Phytochemicals in Human Milk and Their Potential Antioxidative Protection


Abstract
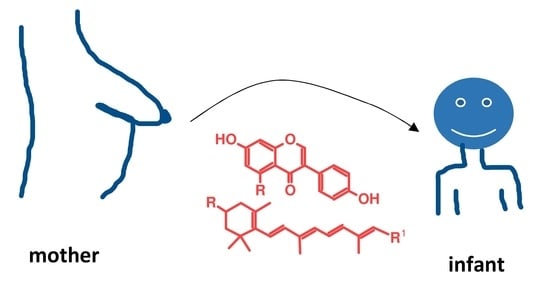
:
1. Introduction
2. Phytochemicals in Human Milk
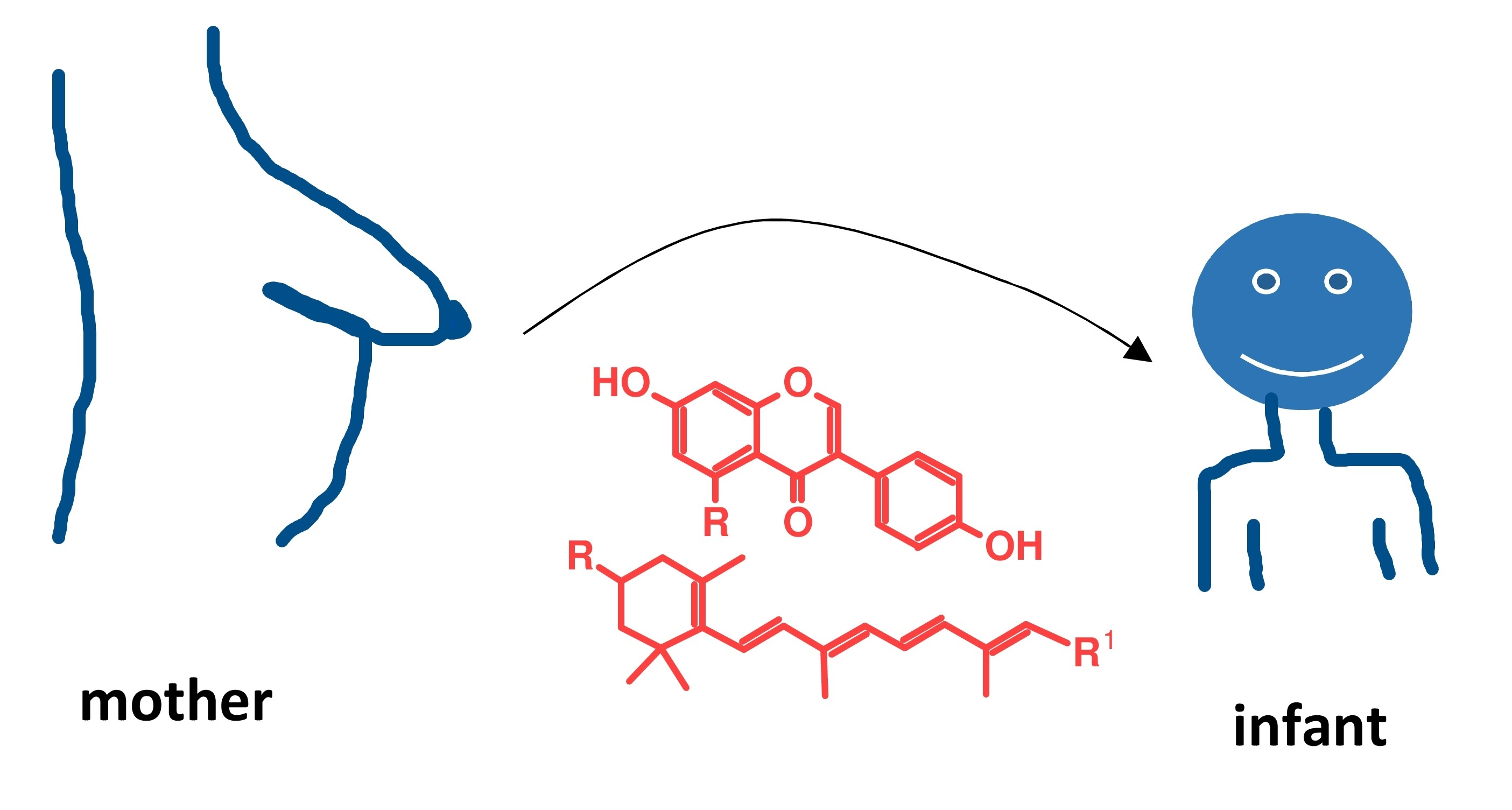
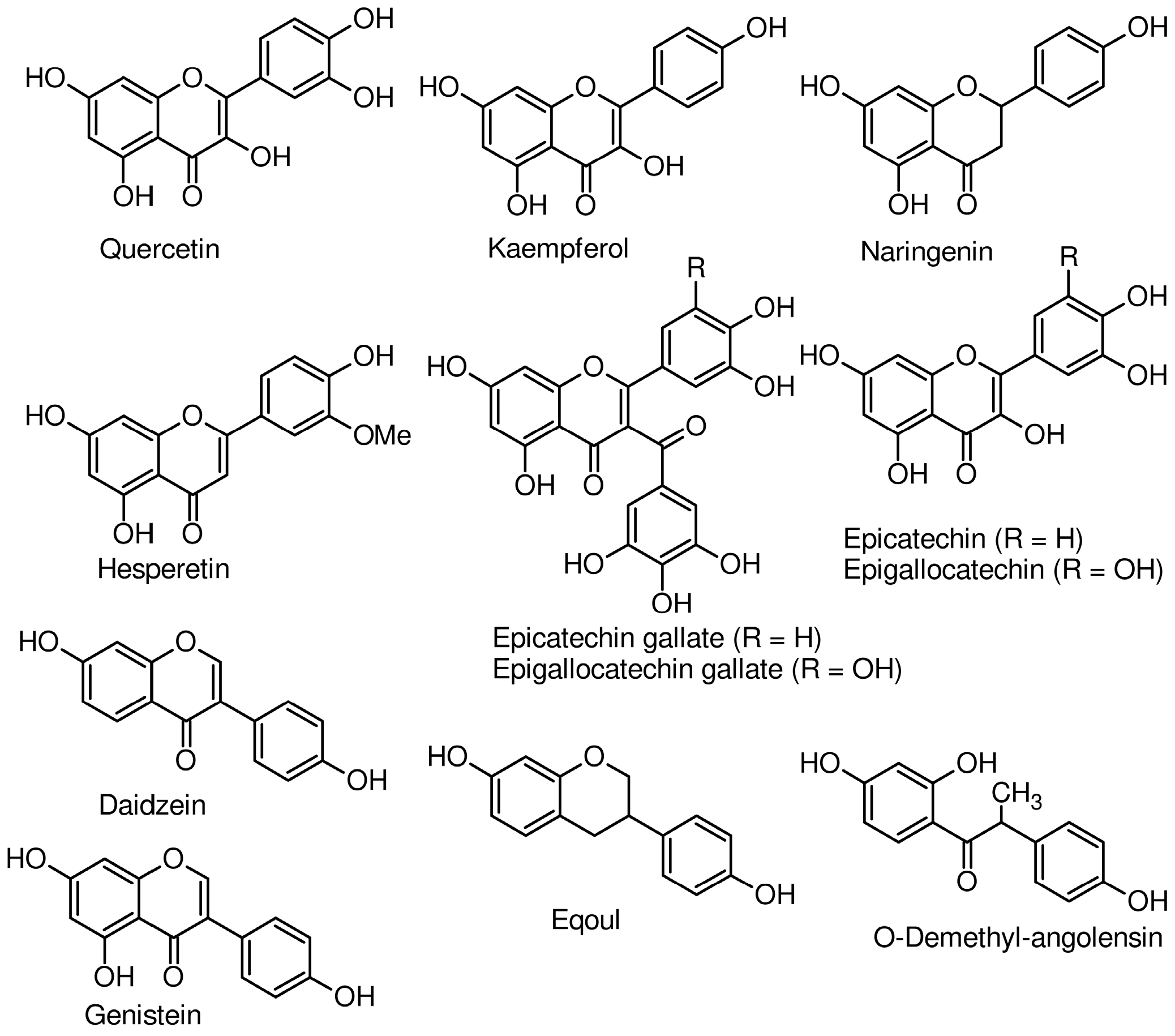
2.1. Flavonoids in Human Milk
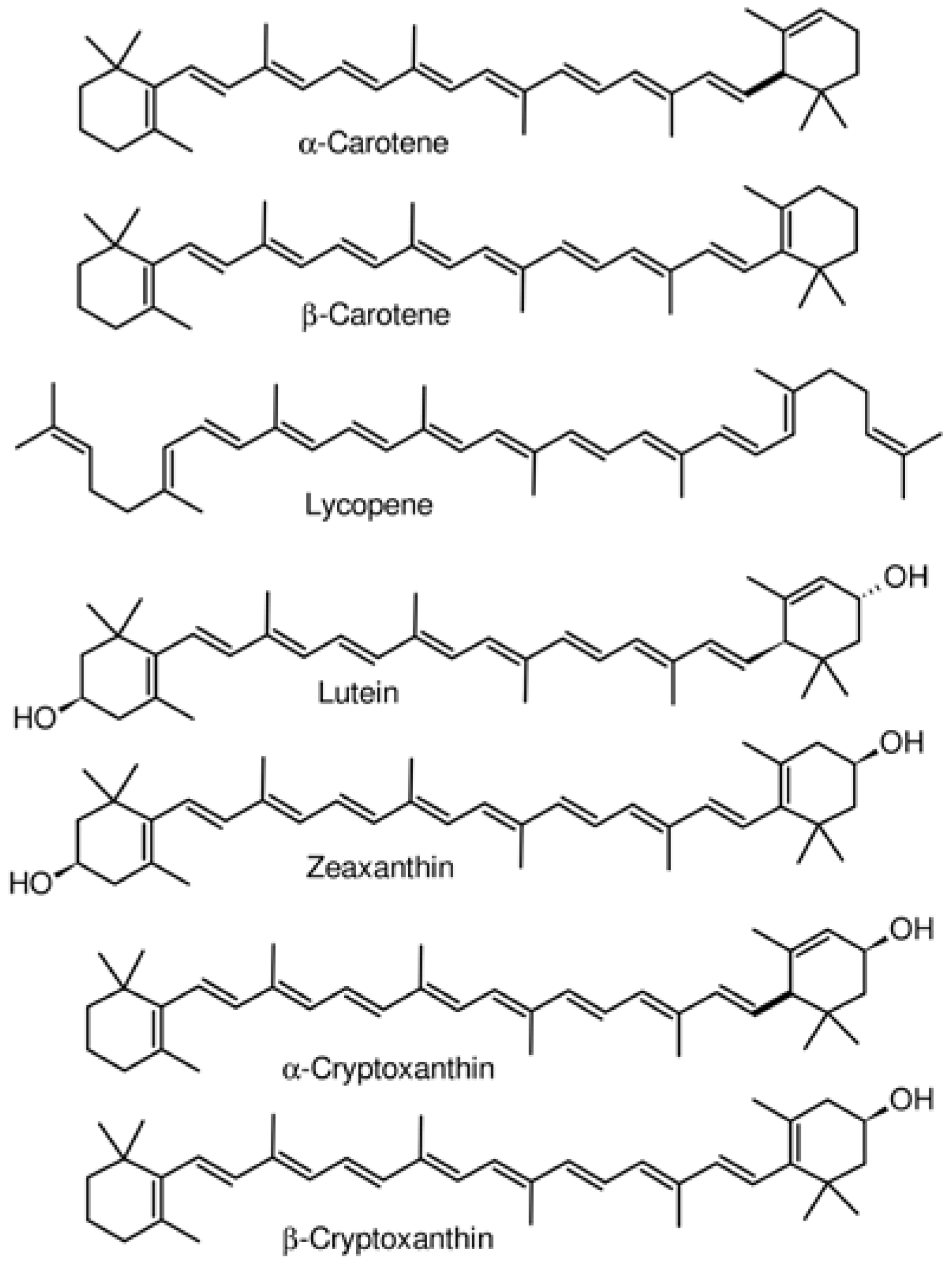
2.2. Carotenoids
2.3. Other Phytochemicals
3. Oxidative Stress in Infants
4. Antioxidant Phytochemicals in Human Milk and Redox Balance in Infants
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Fiedor, J.; Burda, K. Potential role of carotenoids as antioxidants in human health and disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, K.; Aggarwal, B.; Singh, R.; Buttar, H.; Wilson, D.; De Meester, F. Food antioxidants and their anti-inflammatory properties: A potential role in cardiovascular diseases and cancer prevention. Diseases 2016, 4, e28. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, S.I.; Sekiguchi, K.; Akaishi, M.; Anan, A.; Maeda, T.; Izumi, T. Characterization and chronological changes of preterm human milk gangliosides. Nutrition 2011, 27, 998–1001. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B. Bioactive proteins in human milk: Health, nutrition, and implications for infant formulas. J. Pediatr. 2016, 173, S4–S9. [Google Scholar] [CrossRef] [PubMed]
- Tsopmo, A.; Diehl-Jones, B.W.; Aluko, R.E.; Kitts, D.D.; Elisia, I.; Friel, J.K. Tryptophan released from mother’s milk has antioxidant properties. Pediatr. Res. 2009, 66, 614–618. [Google Scholar] [CrossRef] [PubMed]
- Tsopmo, A.; Romanowski, A.; Banda, L.; Lavoie, J.C.; Jenssen, H.; Friel, J.K. Novel anti-oxidative peptides from enzymatic digestion of human milk. Food Chem. 2011, 126, 1138–1143. [Google Scholar] [CrossRef]
- Burrin, D.G.; Stoll, B. Metabolic fate and function of dietary glutamate in the gut. Am. J. Clin. Nutr. 2009, 90, 850S–856S. [Google Scholar] [CrossRef] [PubMed]
- Kunz, C.; Rodriguez-Palmero, M.; Koletzko, B.; Jensen, R. Nutritional and biochemical properties of human milk, part I: General aspects, proteins, and carbohydrates. Clin. Perinatol. 1999, 26, 307–333. [Google Scholar] [PubMed]
