The Oxidized Protein Repair Enzymes Methionine Sulfoxide Reductases and Their Roles in Protecting against Oxidative Stress, in Ageing and in Regulating Protein Function

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methionine Sulfoxide Reductases Discovery
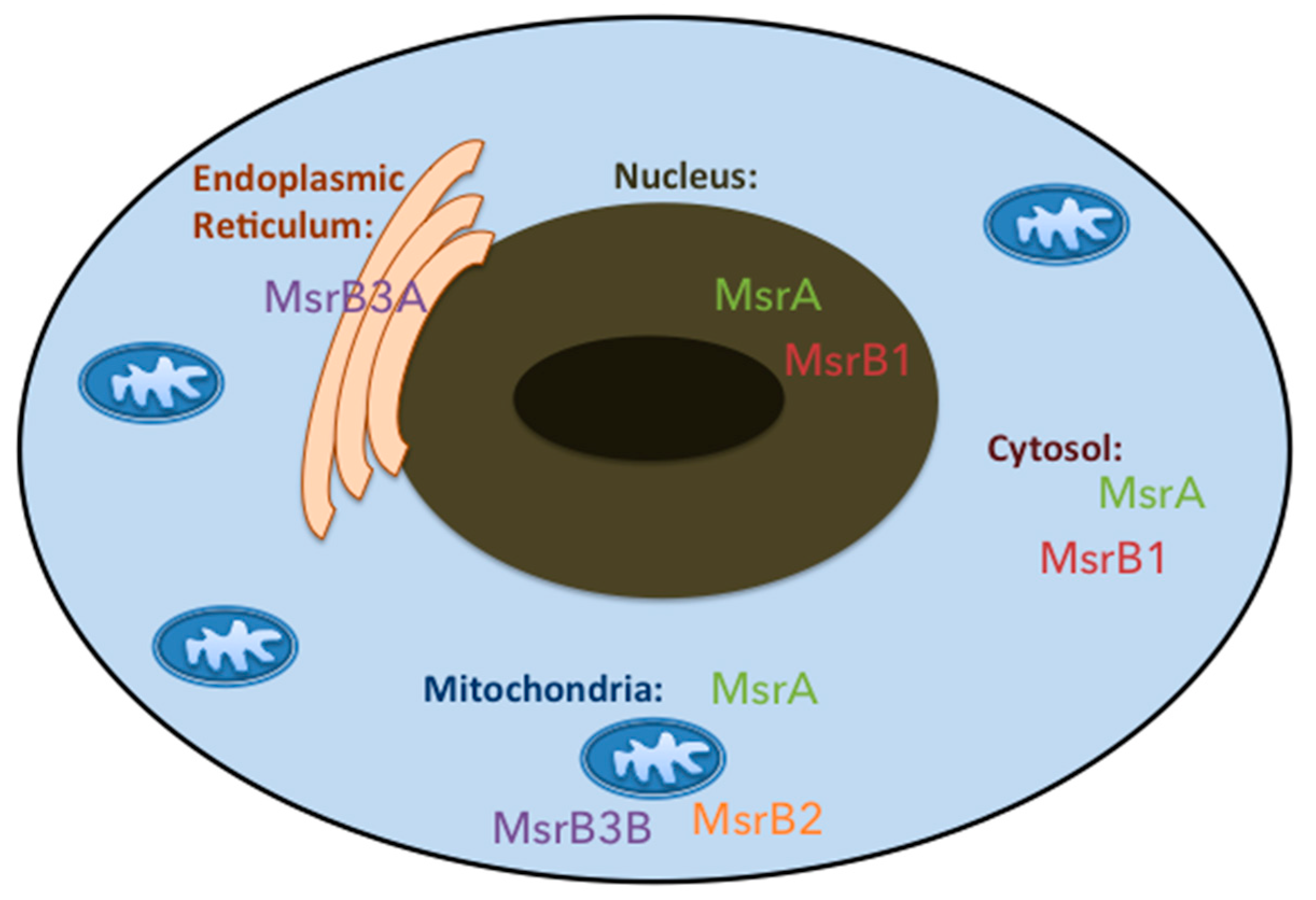
3. Methionine Sulfoxide Reductases Phylogenetic, Tissue and Cellular Distribution
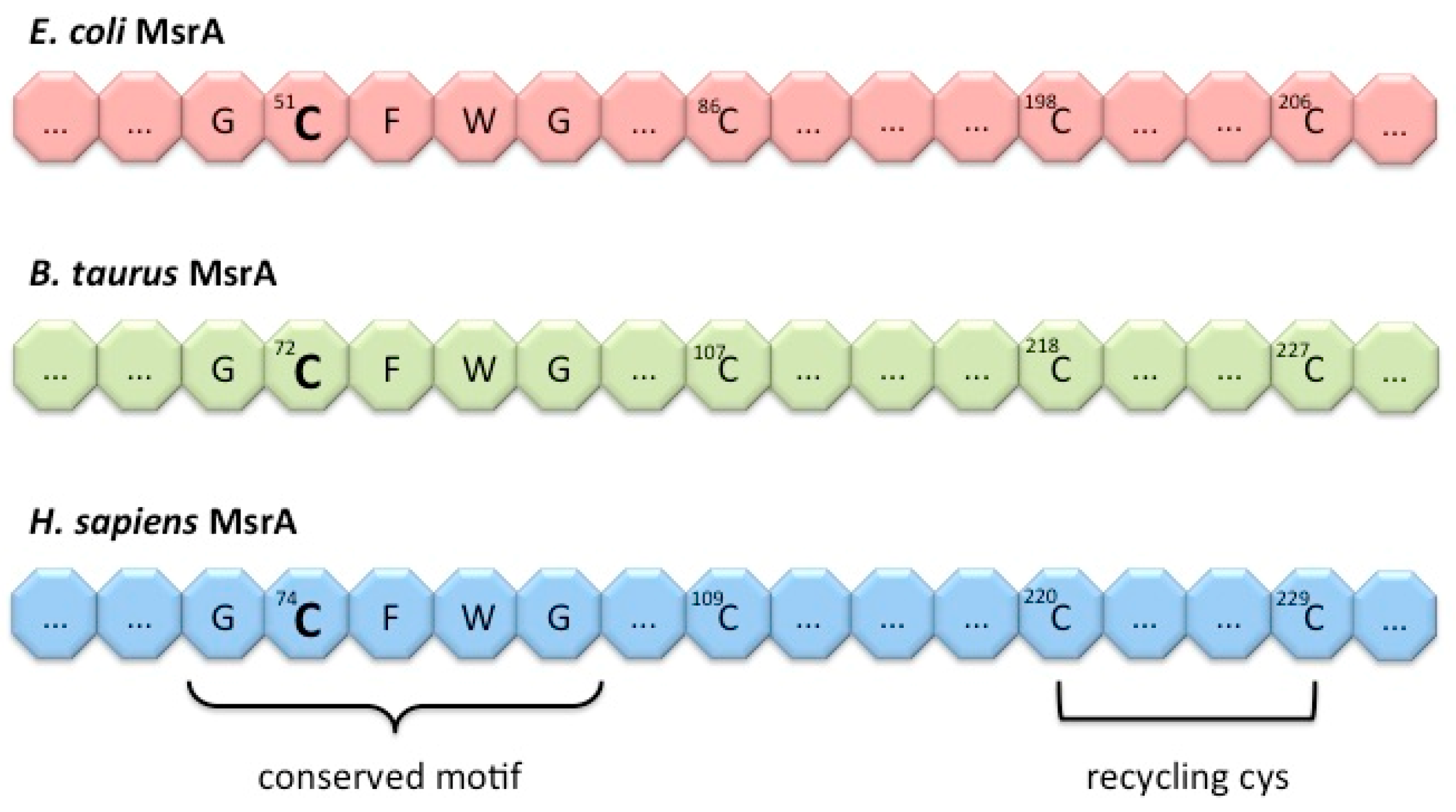
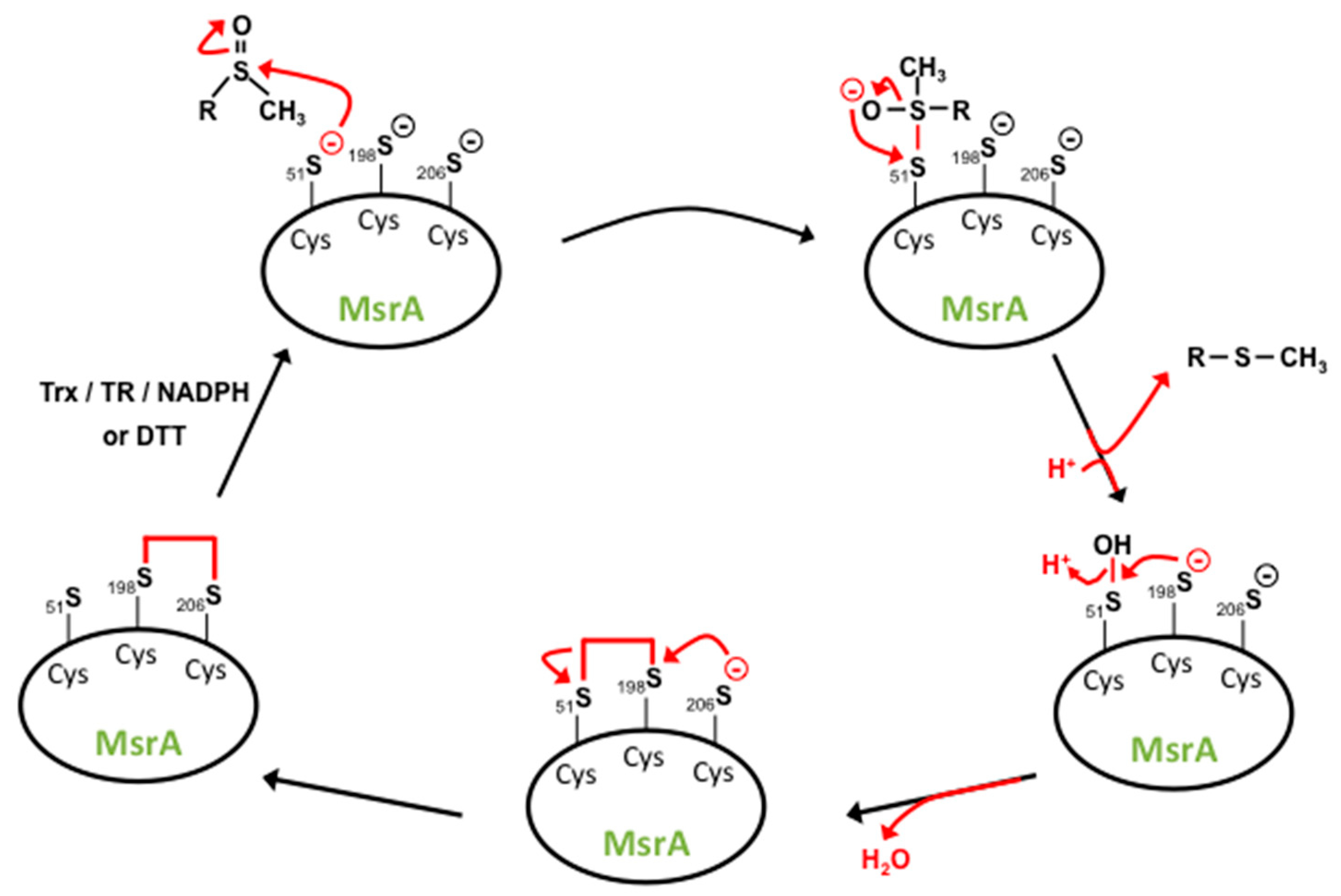
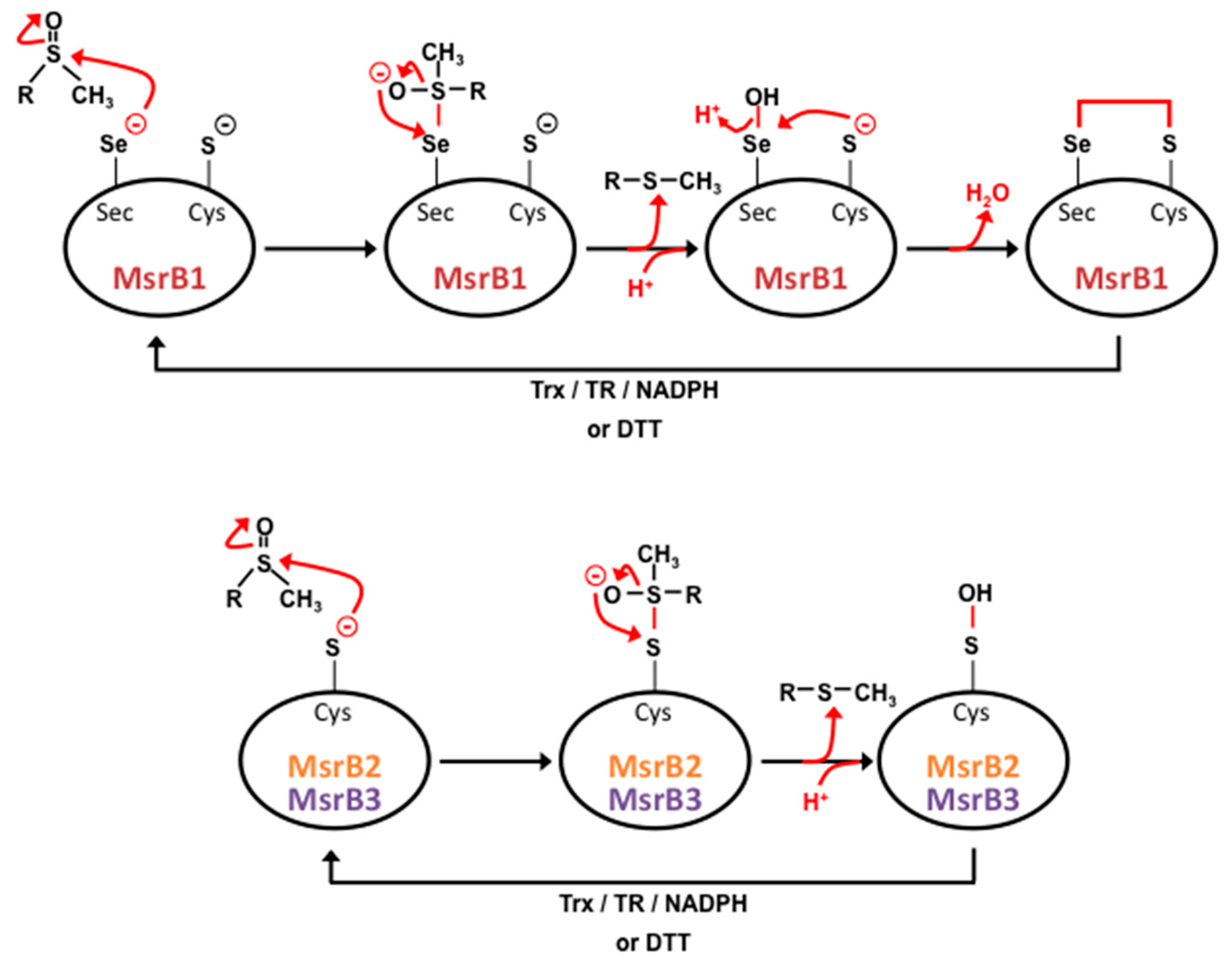
4. Methionine Sulfoxide Reductases Sequence, Structure and Catalytic Activity
5. Methionine Sulfoxide Reductases in Protection Against Oxidative Stress
6. Methionine Sulfoxide Reductases in Disease, Ageing and Longevity
7. Methionine Sulfoxide Reductases as Regulators of Protein and Cellular Functions
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lu, J.; Holmgren, A. The thioredoxin antioxidant system. Free Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Lillig, C.H.; Berndt, C.; Holmgren, A. Glutaredoxin systems. Biochim. Biophys. Acta 2008, 1780, 1304–1317. [Google Scholar] [CrossRef] [PubMed]
- Kalinina, E.V.; Chernov, N.N.; Novichkova, M.D. Role of glutathione, glutathione transferase, and glutaredoxin in regulation of redox-dependent processes. Biochemistry 2014, 79, 1562–1583. [Google Scholar] [CrossRef] [PubMed]
- Johansson, C.; Lillig, C.H.; Holmgren, A. Human mitochondrial glutaredoxin reduces S-glutathionylated proteins with high affinity accepting electrons from either glutathione or thioredoxin reductase. J. Biol. Chem. 2004, 279, 7537–7543. [Google Scholar] [CrossRef] [PubMed]
- Song, J.J.; Rhee, J.G.; Suntharalingam, M.; Walsh, S.A.; Spitz, D.R.; Lee, Y.J. Role of glutaredoxin in metabolic oxidative stress. Glutaredoxin as a sensor of oxidative stress mediated by H2O2. J. Biol. Chem. 2002, 277, 46566–46575. [Google Scholar] [CrossRef] [PubMed]
- Starke, D.W.; Chock, P.B.; Mieyal, J.J. Glutathione-thiyl radical scavenging and transferase properties of human glutaredoxin (thioltransferase). Potential role in redox signal transduction. J. Biol. Chem. 2003, 278, 14607–14613. [Google Scholar] [CrossRef]
- Lee, B.C.; Dikiy, A.; Kim, H.Y.; Gladyshev, V.N. Functions and evolution of selenoprotein methionine sulfoxide reductases. Biochim. Biophys. Acta 2009, 1790, 1471–1477. [Google Scholar] [CrossRef] [Green Version]
- Cabreiro, F.; Picot, C.R.; Perichon, M.; Castel, J.; Friguet, B.; Petropoulos, I. Overexpression of mitochondrial methionine sulfoxide reductase B2 protects leukemia cells from oxidative stress-induced cell death and protein damage. J. Biol. Chem. 2008, 283, 16673–16681. [Google Scholar] [CrossRef]
- Salmon, A.B.; Perez, V.I.; Bokov, A.; Jernigan, A.; Kim, G.; Zhao, H.; Levine, R.L.; Richardson, A. Lack of methionine sulfoxide reductase A in mice increases sensitivity to oxidative stress but does not diminish life span. FASEB J. 2009, 23, 3601–3608. [Google Scholar] [CrossRef] [Green Version]
- Ugarte, N.; Petropoulos, I.; Friguet, B. Oxidized mitochondrial protein degradation and repair in aging and oxidative stress. Antioxid. Redox Signal. 2010, 13, 539–549. [Google Scholar] [CrossRef]
- Chung, H.; Kim, A.K.; Jung, S.A.; Kim, S.W.; Yu, K.; Lee, J.H. The Drosophila homolog of methionine sulfoxide reductase A extends lifespan and increases nuclear localization of FOXO. FEBS Lett. 2010, 584, 3609–3614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minniti, A.N.; Cataldo, R.; Trigo, C.; Vasquez, L.; Mujica, P.; Leighton, F.; Inestrosa, N.C.; Aldunate, R. Methionine sulfoxide reductase A expression is regulated by the DAF-16/FOXO pathway in Caenorhabditis elegans. Aging Cell 2009, 8, 690–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koc, A.; Gasch, A.P.; Rutherford, J.C.; Kim, H.Y.; Gladyshev, V.N. Methionine sulfoxide reductase regulation of yeast lifespan reveals reactive oxygen species-dependent and -independent components of aging. Proc. Natl. Acad. Sci. USA 2004, 101, 7999–8004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaya, A.; Koc, A.; Lee, B.C.; Fomenko, D.E.; Rederstorff, M.; Krol, A.; Lescure, A.; Gladyshev, V.N. Compartmentalization and regulation of mitochondrial function by methionine sulfoxide reductases in yeast. Biochemistry 2010, 49, 8618–8625. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.C.; Peterfi, Z.; Hoffmann, F.W.; Moore, R.E.; Kaya, A.; Avanesov, A.; Tarrago, L.; Zhou, Y.; Weerapana, E.; Fomenko, D.E.; et al. MsrB1 and MICALs regulate actin assembly and macrophage function via reversible stereoselective methionine oxidation. Mol. Cell 2013, 51, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Bigelow, D.J.; Squier, T.C. Thioredoxin-dependent redox regulation of cellular signaling and stress response through reversible oxidation of methionines. Mol. Biosyst. 2011, 7, 2101–2109. [Google Scholar] [CrossRef] [PubMed]
- Waelsch, H.; Owades, P.; Miller, H.K.; Borek, E. Glutamic acid antimetabolites; the sulfoxide derived from methionine. J. Biol. Chem. 1946, 166, 273–281. [Google Scholar]
- Krask, B.J. Methionine sulfoxide and specific inhibition of sporulation in Bacillus subtilis. J. Bacteriol. 1953, 66, 374. [Google Scholar]
- Black, S.D.; Harte, E.M.; Hudson, B.; Wartofsky, L. A specific enzymatic reduction of L(-) methionine sulfoxide and a related non-specific reduction of disulfides. J. Biol. Chem. 1960, 235, 2910–2916. [Google Scholar]
- Ejiri, S.I.; Weissbach, H.; Brot, N. Reduction of methionine sulfoxide to methionine by Escherichia coli. J. Bacteriol. 1979, 139, 161–164. [Google Scholar]
