroxiredoxins—Molecular mechanisms and health significance: From cofactors to antioxidants to redox signaling. Antioxid. Redox Signal. 2013, 19, 1539–1605. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, S.M.; Lee, R.T. Thioredoxin and thioredoxin target proteins: From molecular mechanisms to functional significance. Antioxid. Redox Signal. 2013, 18, 1165–1207. [Google Scholar] [CrossRef] [PubMed]
- Toledano, M.B.; Delaunay-Moisan, A.; Outten, C.E.; Igbaria, A. Functions and cellular compartmentation of the thioredoxin and glutathione pathways in yeast. Antioxid. Redox Signal. 2013, 18, 1699–1711. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, S.D.; Michelet, L.; Zaffagnini, M.; Massot, V.; Issakidis-Bourguet, E. Thioredoxins in chloroplasts. Curr. Genet. 2007, 51, 343–365. [Google Scholar] [CrossRef] [PubMed]
- Meyer, Y.; Belin, C.; Delorme-Hinoux, V.; Reichheld, J.P.; Riondet, C. Thioredoxin and glutaredoxin systems in plants: Molecular mechanisms, crosstalks, and functional significance. Antioxid. Redox Signal. 2012, 17, 1124–1160. [Google Scholar] [CrossRef] [PubMed]
- Serrato, A.J.; Fernandez-Trijueque, J.; Barajas-Lopez, J.D.; Chueca, A.; Sahrawy, M. Plastid thioredoxins: A “one-for-all” redox-signaling system in plants. Front. Plant Sci. 2013, 4, 463. [Google Scholar] [CrossRef] [PubMed]
- Geigenberger, P.; Thormahlen, I.; Daloso, D.M.; Fernie, A.R. The Unprecedented Versatility of the Plant Thioredoxin System. Trendsplant Sci. 2017, 22, 249–262. [Google Scholar]
- Balsera, M.; Uberegui, E.; Schurmann, P.; Buchanan, B.B. Evolutionary development of redox regulation in chloroplasts. Antioxid. Redox Signal. 2014, 21, 1327–1355. [Google Scholar] [CrossRef] [PubMed]
- Jacquot, J.P.; Eklund, H.; Rouhier, N.; Schurmann, P. Structural and evolutionary aspects of thioredoxin reductases in photosynthetic organisms. Trends Plant Sci. 2009, 14, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Michelet, L.; Zaffagnini, M.; Morisse, S.; Sparla, F.; Perez-Perez, M.E.; Francia, F.; Danon, A.; Marchand, C.H.; Fermani, S.; Trost, P.; et al. Redox regulation of the Calvin-Benson cycle: Something old, something new. Front. Plant Sci. 2013, 4, 470. [Google Scholar] [CrossRef] [PubMed]
- Schurmann, P.; Buchanan, B.B. The ferredoxin/thioredoxin system of oxygenic photosynthesis. Antioxid. Redox Signal. 2008, 10, 1235–1274. [Google Scholar] [CrossRef] [PubMed]
- Wolosiuk, R.A.; Buchanan, B.B. Thioredoxin and glutathione regulate photosynthesis in chloroplasts. Nature 1977, 266, 565–567. [Google Scholar] [CrossRef]
- Jacquot, J.-P.; Vidal, J.; Gadal, P.; Schürmann, P. Evidence for the existence of several enzyme-specific thioredoxins in plants. FEBS Lett. 1978, 96, 243–246. [Google Scholar] [CrossRef] [Green Version]
- Scheibe, R.; Anderson, L.E. Dark modulation of NADP-dependent malate dehydrogenase and glucose-6-phosphate dehydrogenase in the chloroplast. Biochim. Biophys. Acta 1981, 636, 58–64. [Google Scholar] [CrossRef]
- Vaseghi, M.J.; Chibani, K.; Telman, W.; Liebthal, M.F.; Gerken, M.; Schnitzer, H.; Mueller, S.M.; Dietz, K.J. The chloroplast 2-cysteine peroxiredoxin functions as thioredoxin oxidase in redox regulation of chloroplast metabolism. eLife 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Ojeda, V.; Perez-Ruiz, J.M.; Cejudo, F.J. 2-Cys Peroxiredoxins Participate in the Oxidation of Chloroplast Enzymes in the Dark. Mol. Plant 2018, 11, 1377–1388. [Google Scholar] [CrossRef] [PubMed]