- Jeong, K.; Nguyen, V.; Kim, J. Human milk oligosaccharides: The novel modulator of intestinal microbiota. BMB Rep. 2012, 45, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Smilowitz, J.T.; O’Sullivan, A.; Barile, D.; German, J.B.; Lönnerdal, B.; Slupsky, C.M. The human milk metabolome reveals diverse oligosaccharide profiles. J. Nutr. 2013, 143, 1709–1718. [Google Scholar] [CrossRef] [PubMed]
- Hallman, M.; Bry, K.; Hoppu, K.; Lappi, M.; Pohjavuori, M. Inositol supplementation in premature infants with respiratory distress syndrome. N. Engl. J. Med. 1992, 326, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Šantak, B.; Schreiber, M.; Kuen, P.; Lang, D.; Radermacher, P. Prostacyclin aerosol in an infant with pulmonary hypertension. Eur. J. Pediatr. 1995, 154, 233–235. [Google Scholar] [CrossRef] [PubMed]
- Neuringer, M. Infant vision and retinal function in studies of dietary long-chain polyunsaturated fatty acids: Methods, results, and implications. Am. J. Clin. Nutr. 2000, 71, 256S–267S. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Palmero, M.; Koletzko, B.; Kunz, C.; Jensen, R. Nutritional and biochemical properties of human milk: II. Lipids, micronutrients, and bioactive factors. Clin. Perinatol. 1999, 26, 335–359. [Google Scholar] [PubMed]
- Ratnasari, N.; Walters, M.; Tsopmo, A. Antioxidant and lipoxygenase activities of polyphenol extracts from oat brans treated with polysaccharide degrading enzymes. Heliyon 2017, 3. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Esfandi, R.; Willmore, W.G.; Tsopmo, A. Antioxidant activity of oat proteins derived peptides in stressed hepatic hepg2 cells. Antioxidants 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M. Updated Knowledge about Polyphenols: Functions, Bioavailability, Metabolism, and Health. Crit. Rev. Food Sci. Nutr. 2012, 52, 936–948. [Google Scholar] [CrossRef] [PubMed]