- Doney, R.C.; Thompson, J.F. The reduction of S-methyl-L-cysteine sulfoxide and L-methionine sulfoxide in turnip and bean leaves. Biochim. Biophys. Acta 1966, 124, 39–49. [Google Scholar] [CrossRef]
- Aymarda, C.; Seyera, L.; Cheftela, J.-C. Enzymatic Reduction of Methionine Sulfoxide. In Vitro Experiments with Rat Liver and Kidney. Biol. Chem. 1979, 43, 1869–1876. [Google Scholar] [CrossRef]
- Brot, N.; Weissbach, L.; Werth, J.; Weissbach, H. Enzymatic reduction of protein-bound methionine sulfoxide. Proc. Natl. Acad. Sci. USA 1981, 78, 2155–2158. [Google Scholar] [CrossRef] [PubMed]
- Brot, N.; Werth, J.; Koster, D.; Weissbach, H. Reduction of N-acetyl methionine sulfoxide: A simple assay for peptide methionine sulfoxide reductase. Anal. Biochem. 1982, 122, 291–294. [Google Scholar] [CrossRef]
- Moskovitz, J.; Jenkins, N.A.; Gilbert, D.J.; Copeland, N.G.; Jursky, F.; Weissbach, H.; Brot, N. Chromosomal localization of the mammalian peptide-methionine sulfoxide reductase gene and its differential expression in various tissues. Proc. Natl. Acad. Sci. USA 1996, 93, 3205–3208. [Google Scholar] [CrossRef] [PubMed]
- Sharov, V.S.; Ferrington, D.A.; Squier, T.C.; Schoneich, C. Diastereoselective reduction of protein-bound methionine sulfoxide by methionine sulfoxide reductase. FEBS Lett. 1999, 455, 247–250. [Google Scholar] [CrossRef] [Green Version]
- Sharov, V.S.; Schoneich, C. Diastereoselective protein methionine oxidation by reactive oxygen species and diastereoselective repair by methionine sulfoxide reductase. Free Radic. Biol. Med. 2000, 29, 986–994. [Google Scholar] [CrossRef]
- Grimaud, R.; Ezraty, B.; Mitchell, J.K.; Lafitte, D.; Briand, C.; Derrick, P.J.; Barras, F. Repair of oxidized proteins. Identification of a new methionine sulfoxide reductase. J. Biol. Chem. 2001, 276, 48915–48920. [Google Scholar] [CrossRef]
- Kryukov, G.V.; Kumar, R.A.; Koc, A.; Sun, Z.; Gladyshev, V.N. Selenoprotein R is a zinc-containing stereo-specific methionine sulfoxide reductase. Proc. Natl. Acad. Sci. USA 2002, 99, 4245–4250. [Google Scholar] [CrossRef] [Green Version]
- Olry, A.; Boschi-Muller, S.; Marraud, M.; Sanglier-Cianferani, S.; Van Dorsselear, A.; Branlant, G. Characterization of the methionine sulfoxide reductase activities of PILB, a probable virulence factor from Neisseria meningitidis. J. Biol. Chem. 2002, 277, 12016–12022. [Google Scholar] [CrossRef]
- Lescure, A.; Gautheret, D.; Carbon, P.; Krol, A. Novel selenoproteins identified in silico and in vivo by using a conserved RNA structural motif. J. Biol. Chem. 1999, 274, 38147–38154. [Google Scholar] [CrossRef] [PubMed]
- Etienne, F.; Spector, D.; Brot, N.; Weissbach, H. A methionine sulfoxide reductase in Escherichia coli that reduces the R enantiomer of methionine sulfoxide. Biochem. Biophys. Res. Commun. 2003, 300, 378–382. [Google Scholar] [CrossRef]
- Lin, Z.; Johnson, L.C.; Weissbach, H.; Brot, N.; Lively, M.O.; Lowther, W.T. Free methionine-(R)-sulfoxide reductase from Escherichia coli reveals a new GAF domain function. Proc. Natl. Acad. Sci. USA 2007, 104, 9597–9602. [Google Scholar] [CrossRef] [PubMed]
- Gennaris, A.; Ezraty, B.; Henry, C.; Agrebi, R.; Vergnes, A.; Oheix, E.; Bos, J.; Leverrier, P.; Espinosa, L.; Szewczyk, J.; et al. Repairing oxidized proteins in the bacterial envelope using respiratory chain electrons. Nature 2015, 528, 409–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.H.; Weissbach, H. Origin and evolution of the protein-repairing enzymes methionine sulphoxide reductases. Biol. Rev. 2008, 83, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, E.; Shinka, Y.; Fukui, T.; Atomi, H.; Imanaka, T. Methionine sulfoxide reductase from the hyperthermophilic archaeon Thermococcus kodakaraensis, an enzyme designed to function at suboptimal growth temperatures. J. Bacteriol. 2007, 189, 7134–7144. [Google Scholar] [CrossRef]
- Kim, H.S.; Kwak, G.H.; Lee, K.; Jo, C.H.; Hwang, K.Y.; Kim, H.Y. Structural and biochemical analysis of a type II free methionine-R-sulfoxide reductase from Thermoplasma acidophilum. Arch. Biochem. Biophys. 2014, 560, 10–19. [Google Scholar] [CrossRef]
- Rouhier, N.; Vieira Dos Santos, C.; Tarrago, L.; Rey, P. Plant methionine sulfoxide reductase A and B multigenic families. Photosynth. Res. 2006, 89, 247–262. [Google Scholar] [CrossRef]
- Kim, H.Y.; Gladyshev, V.N. Methionine sulfoxide reduction in mammals: Characterization of methionine-R-sulfoxide reductases. Mol. Biol. Cell 2004, 15, 1055–1064. [Google Scholar] [CrossRef]
- Kwon, S.J.; Kwon, S.I.; Bae, M.S.; Cho, E.J.; Park, O.K. Role of the methionine sulfoxide reductase MsrB3 in cold acclimation in Arabidopsis. Plant Cell Physiol. 2007, 48, 1713–1723. [Google Scholar] [CrossRef]
- Lim, D.H.; Han, J.Y.; Kim, J.R.; Lee, Y.S.; Kim, H.Y. Methionine sulfoxide reductase B in the endoplasmic reticulum is critical for stress resistance and aging in Drosophila. Biochem. Biophys. Res. Commun. 2012, 419, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Kuschel, L.; Hansel, A.; Schonherr, R.; Weissbach, H.; Brot, N.; Hoshi, T.; Heinemann, S.H. Molecular cloning and functional expression of a human peptide methionine sulfoxide reductase (hMsrA). FEBS Lett. 1999, 456, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Moskovitz, J.; Weissbach, H.; Brot, N. Cloning the expression of a mammalian gene involved in the reduction of methionine sulfoxide residues in proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 2095–2099. [Google Scholar] [CrossRef] [PubMed]