- Perez-Perez, M.E.; Mauries, A.; Maes, A.; Tourasse, N.J.; Hamon, M.; Lemaire, S.D.; Marchand, C.H. The Deep Thioredoxome in Chlamydomonas reinhardtii: New Insights into Redox Regulation. Mol. Plant 2017, 10, 1107–1125. [Google Scholar] [CrossRef] [PubMed]
- Ingles-Prieto, A.; Ibarra-Molero, B.; Delgado-Delgado, A.; Perez-Jimenez, R.; Fernandez, J.M.; Gaucher, E.A.; Sanchez-Ruiz, J.M.; Gavira, J.A. Conservation of protein structure over four billion years. Structure 2013, 21, 1690–1697. [Google Scholar] [CrossRef] [PubMed]
- Mouaheb, N.; Thomas, D.; Verdoucq, L.; Monfort, P.; Meyer, Y. In vivo functional discrimination between plant thioredoxins by heterologous expression in the yeast Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1998, 95, 3312–3317. [Google Scholar] [CrossRef] [PubMed]
- Issakidis-Bourguet, E.; Mouaheb, N.; Meyer, Y.; Miginiac-Maslow, M. Heterologous complementation of yeast reveals a new putative function for chloroplast m-type thioredoxin. Plant J. 2001, 25, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Morisse, S.; Michelet, L.; Bedhomme, M.; Marchand, C.H.; Calvaresi, M.; Trost, P.; Fermani, S.; Zaffagnini, M.; Lemaire, S.D. Thioredoxin-dependent redox regulation of chloroplastic phosphoglycerate kinase from Chlamydomonas reinhardtii. J. Biol. Chem. 2014, 289, 30012–30024. [Google Scholar] [CrossRef] [PubMed]
- Huppe, H.C.; de Lamotte-Guery, F.; Jacquot, J.P.; Buchanan, B.B. The ferredoxin-thioredoxin system of a green alga, Chlamydomonas reinhardtii: Identification and characterization of thioredoxins and ferredoxin-thioredoxin reductase components. Planta 1990, 180, 341–351. [Google Scholar] [PubMed]
- Collin, V.; Issakidis-Bourguet, E.; Marchand, C.; Hirasawa, M.; Lancelin, J.M.; Knaff, D.B.; Miginiac-Maslow, M. The Arabidopsis plastidial thioredoxins: New functions and new insights into specificity. J. Biol. Chem. 2003, 278, 23747–23752. [Google Scholar] [CrossRef] [PubMed]
- Collin, V.; Lamkemeyer, P.; Miginiac-Maslow, M.; Hirasawa, M.; Knaff, D.B.; Dietz, K.J.; Issakidis-Bourguet, E. Characterization of plastidial thioredoxins from Arabidopsis belonging to the new y-type. Plant Physiol. 2004, 136, 4088–4095. [Google Scholar] [CrossRef] [PubMed]
- Schurmann, P.; Buchanan, B.B. Role of ferredoxin in the activation of sedoheptulose diphosphatase in isolated chloroplasts. Biochim. Biophys. Acta 1975, 376, 189–192. [Google Scholar] [CrossRef]
- Gutle, D.D.; Roret, T.; Muller, S.J.; Couturier, J.; Lemaire, S.D.; Hecker, A.; Dhalleine, T.; Buchanan, B.B.; Reski, R.; Einsle, O.; et al. Chloroplast FBPase and SBPase are thioredoxin-linked enzymes with similar architecture but different evolutionary histories. Proc. Natl. Acad. Sci. USA 2016, 113, 6779–6784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marri, L.; Zaffagnini, M.; Collin, V.; Issakidis-Bourguet, E.; Lemaire, S.D.; Pupillo, P.; Sparla, F.; Miginiac-Maslow, M.; Trost, P. Prompt and easy activation by specific thioredoxins of calvin cycle enzymes of Arabidopsis thaliana associated in the GAPDH/CP12/PRK supramolecular complex. Mol. Plant 2009, 2, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Trost, P.; Fermani, S.; Marri, L.; Zaffagnini, M.; Falini, G.; Scagliarini, S.; Pupillo, P.; Sparla, F. Thioredoxin-dependent regulation of photosynthetic glyceraldehyde-3-phosphate dehydrogenase: Autonomous vs. CP12-dependent mechanisms. Photosynthesis Res. 2006, 89, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Fermani, S.; Sparla, F.; Marri, L.; Thumiger, A.; Pupillo, P.; Falini, G.; Trost, P. Structure of photosynthetic glyceraldehyde-3-phosphate dehydrogenase (isoform A4) from Arabidopsis thaliana in complex with NAD. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66 Pt 6, 621–626. [Google Scholar] [CrossRef]