- Song, B.J.; Jouni, Z.E.; Ferruzzi, M.G. Assessment of phytochemical content in human milk during different stages of lactation. Nutrition 2013, 29, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Hughes, L.A.E.; Arts, I.C.W.; Ambergen, T.; Brants, H.A.M.; Dagnelie, P.C.; Goldbohm, R.A.; Van Den Brandt, P.A.; Weijenberg, M.P. Higher dietary flavone, flavonol, and catechin intakes are associated with less of an increase in BMI over time in women: A longitudinal analysis from The Netherlands Cohort Study. Am. J. Clin. Nutr. 2008, 88, 1341–1352. [Google Scholar] [CrossRef] [PubMed]
- Tsopmo, A.; Friel, J.K. Human milk has anti-oxidant properties to protect premature infants. Curr. Pediatr. Rev. 2007, 3, 45–51. [Google Scholar] [CrossRef]
- Miloudi, K.; Tsopmo, A.; Friel, J.K.; Rouleau, T.; Comte, B.; Lavoie, J.C. Hexapeptides from human milk prevent the induction of oxidative stress from parenteral nutrition in the newborn guinea pig. Pediatr. Res. 2012, 71, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Franke, A.A.; Custer, L.J.; Tanaka, Y. Isoflavones in human breast milk and other biological fluids. Am. J. Clin. Nutr. 1998, 68, 1466s–1473s. [Google Scholar] [CrossRef] [PubMed]
- Khymenets, O.; Rabassa, M.; Rodríguez-Palmero, M.; Rivero-Urgell, M.; Urpi-Sarda, M.; Tulipani, S.; Brandi, P.; Campoy, C.; Santos-Buelga, C.; Andres-Lacueva, C. Dietary epicatechin is available to breastfed infants through human breast milk in the form of host and microbial metabolites. J. Agric. Food Chem. 2016, 64. [Google Scholar] [CrossRef] [PubMed]
- Franke, A.A.; Halm, B.M.; Custer, L.J.; Tatsumura, Y.; Hebshi, S. Isoflavones in breastfed infants after mothers consume soy. Am. J. Clin. Nutr. 2006, 84, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Jochum, F.; Alteheld, B.; Meinardus, P.; Dahlinger, N.; Nomayo, A.; Stehle, P. Mothers’ Consumption of Soy Drink but Not Black Tea Increases the Flavonoid Content of Term Breast Milk: A Pilot Randomized, Controlled Intervention Study. Ann. Nutr. Metab. 