- Brot, N.; Fliss, H.; Coleman, T.; Weissbach, H. Enzymatic reduction of methionine sulfoxide residues in proteins and peptides. Methods Enzymol. 1984, 107, 352–360. [Google Scholar] [PubMed]
- Achilli, C.; Ciana, A.; Rossi, A.; Balduini, C.; Minetti, G. Neutrophil granulocytes uniquely express, among human blood cells, high levels of Methionine-sulfoxide-reductase enzymes. J. Leukoc. Biol. 2008, 83, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, F.; Sander, C.S.; Hansel, A.; Oehrl, W.; Kasperczyk, H.; Elsner, P.; Shimizu, K.; Heinemann, S.H.; Thiele, J.J. The repair enzyme peptide methionine-S-sulfoxide reductase is expressed in human epidermis and upregulated by UVA radiation. J. Investig. Dermatol. 2006, 126, 1128–1134. [Google Scholar] [CrossRef] [PubMed]
- Picot, C.R.; Moreau, M.; Juan, M.; Noblesse, E.; Nizard, C.; Petropoulos, I.; Friguet, B. Impairment of methionine sulfoxide reductase during UV irradiation and photoaging. Exp. Gerontol. 2007, 42, 859–863. [Google Scholar] [CrossRef]
- Schallreuter, K.U.; Rubsam, K.; Chavan, B.; Zothner, C.; Gillbro, J.M.; Spencer, J.D.; Wood, J.M. Functioning methionine sulfoxide reductases A and B are present in human epidermal melanocytes in the cytosol and in the nucleus. Biochem. Biophys. Res. Commun. 2006, 342, 145–152. [Google Scholar] [CrossRef]
- Taungjaruwinai, W.M.; Bhawan, J.; Keady, M.; Thiele, J.J. Differential expression of the antioxidant repair enzyme methionine sulfoxide reductase (MSRA and MSRB) in human skin. Am. J. Dermatopathol. 2009, 31, 427–431. [Google Scholar] [CrossRef]
- Hansel, A.; Kuschel, L.; Hehl, S.; Lemke, C.; Agricola, H.J.; Hoshi, T.; Heinemann, S.H. Mitochondrial targeting of the human peptide methionine sulfoxide reductase (MSRA), an enzyme involved in the repair of oxidized proteins. FASEB J. 2002, 16, 911–913. [Google Scholar] [CrossRef]
- Vougier, S.; Mary, J.; Friguet, B. Subcellular localization of methionine sulphoxide reductase A (MsrA): Evidence for mitochondrial and cytosolic isoforms in rat liver cells. Biochem. J. 2003, 373, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Gladyshev, V.N. Role of structural and functional elements of mouse methionine-S-sulfoxide reductase in its subcellular distribution. Biochemistry 2005, 44, 8059–8067. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Gladyshev, V.N. Alternative first exon splicing regulates subcellular distribution of methionine sulfoxide reductases. BMC Mol. Biol. 2006, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.M.; Lim, J.C.; Kim, G.; Levine, R.L. Myristoylated methionine sulfoxide reductase A is a late endosomal protein. J. Biol. Chem. 2018, 293, 7355–7366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.Y.; Gladyshev, V.N. Characterization of mouse endoplasmic reticulum methionine-R-sulfoxide reductase. Biochem. Biophys. Res. Commun. 2004, 320, 1277–1283. [Google Scholar] [CrossRef]
- Lowther, W.T.; Brot, N.; Weissbach, H.; Honek, J.F.; Matthews, B.W. Thiol-disulfide exchange is involved in the catalytic mechanism of peptide methionine sulfoxide reductase. Proc. Natl. Acad. Sci. USA 2000, 97, 6463–6468. [Google Scholar] [CrossRef] [Green Version]
- Tete-Favier, F.; Cobessi, D.; Leonard, G.A.; Azza, S.; Talfournier, F.; Boschi-Muller, S.; Branlant, G.; Aubry, A. Crystallization and preliminary X-ray diffraction studies of the peptide methionine sulfoxide reductase from Escherichia coli. Acta Crystallogr. D Biol. Crystallogr. 2000, 56, 1194–1197. [Google Scholar] [CrossRef]
- Lowther, W.T.; Brot, N.; Weissbach, H.; Matthews, B.W. Structure and mechanism of peptide methionine sulfoxide reductase, an “anti-oxidation” enzyme. Biochemistry 2000, 39, 13307–13312. [Google Scholar] [CrossRef]
- Taylor, A.B.; Benglis, D.M., Jr.; Dhandayuthapani, S.; Hart, P.J. Structure of Mycobacterium tuberculosis methionine sulfoxide reductase A in complex with protein-bound methionine. J. Bacteriol. 2003, 185, 4119–4126. [Google Scholar] [CrossRef]
- Rouhier, N.; Kauffmann, B.; Tete-Favier, F.; Palladino, P.; Gans, P.; Branlant, G.; Jacquot, J.P.; Boschi-Muller, S. Functional and structural aspects of poplar cytosolic and plastidial type a methionine sulfoxide reductases. J. Biol. Chem. 2007, 282, 3367–3378. [Google Scholar] [CrossRef]
- Ranaivoson, F.M.; Antoine, M.; Kauffmann, B.; Boschi-Muller, S.; Aubry, A.; Branlant, G.; Favier, F. A structural analysis of the catalytic mechanism of methionine sulfoxide reductase A from Neisseria meningitidis. J. Mol. Biol. 2008, 377, 268–280. [Google Scholar] [CrossRef]
- Coudevylle, N.; Antoine, M.; Bouguet-Bonnet, S.; Mutzenhardt, P.; Boschi-Muller, S.; Branlant, G.; Cung, M.T. Solution structure and backbone dynamics of the reduced form and an oxidized form of E. coli methionine sulfoxide reductase A (MsrA): Structural insight of the MsrA catalytic cycle. J. Mol. Biol. 2007, 366, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Boschi-Muller, S.; Azza, S.; Sanglier-Cianferani, S.; Talfournier, F.; Van Dorsselear, A.; Branlant, G. A sulfenic acid enzyme intermediate is involved in the catalytic mechanism of peptide methionine sulfoxide reductase from Escherichia coli. J. Biol. Chem. 2000, 275, 35908–35913. [Google Scholar] [CrossRef] [PubMed]
- Olry, A.; Boschi-Muller, S.; Branlant, G. Kinetic characterization of the catalytic mechanism of methionine sulfoxide reductase B from Neisseria meningitidis. Biochemistry 2004, 43, 11616–11622. [Google Scholar] [CrossRef] [PubMed]