- Wolosiuk, R.A.; Buchanan, B.B. Regulation of chloroplast phosphoribulokinase by the ferredoxin/thioredoxin system. Arch. Biochem. Biophys. 1978, 189, 97–101. [Google Scholar] [CrossRef]
- Setterdahl, A.T.; Chivers, P.T.; Hirasawa, M.; Lemaire, S.D.; Keryer, E.; Miginiac-Maslow, M.; Kim, S.K.; Mason, J.; Jacquot, J.P.; Longbine, C.C.; et al. Effect of pH on the oxidation-reduction properties of thioredoxins. Biochemistry 2003, 42, 14877–14884. [Google Scholar] [CrossRef] [PubMed]
- Hirasawa, M.; Schurmann, P.; Jacquot, J.P.; Manieri, W.; Jacquot, P.; Keryer, E.; Hartman, F.C.; Knaff, D.B. Oxidation-reduction properties of chloroplast thioredoxins, ferredoxin:thioredoxin reductase, and thioredoxin f-regulated enzymes. Biochemistry 1999, 38, 5200–5205. [Google Scholar] [CrossRef] [PubMed]
- Nikkanen, L.; Toivola, J.; Diaz, M.G.; Rintamaki, E. Chloroplast thioredoxin systems: Prospects for improving photosynthesis. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2017, 372, 20160474. [Google Scholar] [CrossRef] [PubMed]
- Emanuelsson, O.; Nielsen, H.; von Heijne, G. ChloroP, a neural network-based method for predicting chloroplast transit peptides and their cleavage sites. Protein Sci. 1999, 8, 978–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquini, M.; Fermani, S.; Tedesco, D.; Sciabolini, C.; Crozet, P.; Naldi, M.; Henri, J.; Vothknecht, U.; Bertucci, C.; Lemaire, S.D.; et al. Structural basis for the magnesium-dependent activation of transketolase from Chlamydomonas reinhardtii. Biochim. Biophys. Acta 2017, 1861, 2132–2145. [Google Scholar] [CrossRef] [PubMed]
- Pace, C.N.; Vajdos, F.; Fee, L.; Grimsley, G.; Gray, T. How to measure and predict the molar absorption coefficient of a protein. Protein Sci. 1995, 4, 2411–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaffagnini, M.; Michelet, L.; Massot, V.; Trost, P.; Lemaire, S.D. Biochemical characterization of glutaredoxins from Chlamydomonas reinhardtii reveals the unique properties of a chloroplastic CGFS-type glutaredoxin. J. Biol. Chem. 2008, 283, 8868–8876. [Google Scholar] [CrossRef] [PubMed]
- McWilliam, H.; Li, W.; Uludag, M.; Squizzato, S.; Park, Y.M.; Buso, N.; Cowley, A.P.; Lopez, R. Analysis Tool Web Services from the EMBL-EBI. Nucleic Acids Res. 2013, 41, W597–600. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Zwart, P.H.; Afonine, P.V.; Grosse-Kunstleve, R.W.; Hung, L.W.; Ioerger, T.R.; McCoy, A.J.; McKee, E.; Moriarty, N.W.; Read, R.J.; Sacchettini, J.C.; et al. Automated structure solution with the PHENIX suite. Methods Mol. Biol. 2008, 426, 419–435. [Google Scholar] [PubMed]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonine, P.V.; Grosse-Kunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. Sect. D Biol. Crystallogr. 2012, 68 Pt 4, 352–367. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66 Pt 4, 86–501. [Google Scholar]