2017, 70, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Romaszko, E.; Wiczkowski, W.; Romaszko, J.; Honke, J.; Piskula, M.K. Exposure of breastfed infants to quercetin after consumption of a single meal rich in quercetin by their mothers. Mol. Nutr. Food Res. 2014, 58, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Haftel, L.; Berkovich, Z.; Reifen, R. Elevated milk β-carotene and lycopene after carrot and tomato paste supplementation. Nutrition 2015, 31, 443–445. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, J.; Noda, K.; Uchikawa, T.; Maruyama, I.; Shimomura, H.; Miyahara, M. Effect of maternal Chlorella supplementation on carotenoid concentration in breast milk at early lactation. Int. J. Food Sci. Nutr. 2014, 65, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Hanson, C.; Lyden, E.; Furtado, J.; Van Ormer, M.; Anderson-Berry, A. A comparison of nutritional antioxidant content in breast milk, donor milk, and infant formulas. Nutrients 2016, 8, e681. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, D.L.; Ewaschuk, J.B.; Unger, S. Human milk pasteurization: Benefits and risks. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, T.; Nakagawa, K.; Kimura, F.; Nakashima, Y.; Maruyama, I.; Higuchi, O.; Miyazawa, T. Chlorella is an Effective Dietary Source of Lutein for Human Erythrocytes. J. Oleo Sci. 2013, 10, 905–914. [Google Scholar] [CrossRef]
- Cena, H.; Castellazzi, A.M.; Pietri, A.; Roggi, C.; Turconi, G. Lutein concentration in human milk during early lactation and its relationship with dietary lutein intake. Public Health Nutr. 2009, 12, 1878–1884. [Google Scholar] [CrossRef] [PubMed]
- Schweigert, F.J.; Bathe, K.; Chen, F.; Büscher, U.; Dudenhausen, J.W. Effect of the stage of lactation in humans on carotenoid levels in milk, blood plasma and plasma lipoprotein fractions. Eur. J. Nutr. 2004, 43, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.G.; Lien, E.L.; White, S.J.; Bruns, N.J.; Kuhlman, C.F. Major carotenoids in mature human milk: Longitudinal and diurnal patterns. J. Nutr. Biochem. 1998, 9, 2–7. [Google Scholar] [CrossRef]