- Antoine, M.; Boschi-Muller, S.; Branlant, G. Kinetic characterization of the chemical steps involved in the catalytic mechanism of methionine sulfoxide reductase A from Neisseria meningitidis. J. Biol. Chem. 2003, 278, 45352–45357. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.C.; You, Z.; Kim, G.; Levine, R.L. Methionine sulfoxide reductase A is a stereospecific methionine oxidase. Proc. Natl. Acad. Sci. USA 2011, 108, 10472–10477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriznik, A.; Boschi-Muller, S.; Branlant, G. Kinetic evidence that methionine sulfoxide reductase A can reveal its oxidase activity in the presence of thioredoxin. Arch. Biochem. Biophys. 2014, 548, 54–59. [Google Scholar] [CrossRef]
- Lowther, W.T.; Weissbach, H.; Etienne, F.; Brot, N.; Matthews, B.W. The mirrored methionine sulfoxide reductases of Neisseria gonorrhoeae pilB. Nat. Struct. Biol. 2002, 9, 348–352. [Google Scholar] [CrossRef]
- Wu, J.; Neiers, F.; Boschi-Muller, S.; Branlant, G. The N-terminal domain of PILB from Neisseria meningitidis is a disulfide reductase that can recycle methionine sulfoxide reductases. J. Biol. Chem. 2005, 280, 12344–12350. [Google Scholar] [CrossRef]
- Kim, H.Y.; Gladyshev, V.N. Different catalytic mechanisms in mammalian selenocysteine- and cysteine-containing methionine-R-sulfoxide reductases. PLoS Biol. 2005, 3, E375. [Google Scholar] [CrossRef]
- Jung, S.; Hansel, A.; Kasperczyk, H.; Hoshi, T.; Heinemann, S.H. Activity, tissue distribution and site-directed mutagenesis of a human peptide methionine sulfoxide reductase of type B: HCBS1. FEBS Lett. 2002, 527, 91–94. [Google Scholar] [CrossRef]
- Sagher, D.; Brunell, D.; Hejtmancik, J.F.; Kantorow, M.; Brot, N.; Weissbach, H. Thionein can serve as a reducing agent for the methionine sulfoxide reductases. Proc. Natl. Acad. Sci. USA 2006, 103, 8656–8661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagher, D.; Brunell, D.; Brot, N.; Vallee, B.L.; Weissbach, H. Selenocompounds can serve as oxidoreductants with the methionine sulfoxide reductase enzymes. J. Biol. Chem. 2006, 281, 31184–31187. [Google Scholar] [CrossRef] [PubMed]
- Couturier, J.; Vignols, F.; Jacquot, J.P.; Rouhier, N. Glutathione- and glutaredoxin-dependent reduction of methionine sulfoxide reductase A. FEBS Lett. 2012, 586, 3894–3899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.J.; Lee, B.C.; Jeong, J.; Lee, K.J.; Hwang, K.Y.; Gladyshev, V.N.; Kim, H.Y. Tandem use of selenocysteine: Adaptation of a selenoprotein glutaredoxin for reduction of selenoprotein methionine sulfoxide reductase. Mol. Microbiol. 2011, 79, 1194–1203. [Google Scholar] [CrossRef] [PubMed]
- Tarrago, L.; Laugier, E.; Zaffagnini, M.; Marchand, C.; Le Marechal, P.; Rouhier, N.; Lemaire, S.D.; Rey, P. Regeneration mechanisms of Arabidopsis thaliana methionine sulfoxide reductases B by glutaredoxins and thioredoxins. J. Biol. Chem. 2009, 284, 18963–18971. [Google Scholar] [CrossRef] [PubMed]
- Vieira Dos Santos, C.; Laugier, E.; Tarrago, L.; Massot, V.; Issakidis-Bourguet, E.; Rouhier, N.; Rey, P. Specificity of thioredoxins and glutaredoxins as electron donors to two distinct classes of Arabidopsis plastidial methionine sulfoxide reductases B. FEBS Lett. 2007, 581, 4371–4376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, C.; Kriznik, A.; Boschi-Muller, S.; Branlant, G. Thioredoxin 2 from Escherichia coli is not involved in vivo in the recycling process of methionine sulfoxide reductase activities. FEBS Lett. 2011, 585, 1905–1909. [Google Scholar] [CrossRef]
- Levine, R.L.; Mosoni, L.; Berlett, B.S.; Stadtman, E.R. Methionine residues as endogenous antioxidants in proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 15036–15040. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.R.; Narhi, L.O.; Spahr, C.; Langley, K.E.; Lu, H.S. In vitro methionine oxidation of Escherichia coli-derived human stem cell factor: Effects on the molecular structure, biological activity, and dimerization. Protein Sci. 1996, 5, 1165–1173. [Google Scholar] [CrossRef]
- Jones, T.E.; Alexander, R.W.; Pan, T. Misacylation of specific nonmethionyl tRNAs by a bacterial methionyl-tRNA synthetase. Proc. Natl. Acad. Sci. USA 2011, 108, 6933–6938. [Google Scholar] [CrossRef] [PubMed]
- Wiltrout, E.; Goodenbour, J.M.; Frechin, M.; Pan, T. Misacylation of tRNA with methionine in Saccharomyces cerevisiae. Nucleic Acids Res. 2012, 40, 10494–10506. [Google Scholar] [CrossRef] [PubMed]
- Netzer, N.; Goodenbour, J.M.; David, A.; Dittmar, K.A.; Jones, R.B.; Schneider, J.R.; Boone, D.; Eves, E.M.; Rosner, M.R.; Gibbs, J.S.; et al. Innate immune and chemically triggered oxidative stress modifies translational fidelity. Nature 2009, 462, 522–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Pan, T. Methionine Mistranslation Bypasses the Restraint of the Genetic Code to Generate Mutant Proteins with Distinct Activities. PLOS Genet. 2015, 11, e1005745. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Levine, R.L. Methionine in proteins defends against oxidative stress. FASEB J. 2009, 23, 464–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St John, G.; Brot, N.; Ruan, J.; Erdjument-Bromage, H.; Tempst, P.; Weissbach, H.; Nathan, C. Peptide methionine sulfoxide reductase from Escherichia coli and Mycobacterium tuberculosis protects bacteria against oxidative damage from reactive nitrogen intermediates. Proc. Natl. Acad. Sci. USA 2001, 98, 9901–9906. [Google Scholar] [CrossRef] [PubMed]