- Menchise, V.; Corbier, C.; Didierjean, C.; Saviano, M.; Benedetti, E.; Jacquot, J.P.; Aubry, A. Crystal structure of the wild-type and D30A mutant thioredoxin h of Chlamydomonas reinhardtii and implications for the catalytic mechanism. Biochem. J. 2001, 359 Pt 1, 65–75. [Google Scholar] [CrossRef]
- Kinoshita, K.; Nakamura, H. eF-site and PDBjViewer: Database and viewer for protein functional sites. Bioinformatics 2004, 20, 1329–1330. [Google Scholar] [CrossRef] [PubMed]
- Micsonai, A.; Wien, F.; Kernya, L.; Lee, Y.H.; Goto, Y.; Refregiers, M.; Kardos, J. Accurate secondary structure prediction and fold recognition for circular dichroism spectroscopy. Proc. Natl. Acad. Sci. USA 2015, 112, E3095–E3103. [Google Scholar] [CrossRef] [PubMed]
- Micsonai, A.; Wien, F.; Bulyaki, E.; Kun, J.; Moussong, E.; Lee, Y.H.; Goto, Y.; Refregiers, M.; Kardos, J. BeStSel: A web server for accurate protein secondary structure prediction and fold recognition from the circular dichroism spectra. Nucl. Acids Res. 2018, 46, W315–W322. [Google Scholar] [CrossRef] [PubMed]
- Menchise, V.; Corbier, C.; Didierjean, C.; Jacquot, J.P.; Benedetti, E.; Saviano, M.; Aubry, A. Crystal structure of the W35A mutant thioredoxin h from Chlamydomonas reinhardtii: The substitution of the conserved active site Trp leads to modifications in the environment of the two catalytic cysteines. Biopolymers 2000, 56, 1–7. [Google Scholar] [CrossRef]
- De Lamotte-Guery, F.; Pruvost, C.; Minard, P.; Delsuc, M.A.; Miginiac-Maslow, M.; Schmitter, J.M.; Stein, M.; Decottignies, P. Structural and functional roles of a conserved proline residue in the alpha2 helix of Escherichia coli thioredoxin. Protein Eng. 1997, 10, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, A.; Gomes-Neto, F.; Myiamoto, C.A.; Valente, A.P.; Almeida, F.C. Dissection of the water cavity of yeast thioredoxin 1: The effect of a hydrophobic residue in the cavity. Biochemistry 2015, 54, 2429–2442. [Google Scholar] [CrossRef] [PubMed]
- Ladbury, J.E.; Wynn, R.; Hellinga, H.W.; Sturtevant, J.M. Stability of oxidized Escherichia coli thioredoxin and its dependence on protonation of the aspartic acid residue in the 26 position. Biochemistry 1993, 32, 7526–7530. [Google Scholar] [CrossRef] [PubMed]
- Bjornberg, O.; Maeda, K.; Svensson, B.; Hagglund, P. Dissecting molecular interactions involved in recognition of target disulfides by the barley thioredoxin system. Biochemistry 2012, 51, 9930–9939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchand, C.H.; Fermani, S.; Rossi, J.; Gurrieri, L.; Tedesco, D.; Henri, J.; Sparla, F.; Trost, P.; Lemaire, S.D.; Zaffagnini, M. Structural and biochemical insights into the reactivity of thioredoxin h1 from Chlamydomonas reinhardtii. Antioxidants 2018, 7. under review. [Google Scholar]
- Capitani, G.; Markovic-Housley, Z.; DelVal, G.; Morris, M.; Jansonius, J.N.; Schurmann, P. Crystal structures of two functionally different thioredoxins in spinach chloroplasts. J. Mol. Biol. 2000, 302, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Lancelin, J.M.; Guilhaudis, L.; Krimm, I.; Blackledge, M.J.; Marion, D.; Jacquot, J.P. NMR structures of thioredoxin m from the green alga Chlamydomonas reinhardtii. Proteins 2000, 41, 334–349. [Google Scholar] [CrossRef]