- Canfield, L.M.; Clandinin, M.T.; Davies, D.P.; Fernandez, M.C.; Jackson, J.; Hawkes, J.; Goldman, W.J.; Pramuk, K.; Reyes, H.; Sablan, B.; et al. Multinational study of major breast milk carotenoids of healthy mothers. Eur. J. Nutr. 2003, 42, 133–141. [Google Scholar] [PubMed]
- Scheffler, L.; Sauermann, Y.; Zeh, G.; Hauf, K.; Heinlein, A.; Sharapa, C.; Buettner, A. Detection of volatile metabolites of garlic in human breast milk. Metabolites 2016, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Amorati, R.; Pedulli, G.F. Do garlic-derived allyl sulfides scavenge peroxyl radicals? Org. Biomol. Chem. 2008, 6, 1103–1107. [Google Scholar] [CrossRef] [PubMed]
- Aresta, A.; Palmisano, F.; Zambonin, C.G. Simultaneous determination of caffeine, theobromine, theophylline, paraxanthine and nicotine in human milk by liquid chromatography with diode array UV detection. Food Chem. 2005, 93, 177–181. [Google Scholar] [CrossRef]
- Azam, S.; Hadi, N.; Khan, N.U.; Hadi, S.M. Antioxidant and prooxidant properties of caffeine, theobromine and xanthine. Med. Sci. Monit. 2003, 9, BR325–BR330. [Google Scholar] [PubMed]
- Gitto, E.; Pellegrino, S.; Gitto, P.; Barberi, I.; Reiter, R.J. Oxidative stress of the newborn in the pre- and postnatal period and the clinical utility of melatonin. J. Pineal Res. 2009, 46, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Elisia, I.; Tsopmo, A.; Friel, J.K.; Diehl-Jones, W.; Kitts, D.D. Tryptophan from human milk induces oxidative stress and upregulates the Nrf-2-mediated stress response in human intestinal cell lines. J. Nutr. 2011, 141, 1417–1423. [Google Scholar] [CrossRef] [PubMed]
- Friel, J.K.; Diehl-Jones, W.L.; Suh, M.; Tsopmo, A.; Shirwadkar, V.P. Impact of iron and vitamin C-containing supplements on preterm human milk: in vitro. Free Radic. Biol. Med. 2007, 42, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Friel, J.K.; Friesen, R.W.; Harding, S.V.; Roberts, L.J. Evidence of oxidative stress in full-term healthy infants. Pediatr. Res. 2004, 56, 878–882. [Google Scholar] [CrossRef] [PubMed]