- Moskovitz, J.; Rahman, M.A.; Strassman, J.; Yancey, S.O.; Kushner, S.R.; Brot, N.; Weissbach, H. Escherichia coli peptide methionine sulfoxide reductase gene: Regulation of expression and role in protecting against oxidative damage. J. Bacteriol. 1995, 177, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Tamburro, A.; Robuffo, I.; Heipieper, H.J.; Allocati, N.; Rotilio, D.; Di Ilio, C.; Favaloro, B. Expression of glutathione S-transferase and peptide methionine sulphoxide reductase in Ochrobactrum anthropi is correlated to the production of reactive oxygen species caused by aromatic substrates. FEMS Microbiol. Lett. 2004, 241, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Moskovitz, J.; Wilkinson, B.J.; Jayaswal, R.K. Molecular characterization of a chromosomal locus in Staphylococcus aureus that contributes to oxidative defence and is highly induced by the cell-wall-active antibiotic oxacillin. Microbiology 2001, 147, 3037–3045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, T.; Daniel, D.S.; Parida, B.K.; Jagannath, C.; Dhandayuthapani, S. Methionine sulfoxide reductase A (MsrA) deficiency affects the survival of Mycobacterium smegmatis within macrophages. J. Bacteriol. 2004, 186, 3590–3598. [Google Scholar] [CrossRef] [PubMed]
- Oien, D.; Moskovitz, J. Protein-carbonyl accumulation in the non-replicative senescence of the methionine sulfoxide reductase A (msrA) knockout yeast strain. Amino Acids 2007, 32, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Moskovitz, J.; Berlett, B.S.; Poston, J.M.; Stadtman, E.R. The yeast peptide-methionine sulfoxide reductase functions as an antioxidant in vivo. Proc. Natl. Acad. Sci. USA 1997, 94, 9585–9589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moskovitz, J.; Flescher, E.; Berlett, B.S.; Azare, J.; Poston, J.M.; Stadtman, E.R. Overexpression of peptide-methionine sulfoxide reductase in Saccharomyces cerevisiae and human T cells provides them with high resistance to oxidative stress. Proc. Natl. Acad. Sci. USA 1998, 95, 14071–14075. [Google Scholar] [CrossRef] [PubMed]
- Sumner, E.R.; Shanmuganathan, A.; Sideri, T.C.; Willetts, S.A.; Houghton, J.E.; Avery, S.V. Oxidative protein damage causes chromium toxicity in yeast. Microbiology 2005, 151, 1939–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechtold, U.; Murphy, D.J.; Mullineaux, P.M. Arabidopsis peptide methionine sulfoxide reductase2 prevents cellular oxidative damage in long nights. Plant Cell 2004, 16, 908–919. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.S.; Cho, E.J.; Choi, E.Y.; Park, O.K. Analysis of the Arabidopsis nuclear proteome and its response to cold stress. Plant J. 2003, 36, 652–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, H.; Tang, X.D.; Chen, M.L.; Joiner, M.L.; Sun, G.; Brot, N.; Weissbach, H.; Heinemann, S.H.; Iverson, L.; Wu, C.F.; et al. High-quality life extension by the enzyme peptide methionine sulfoxide reductase. Proc. Natl. Acad. Sci. USA 2002, 99, 2748–2753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roesijadi, G.; Rezvankhah, S.; Binninger, D.M.; Weissbach, H. Ecdysone induction of MsrA protects against oxidative stress in Drosophila. Biochem. Biophys. Res. Commun. 2007, 354, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Moskovitz, J.; Bar-Noy, S.; Williams, W.M.; Requena, J.; Berlett, B.S.; Stadtman, E.R. Methionine sulfoxide reductase (MsrA) is a regulator of antioxidant defense and lifespan in mammals. Proc. Natl. Acad. Sci. USA 2001, 98, 12920–12925. [Google Scholar] [CrossRef] [Green Version]
- Nan, C.; Li, Y.; Jean-Charles, P.Y.; Chen, G.; Kreymerman, A.; Prentice, H.; Weissbach, H.; Huang, X. Deficiency of methionine sulfoxide reductase A causes cellular dysfunction and mitochondrial damage in cardiac myocytes under physical and oxidative stresses. Biochem. Biophys. Res. Commun. 2010, 402, 608–613. [Google Scholar] [CrossRef]
- Picot, C.R.; Perichon, M.; Lundberg, K.C.; Friguet, B.; Szweda, L.I.; Petropoulos, I. Alterations in mitochondrial and cytosolic methionine sulfoxide reductase activity during cardiac ischemia and reperfusion. Exp. Gerontol. 2006, 41, 663–667. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Sun, J.; Deschamps, A.M.; Kim, G.; Liu, C.; Murphy, E.; Levine, R.L. Myristoylated methionine sulfoxide reductase A protects the heart from ischemia-reperfusion injury. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H1513–H1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.I.; Choi, S.H.; Jung, K.J.; Lee, E.; Kim, H.Y.; Park, K.M. Protective role of methionine sulfoxide reductase A against ischemia/reperfusion injury in mouse kidney and its involvement in the regulation of trans-sulfuration pathway. Antioxid. Redox Signal. 2013, 18, 2241–2250. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.I.; Noh, M.R.; Kim, K.Y.; Jang, H.S.; Kim, H.Y.; Park, K.M. Methionine sulfoxide reductase A deficiency exacerbates progression of kidney fibrosis induced by unilateral ureteral obstruction. Free Radic. Biol. Med. 2015, 89, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Fomenko, D.E.; Novoselov, S.V.; Natarajan, S.K.; Lee, B.C.; Koc, A.; Carlson, B.A.; Lee, T.H.; Kim, H.Y.; Hatfield, D.L.; Gladyshev, V.N. MsrB1 (methionine-R-sulfoxide reductase 1) knock-out mice: Roles of MsrB1 in redox regulation and identification of a novel selenoprotein form. J. Biol. Chem. 2009, 284, 5986–5993. [Google Scholar] [CrossRef] [PubMed]