- Michelet, L.; Zaffagnini, M.; Marchand, C.; Collin, V.; Decottignies, P.; Tsan, P.; Lancelin, J.M.; Trost, P.; Miginiac-Maslow, M.; Noctor, G.; et al. Glutathionylation of chloroplast thioredoxin f is a redox signaling mechanism in plants. Proc. Natl. Acad. Sci. USA 2005, 102, 16478–16483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reutimann, H.; Straub, B.; Luisi, P.L.; Holmgren, A. A conformational study of thioredoxin and its tryptic fragments. J. Biol. Chem. 1981, 256, 6796–6803. [Google Scholar] [PubMed]
- Collet, J.F.; D’Souza, J.C.; Jakob, U.; Bardwell, J.C. Thioredoxin 2, an oxidative stress-induced protein, contains a high affinity zinc binding site. J. Biol. Chem. 2003, 278, 45325–45332. [Google Scholar] [CrossRef] [PubMed]
- Ado, K.; Taniguchi, Y. Pressure effects on the structure and function of human thioredoxin. Biochim. Biophys. Acta 2007, 1774, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Kabsch, W.; Sander, C. Dictionary of protein secondary structure: Pattern recognition of hydrogen-bonded and geometrical features. Biopolymers 1983, 22, 2577–2637. [Google Scholar] [CrossRef] [PubMed]
- Sikic, K.; Tomic, S.; Carugo, O. Systematic comparison of crystal and NMR protein structures deposited in the protein data bank. Open Biochem. J. 2010, 4, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Dawson, N.L.; Lewis, T.E.; Das, S.; Lees, J.G.; Lee, D.; Ashford, P.; Orengo, C.A.; Sillitoe, I. CATH: An expanded resource to predict protein function through structure and sequence. Nucl. Acids Res. 2017, 45, D289–D295. [Google Scholar] [CrossRef] [PubMed]
- Matthews, B.W.; Liu, L. A review about nothing: Are apolar cavities in proteins really empty? Protein Sci. 2009, 18, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Chiadmi, M.; Navaza, A.; Miginiac-Maslow, M.; Jacquot, J.P.; Cherfils, J. Redox signalling in the chloroplast: Structure of oxidized pea fructose-1,6-bisphosphate phosphatase. EMBO J. 1999, 18, 6809–6815. [Google Scholar] [CrossRef] [PubMed]
- Garzon, J.I.; Lopez-Blanco, J.R.; Pons, C.; Kovacs, J.; Abagyan, R.; Fernandez-Recio, J.; Chacon, P. FRODOCK: A new approach for fast rotational protein-protein docking. Bioinformatics 2009, 25, 2544–2551. [Google Scholar] [CrossRef] [PubMed]
- Maeda, K.; Hagglund, P.; Finnie, C.; Svensson, B.; Henriksen, A. Structural basis for target protein recognition by the protein disulfide reductase thioredoxin. Structure 2006, 14, 1701–1710. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.M.; 3rd Lemaire, S.D.; Jacquot, J.P.; Makhatadze, G.I. Difference in the mechanisms of the cold and heat induced unfolding of thioredoxin h from Chlamydomonas reinhardtii: Spectroscopic and calorimetric studies. Biochemistry 2000, 39, 11154–11162. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, S.D.; Richardson, J.M.; Goyer, A.; Keryer, E.; Lancelin, J.M.; Makhatadze, G.I.; Jacquot, J.P. Primary structure determinants of the pH- and temperature-dependent aggregation of thioredoxin. Biochim. Biophys. Acta 2000, 1476, 311–323. [Google Scholar] [CrossRef]
- Palde, P.B.; Carroll, K.S. A universal entropy-driven mechanism for thioredoxin-target recognition. Proc. Natl. Acad. Sci. USA 2015, 112, 7960–7965. [Google Scholar] [CrossRef] [PubMed]