- Negi, R.; Pande, D.; Kumar, A.; Khanna, R.S.; Khanna, H.D. In vivo oxidative DNA damage and lipid peroxidation as a biomarker of oxidative stress in preterm low-birthweight infants. J. Trop. Pediatr. 2012, 58, 326–328. [Google Scholar] [CrossRef] [PubMed]
- Gladstone, I.M.; Levine, R.L. Oxidation of Proteins in Neonatal Lungs. Pediatrics 1994, 93, 764–768. [Google Scholar] [PubMed]
- Lubec, G.; Widness, J.A.; Hayde, M.; Menzel, D.; Pollak, A. Hydroxyl radical generation in oxygen-treated infants. Pediatrics 1997, 100, 700–704. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.; Grune, T.; Muller, R.; Siems, W.G.; Wauer, R.R. Increased levels of lipid peroxidation products malondialdehyde and 4-hydroxynonenal after perinatal hypoxia. Pediatr. Res. 1996, 40, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Ambalavanan, N.; Carlo, W.A. Bronchopulmonary dysplasia: New insights. Clin. Perinatol. 2004, 31, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, C.; Dilli, D.; Uras, N.; Ulu, H.O.; Oguz, S.S.; Erdeve, O.; Dilmen, U. Total oxidant status and oxidative stress are increased in infants with necrotizing enterocolitis. J. Pediatr. Surg. 2011, 46, 2096–2100. [Google Scholar] [CrossRef] [PubMed]
- Hellström, A.; Smith, L.E.; Dammann, O. Retinopathy of prematurity. Lancet 2013, 382, 1445–1457. [Google Scholar] [CrossRef]
- Ozsurekci, Y.; Aykac, K. Oxidative stress related diseases in newborns. Oxid. Med. Cell. Longev. 2016, 2016, 2768365. [Google Scholar] [CrossRef] [PubMed]
- Friel, J.K.; Martin, S.M.; Langdon, M.; Herzberg, G.R.; Buettner, G.R. Milk from mothers of both premature and full-term infants provides better antioxidant protection than does infant formula. Pediatr. Res. 2002, 51, 612–618. [Google Scholar] [CrossRef] [PubMed]
- L’Abbe, M.R.; Friel, J.K. Copper status of very low birth weight infants during the first 12 months of infancy. Pediatr. Res. 1992, 32, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Celep, G.S.; Rastmanesh, R.; Marotta, F. Polyphenols in Human Health and Disease. In Polyphenols in Human Health and Disease; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Academic Press, Elsevier Inc.: Tokyo, Japan, 2014; pp. 577–589. ISBN 9780123984562. [Google Scholar]