- Moskovitz, J. Prolonged selenium-deficient diet in MsrA knockout mice causes enhanced oxidative modification to proteins and affects the levels of antioxidant enzymes in a tissue-specific manner. Free Radic. Res. 2007, 41, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Kwak, G.H.; Singh, M.P.; Gladyshev, V.N.; Kim, H.Y. Selenoprotein MsrB1 deficiency exacerbates acetaminophen-induced hepatotoxicity via increased oxidative damage. Arch. Biochem. Biophys. 2017, 634, 69–75. [Google Scholar] [CrossRef]
- Sreekumar, P.G.; Kannan, R.; Yaung, J.; Spee, C.K.; Ryan, S.J.; Hinton, D.R. Protection from oxidative stress by methionine sulfoxide reductases in RPE cells. Biochem. Biophys. Res. Commun. 2005, 334, 245–253. [Google Scholar] [CrossRef]
- Lee, J.W.; Gordiyenko, N.V.; Marchetti, M.; Tserentsoodol, N.; Sagher, D.; Alam, S.; Weissbach, H.; Kantorow, M.; Rodriguez, I.R. Gene structure, localization and role in oxidative stress of methionine sulfoxide reductase A (MSRA) in the monkey retina. Exp. Eye Res. 2006, 82, 816–827. [Google Scholar] [CrossRef]
- Dun, Y.; Vargas, J.; Brot, N.; Finnemann, S.C. Independent roles of methionine sulfoxide reductase A in mitochondrial ATP synthesis and as antioxidant in retinal pigment epithelial cells. Free Radic. Biol. Med. 2013, 65, 1340–1351. [Google Scholar] [CrossRef] [Green Version]
- Kantorow, M.; Hawse, J.R.; Cowell, T.L.; Benhamed, S.; Pizarro, G.O.; Reddy, V.N.; Hejtmancik, J.F. Methionine sulfoxide reductase A is important for lens cell viability and resistance to oxidative stress. Proc. Natl. Acad. Sci. USA 2004, 101, 9654–9659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchetti, M.A.; Pizarro, G.O.; Sagher, D.; Deamicis, C.; Brot, N.; Hejtmancik, J.F.; Weissbach, H.; Kantorow, M. Methionine sulfoxide reductases B1, B2, and B3 are present in the human lens and confer oxidative stress resistance to lens cells. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2107–2112. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Liu, H.; Zhou, J.; Huang, K. Selenoprotein R Protects Human Lens Epithelial Cells against d-Galactose-Induced Apoptosis by Regulating Oxidative Stress and Endoplasmic Reticulum Stress. Int. J. Mol. Sci. 2016, 17, 231. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Li, Y.; Du, S.; Huang, K. Involvement of MsrB1 in the regulation of redox balance and inhibition of peroxynitrite-induced apoptosis in human lens epithelial cells. Exp. Eye Res. 2012, 100, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jia, Y.; Zhou, J.; Huang, K. Effect of methionine sulfoxide reductase B1 silencing on high-glucose-induced apoptosis of human lens epithelial cells. Life Sci. 2013, 92, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Zhou, J.; Liu, H.; Huang, K. Effect of methionine sulfoxide reductase B1 (SelR) gene silencing on peroxynitrite-induced F-actin disruption in human lens epithelial cells. Biochem. Biophys. Res. Commun. 2014, 443, 876–881. [Google Scholar] [CrossRef] [PubMed]
- Ugarte, N.; Ladouce, R.; Radjei, S.; Gareil, M.; Friguet, B.; Petropoulos, I. Proteome alteration in oxidative stress-sensitive methionine sulfoxide reductase-silenced HEK293 cells. Free Radic. Biol. Med. 2013, 65, 1023–1036. [Google Scholar] [CrossRef] [PubMed]
- Cabreiro, F.; Picot, C.R.; Perichon, M.; Friguet, B.; Petropoulos, I. Overexpression of methionine sulfoxide reductases A and B2 protects MOLT-4 cells against zinc-induced oxidative stress. Antioxid. Redox Signal. 2009, 11, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Picot, C.R.; Petropoulos, I.; Perichon, M.; Moreau, M.; Nizard, C.; Friguet, B. Overexpression of MsrA protects WI-38 SV40 human fibroblasts against H2O2-mediated oxidative stress. Free Radic. Biol. Med. 2005, 39, 1332–1341. [Google Scholar] [CrossRef] [PubMed]
- Cabreiro, F.; Picot, C.R.; Perichon, M.; Mary, J.; Friguet, B.; Petropoulos, I. Identification of proteins undergoing expression level modifications in WI-38 SV40 fibroblasts overexpressing methionine sulfoxide reductase A. Biochimie 2007, 89, 1388–1395. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, Y.; Kwak, G.H.; Oh, S.Y.; Kim, H.Y. Over-expression of methionine sulfoxide reductase A in the endoplasmic reticulum increases resistance to oxidative and ER stresses. Acta Biochim. Biophys. Sin. (Shanghai) 2014, 46, 415–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, G.H.; Lim, D.H.; Han, J.Y.; Lee, Y.S.; Kim, H.Y. Methionine sulfoxide reductase B3 protects from endoplasmic reticulum stress in Drosophila and in mammalian cells. Biochem. Biophys. Res. Commun. 2012, 420, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Pelle, E.; Maes, D.; Huang, X.; Frenkel, K.; Pernodet, N.; Yarosh, D.B.; Zhang, Q. Protection against UVB-induced oxidative stress in human skin cells and skin models by methionine sulfoxide reductase A. J. Cosmet. Sci. 2012, 63, 359–364. [Google Scholar] [PubMed]
- Zhou, Z.; Li, C.Y.; Li, K.; Wang, T.; Zhang, B.; Gao, T.W. Decreased methionine sulphoxide reductase A expression renders melanocytes more sensitive to oxidative stress: A possible cause for melanocyte loss in vitiligo. Br. J. Dermatol. 2009, 161, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Gil-Mohapel, J.; Brocardo, P.S.; Christie, B.R. The role of oxidative stress in Huntington’s disease: Are antioxidants good therapeutic candidates? Curr. Drug Targets 2014, 15, 454–468. [Google Scholar] [CrossRef] [PubMed]
- Swomley, A.M.; Butterfield, D.A. Oxidative stress in Alzheimer disease and mild cognitive impairment: Evidence from human data provided by redox proteomics. Arch. Toxicol. 2015, 89, 1669–1680. [Google Scholar] [CrossRef] [PubMed]