- Balmer, Y.; Koller, A.; Val, G.D.; Schurmann, P.; Buchanan, B.B. Proteomics uncovers proteins interacting electrostatically with thioredoxin in chloroplasts. Photosynthesis Res. 2004, 79, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Crozet, P.; Navarro, F.J.; Willmund, F.; Mehrshahi, P.; Bakowski, K.; Lauersen, K.J.; Perez-Perez, M.E.; Auroy, P.; Gorchs Rovira, A.; Sauret-Gueto, S.; et al. Birth of a Photosynthetic Chassis: A MoClo Toolkit Enabling Synthetic Biology in the Microalga Chlamydomonas reinhardtii. ACS Synth. Biol. 2018, 7, 2074–2086. [Google Scholar] [CrossRef] [PubMed]
- Greiner, A.; Kelterborn, S.; Evers, H.; Kreimer, G.; Sizova, I.; Hegemann, P. Targeting of Photoreceptor Genes in Chlamydomonas reinhardtii via Zinc-Finger Nucleases and CRISPR/Cas9. Plant Cell 2017, 29, 2498–2518. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CrTRXf2 | CrTRXh1 | |
|---|---|---|
| Wavelength (Å) | 0.9762 | 0.9677 |
| Resolution range | 42.85–2.01 (2.082–2.01) | 36.42–1.378 (1.427–1.378) |
| Space group | I 2 2 2 | P 31 2 1 |
| Unit cell | 65.383 97.475 139.545 90 90 90 | 48.76 48.76 143.97 90 90 120 |
| Total reflections | 57,413 (5796) | 83,812 (8115) |
| Unique reflections | 29,702 (2973) | 41,955 (4085) |
| Multiplicity | 1.9 (2.0) | 2.0 (2.0) |
| Completeness (%) | 98.74 (99.80) | 99.80 (98.81) |
| Mean I/sigma (I) | 6.69 (0.95) | 25.45 (3.98) |
| Wilson B-factor | 34.89 | 16.29 |
| R-merge | 0.0697 (0.7309) | 0.01107 (0.1238) |
| R-meas | 0.09857 (1.034) | 0.01565 (0.1751) |
| R-pim | 0.0697 (0.7309) | 0.01107 (0.1238) |
| CC1/2 | 0.997 (0.485) | 1 (0.968) |
| CC * | 0.999 (0.808) | 1 (0.992) |
| Reflections used in refinement | 29,749 (2970) | 41,938 (4085) |
| Reflections used for R-free | 1998 (200) | 2095 (204) |
| R-work | 0.2262 (0.3131) | 0.1812 (0.2281) |
| R-free | 0.2638 (0.3606) | 0.2168 (0.2690) |
| CC (work) | 0.951 (0.709) | 0.964 (0.914) |
| CC (free) | 0.915 (0.649) | 0.946 (0.940) |
| Number of non-hydrogen atoms | 2764 | 2033 |
| Macromolecules | 2585 | 1638 |
| Solvent | 179 | 395 |
| Protein residues | 318 | 222 |
| RMS (bonds) | 0.008 | 0.005 |
| RMS (angles) | 0.97 | 0.77 |
| Ramachandran favored (%) | 98.34 | 98.17 |
| Ramachandran allowed (%) | 1.33 | 1.83 |
| Ramachandran outliers (%) | 0.33 | 0.00 |
| Rotamer outliers (%) | 0.34 | 0.60 |
| Clashscore | 5.18 | 4.24 |
| Average B-factor | 39.23 | 19.69 |
| Macromolecules | 39.04 | 17.65 |
| Solvent | 42.01 | 28.15 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lemaire, S.D.; Tedesco, D.; Crozet, P.; Michelet, L.; Fermani, S.; Zaffagnini, M.; Henri, J. Crystal Structure of Chloroplastic Thioredoxin f2 from Chlamydomonas reinhardtii Reveals Distinct Surface Properties. Antioxidants 2018, 7, 171. https://doi.org/10.3390/antiox7120171
Lemaire SD, Tedesco D, Crozet P, Michelet L, Fermani S, Zaffagnini M, Henri J. Crystal Structure of Chloroplastic Thioredoxin f2 from Chlamydomonas reinhardtii Reveals Distinct Surface Properties. Antioxidants. 2018; 7(12):171. https://doi.org/10.3390/antiox7120171
Chicago/Turabian StyleLemaire, Stéphane D., Daniele Tedesco, Pierre Crozet, Laure Michelet, Simona Fermani, Mirko Zaffagnini, and Julien Henri. 2018. "Crystal Structure of Chloroplastic Thioredoxin f2 from Chlamydomonas reinhardtii Reveals Distinct Surface Properties" Antioxidants 7, no. 12: 171. https://doi.org/10.3390/antiox7120171
APA StyleLemaire, S. D., Tedesco, D., Crozet, P., Michelet, L., Fermani, S., Zaffagnini, M., & Henri, J. (2018). Crystal Structure of Chloroplastic Thioredoxin f2 from Chlamydomonas reinhardtii Reveals Distinct Surface Properties. Antioxidants, 7(12), 171. https://doi.org/10.3390/antiox7120171