- Poniedziałek, B.; Rzymski, P.; Pięt, M.; Gąsecka, M.; Stroińska, A.; Niedzielski, P.; Mleczek, M.; Rzymski, P.; Wilczak, M. Relation between polyphenols, malondialdehyde, antioxidant capacity, lactate dehydrogenase and toxic elements in human colostrum milk. Chemosphere 2018, 191, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Matos, C.; Ribeiro, M.; Guerra, A. Breastfeeding: Antioxidative properties of breast milk. J. Appl. Biomed. 2015, 13, 169–180. [Google Scholar] [CrossRef]
- Tijerina-Saenz, A.; Innis, S.M.; Kitts, D.D. Antioxidant capacity of human milk and its association with vitamins A and E and fatty acid composition. Acta Paediatr. 2009. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, B.S.; Chan, G.; Hoffman, R.O.; Sharifzadeh, M.; Ermakov, I.V.; Gellermann, W.; Bernstein, P.S. Interrelationships between maternal carotenoid status and newborn infant macular pigment optical density and carotenoid status. Investig. Ophthalmol. Vis. Sci. 2013, 54, 5568–5578. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, J.P.; Hammond, B.R. Possible influences of lutein and zeaxanthin on the developing retina. Clin. Ophthalmol. 2007, 1, 25–35. [Google Scholar] [PubMed]
- Perrone, S.; Longini, M.; Marzocchi, B.; Picardi, A.; Bellieni, C.V.; Proietti, F.; Rodriguez, A.; Turrisi, G.; Buonocore, G. Effects of lutein on oxidative stress in the term newborn: A pilot study. Neonatology 2010, 97, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Rubin, L.P.; Chan, G.M.; Barrett-Reis, B.M.; Fulton, A.B.; Hansen, R.M.; Ashmeade, T.L.; Oliver, J.S.; MacKey, A.D.; Dimmit, R.A.; Hartmann, E.E.; et al. Effect of carotenoid supplementation on plasma carotenoids, inflammation and visual development in preterm infants. J. Perinatol. 2012, 32, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Kent, J.C.; Mitoulas, L.R.; Cregan, M.D.; Ramsay, D.T.; Doherty, D.A.; Hartmann, P.E. Volume and frequency of breastfeedings and fat content of breast milk throughout the day. Pediatrics 2006, 117, e387–e395. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D.R.; Zimmer-Nechemias, L.; Cai, J.; Heubi, J.E. Isoflavone content of infant formulas and the metabolic fate of these phytoestrogens in early life. Am. J. Clin. Nutr. 1998, 68, 1453S–1461S. [Google Scholar] [CrossRef] [PubMed]