- Gabbita, S.P.; Aksenov, M.Y.; Lovell, M.A.; Markesbery, W.R. Decrease in peptide methionine sulfoxide reductase in Alzheimer’s disease brain. J. Neurochem. 1999, 73, 1660–1666. [Google Scholar] [CrossRef]
- Brennan, L.A.; Lee, W.; Cowell, T.; Giblin, F.; Kantorow, M. Deletion of mouse MsrA results in HBO-induced cataract: MsrA repairs mitochondrial cytochrome c. Mol. Vis. 2009, 15, 985–999. [Google Scholar]
- Pal, R.; Oien, D.B.; Ersen, F.Y.; Moskovitz, J. Elevated levels of brain-pathologies associated with neurodegenerative diseases in the methionine sulfoxide reductase A knockout mouse. Exp. Brain Res. 2007, 180, 765–774. [Google Scholar] [CrossRef]
- Hou, L.; Kang, I.; Marchant, R.E.; Zagorski, M.G. Methionine 35 oxidation reduces fibril assembly of the amyloid abeta-(1-42) peptide of Alzheimer’s disease. J. Biol. Chem. 2002, 277, 40173–40176. [Google Scholar] [CrossRef]
- Moskovitz, J.; Du, F.; Bowman, C.F.; Yan, S.S. Methionine sulfoxide reductase A affects beta-amyloid solubility and mitochondrial function in a mouse model of Alzheimer’s disease. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E388–E393. [Google Scholar] [CrossRef] [PubMed]
- Moskovitz, J.; Maiti, P.; Lopes, D.H.; Oien, D.B.; Attar, A.; Liu, T.; Mittal, S.; Hayes, J.; Bitan, G. Induction of methionine-sulfoxide reductases protects neurons from amyloid beta-protein insults in vitro and in vivo. Biochemistry 2011, 50, 10687–10697. [Google Scholar] [CrossRef] [PubMed]
- Glaser, C.B.; Yamin, G.; Uversky, V.N.; Fink, A.L. Methionine oxidation, α-synuclein and Parkinson’s disease. Biochim. Biophys. Acta 2005, 1703, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Chondrogianni, N.; Petropoulos, I.; Grimm, S.; Georgila, K.; Catalgol, B.; Friguet, B.; Grune, T.; Gonos, E.S. Protein damage, repair and proteolysis. Mol. Aspects Med. 2014, 35, 1–71. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, I.; Mary, J.; Perichon, M.; Friguet, B. Rat peptide methionine sulphoxide reductase: Cloning of the cDNA, and down-regulation of gene expression and enzyme activity during aging. Biochem. J. 2001, 355, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Picot, C.R.; Perichon, M.; Cintrat, J.-C.; Friguet, B.; Petropoulos, I. The peptide methionine sulfoxide reductases, MsrA and MsrB (hCBS-1), are downregulated during replicative senescence of human WI-38 fibroblasts. FEBS Lett. 2004, 558, 74–78. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, E.K.; Rogowska-Wrzesinska, A.; Roepstorff, P.; Bulteau, A.L.; Friguet, B. Protein modification and replicative senescence of WI-38 human embryonic fibroblasts. Aging Cell 2010, 9, 252–272. [Google Scholar] [CrossRef] [Green Version]
- Brovelli, A.; Seppi, C.; Castellana, A.M.; De Renzis, M.R.; Blasina, A.; Balduini, C. Oxidative lesion to membrane proteins in senescent erythrocytes. Biomed. Biochim. Acta 1990, 49, S218–S223. [Google Scholar]
- Weissbach, H.; Resnick, L.; Brot, N. Methionine sulfoxide reductases: History and cellular role in protecting against oxidative damage. Biochim. Biophys. Acta 2005, 1703, 203–212. [Google Scholar] [CrossRef]
- Lee, B.C.; Lee, H.M.; Kim, S.; Avanesov, A.S.; Lee, A.; Chun, B.H.; Vorbruggen, G.; Gladyshev, V.N. Gladyshev Expression of the methionine sulfoxide reductase lost during evolution extends Drosophila lifespan in a methionine-dependent manner. Sci. Rep. 2018, 8, 1010. [Google Scholar] [CrossRef]
- Oien, D.B.; Moskovitz, J. Substrates of the methionine sulfoxide reductase system their physiological relevance. Curr. Top. Dev. Biol. 2008, 80, 93–133. [Google Scholar] [PubMed]
- Caldwell, P.; Luk, D.C.; Weissbach, H.; Brot, N. Oxidation of the methionine residues of Escherichia coli ribosomal protein L12 decreases the protein’s biological activity. Proc. Natl. Acad. Sci. USA 1978, 75, 5349–5352. [Google Scholar] [CrossRef] [PubMed]
- Ezraty, B.; Grimaud, R.; El Hassouni, M.; Moinier, D.; Barras, F. Methionine sulfoxide reductases protect Ffh from oxidative damages in Escherichia coli. EMBOJ 2004, 23, 1868–1877. [Google Scholar] [CrossRef] [PubMed]
- Abrams, W.R.; Weinbaum, G.; Weissbach, L.; Weissbach, H.; Brot, N. Enzymatic reduction of oxidized α-1-proteinase inhibitor restores biological activity. Proc. Natl. Acad. Sci. USA 1981, 78, 7483–7486. [Google Scholar] [CrossRef] [PubMed]
- Stief, T.W.; Aab, A.; Heimburger, N. Oxidative inactivation of purified human α-2-antiplasmin, antithrombin III, and C1-inhibitor. Thromb. Res. 1988, 49, 581–589. [Google Scholar] [CrossRef]
- Glaser, C.B.; Karic, L.; Parmelee, S.; Premachandra, B.R.; Hinkston, D.; Abrams, W.R. Studies on the turnover of methionine oxidized α-1-protease inhibitor in rats. Am. Rev. Respir. Dis. 1987, 136, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.A.; Newcomb, F.M.; Moskovitz, J.; Wingfield, P.T.; Stahl, S.J.; Kaufman, J.; Fales, H.M.; Levine, R.L.; Yarchoan, R. HIV-2 protease is inactivated after oxidation at the dimer interface activity can be partly restored with methionine sulphoxide reductase. Biochem. J. 2000, 2 Pt 346, 305–311. [Google Scholar] [CrossRef]
- Kanayama, A.; Inoue, J.; Sugita-Konishi, Y.; Shimizu, M.; Miyamoto, Y. Oxidation of Ikappa Bα at methionine 45 is one cause of taurine chloramine-induced inhibition of NF-kappa B activation. J. Biol. Chem. 2002, 277, 24049–24056. [Google Scholar] [CrossRef]
- Midwinter, R.G.; Cheah, F.C.; Moskovitz, J.; Vissers, M.C.; Winterbourn, C.C. IkappaB is a sensitive target for oxidation by cell-permeable chloramines: Inhibition of NF-kappaB activity by glycine chloramine through methionine oxidation. Biochem. J. 2006, 396, 71–78. [Google Scholar] [CrossRef]
- Mohri, M.; Reinach, P.S.; Kanayama, A.; Shimizu, M.; Moskovitz, J.; Hisatsune, T.; Miyamoto, Y. Suppression of the TNFα-induced increase in IL-1α expression by hypochlorite in human corneal epithelial cells. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3190–3195. [Google Scholar]
- Chen, J.; Avdonin, V.; Ciorba, M.A.; Heinemann, S.H.; Hoshi, T. Acceleration of P/C-type inactivation in voltage-gated K(+) channels by methionine oxidation. Biophys. J. 2000, 78, 174–187. [Google Scholar] [CrossRef]
- Ciorba, M.A.; Heinemann, S.H.; Weissbach, H.; Brot, N.; Hoshi, T. Modulation of potassium channel function by methionine oxidation reduction. Proc. Natl. Acad. Sci. USA 1997, 94, 9932–9937. [Google Scholar] [CrossRef] [PubMed]
- Bigelow, D.J.; Squier, T.C. Redox modulation of cellular signaling metabolism through reversible oxidation of methionine sensors in calcium regulatory proteins. Biochim. Biophys. Acta 2005, 1703, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.C.; Kim, G.; Levine, R.L. Stereospecific oxidation of calmodulin by methionine sulfoxide reductase A. Free Radic. Biol. Med. 2013, 61, 257–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Y.; Yin, D.; Jas, G.S.; Kuczer, K.; Williams, T.D.; Schoneich, C.; Squier, T.C. Oxidative modification of a carboxyl-terminal vicinal methionine in calmodulin by hydrogen peroxide inhibits calmodulin-dependent activation of the plasma membrane Ca-ATPase. Biochemistry 1996, 35, 2767–2787. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Gao, J.; Ferrington, D.A.; Biesiada, H.; Williams, T.D.; Squier, T.C. Repair of oxidized calmodulin by methionine sulfoxide reductase restores ability to activate the plasma membrane Ca-ATPase. Biochemistry 1999, 38, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Vougier, S.; Mary, J.; Dautin, N.; Vinh, J.; Friguet, B.; Ladant, D. Essential role of methionine residues in calmodulin binding to Bordetella pertussis adenylate cyclase, as probed by selective oxidation and repair by the peptide methionine sulfoxide reductases. J. Biol. Chem. 2004, 279, 30210–30218. [Google Scholar] [CrossRef]
- Erickson, J.R.; Joiner, M.L.; Guan, X.; Kutschke, W.; Yang, J.; Oddis, C.V.; Bartlett, R.K.; Lowe, J.S.; O’Donnell, S.E.; Aykin-Burns, N.; et al. A dynamic pathway for calcium-independent activation of CaMKII by methionine oxidation. Cell 2008, 133, 462–474. [Google Scholar] [CrossRef]
- He, B.J.; Joiner, M.L.; Singh, M.V.; Luczak, E.D.; Swaminathan, P.D.; Koval, O.M.; Kutschke, W.; Allamargot, C.; Yang, J.; Guan, X.; et al. Oxidation of CaMKII determines the cardiotoxic effects of aldosterone. Nat. Med. 2011, 17, 1610–1618. [Google Scholar] [CrossRef] [Green Version]
- Hung, R.J.; Yazdani, U.; Yoon, J.; Wu, H.; Yang, T.; Gupta, N.; Huang, Z.; van Berkel, W.J.; Terman, J.R. Mical links semaphorins to F-actin disassembly. Nature 2010, 463, 823–827. [Google Scholar] [CrossRef] [Green Version]
- Hung, R.J.; Spaeth, C.S.; Yesilyurt, H.G.; Terman, J.R. SelR reverses Mical-mediated oxidation of actin to regulate F-actin dynamics. Nat. Cell Biol. 2013, 15, 1445–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manta, B.; Gladyshev, V.N. Regulated methionine oxidation by monooxygenases. Free Radic. Biol. Med. 2017, 109, 141–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.M.; Kim, G.; Levine, R.L. Methionine in Proteins: It’s Not Just for Protein Initiation Anymore. Neurochem. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Wehr, N.B.; Levine, R.L. Wanted and wanting: Antibody against methionine sulfoxide. Free Radic. Biol. Med. 2012, 53, 1222–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghesquière, B.; Gevaert, K. Proteomics methods to study methionine oxidation. Mass Spectrom. Rev. 2014, 33, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Tarrago, L.; Oheix, E.; Péterfi, Z.; Gladyshev, V.N. Monitoring of methionine sulfoxide content and methionine sulfoxide reductase activity. Methods Mol. Biol. 2018, 1661, 285–299. [Google Scholar] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lourenço dos Santos, S.; Petropoulos, I.; Friguet, B. The Oxidized Protein Repair Enzymes Methionine Sulfoxide Reductases and Their Roles in Protecting against Oxidative Stress, in Ageing and in Regulating Protein Function. Antioxidants 2018, 7, 191. https://doi.org/10.3390/antiox7120191
Lourenço dos Santos S, Petropoulos I, Friguet B. The Oxidized Protein Repair Enzymes Methionine Sulfoxide Reductases and Their Roles in Protecting against Oxidative Stress, in Ageing and in Regulating Protein Function. Antioxidants. 2018; 7(12):191. https://doi.org/10.3390/antiox7120191
Chicago/Turabian StyleLourenço dos Santos, Sofia, Isabelle Petropoulos, and Bertrand Friguet. 2018. "The Oxidized Protein Repair Enzymes Methionine Sulfoxide Reductases and Their Roles in Protecting against Oxidative Stress, in Ageing and in Regulating Protein Function" Antioxidants 7, no. 12: 191. https://doi.org/10.3390/antiox7120191
APA StyleLourenço dos Santos, S., Petropoulos, I., & Friguet, B. (2018). The Oxidized Protein Repair Enzymes Methionine Sulfoxide Reductases and Their Roles in Protecting against Oxidative Stress, in Ageing and in Regulating Protein Function. Antioxidants, 7(12), 191. https://doi.org/10.3390/antiox7120191