- Hoey, L.; Rowland, I.R.; Lloyd, A.S.; Clarke, D.B.; Wiseman, H. Influence of soya-based infant formula consumption on isoflavone and gut microflora metabolite concentrations in urine and on faecal microflora composition and metabolic activity in infants and children. Br. J. Nutr. 2004, 91, 607–616. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Compound | Concentration (nmol/L) | Information on Mothers and Milk |
|---|---|---|
| Epicatechin | 63.7–828.5 | Free living mothers, milk at 1, 4 and 13 week [18] |
| Epicatechin gallate | 55.7–645.6 | |
| Epigallocatechin gallate | 215.1–2364.7 | |
| Naringenin | 64.1–722.0 | |
| Kaempferol | 7.8–71.4 | |
| Hesperetin | 74.8–1603.1 | |
| Quercetin | 32.5–108.6 | Free living mothers, milk at 1, 4 and 13 week [18] |
| 68 ± 8.44 | Diet with 1 mg quercetin/kg of body weight [26] | |
| Lutein | 56.4–121.2 | Free living mothers, milk analyzed at 1, 4 and 13 weeks [18] |
| 497–824 | Chlorella supplementation, 6 months from gestational week 16–20 until delivery [28] | |
| 280 ± 22 | Free living mothers. Milk collected at day 3 [32] | |
| Zeaxanthin | 46.3–21.4 | Free living mothers, milk at 1, 4 and 13 weeks [18] |
| 33.2±17.2 | Healthy women. Milk collected at days 2–6 [33] | |
| α-Cryptoxanthin | 13.5–30.6 | Free living mothers, milk at 1, 4 and 13 weeks [18] |
| β-Cryptoxanthin | 24.8–57.4 | Free living mothers, milk at 1–14 weeks [18,34] |
| α-Carotene | 23.2–59.0 | Free living mothers, milk at 1, 4 and 13 weeks [18] |
| β-Carotene | 88.0–164.3 | Free living mothers, milk at 1–14 weeks [18,34] |
| 75–400 | Supplementation, 30 mg β-carotene/d for 28 days [35] | |
| 275–484 | Chlorella supplementation 6 months, from gestational week 16–20 until day of delivery [28] | |
| Lycopene | 119.9–49.5 | Free living mothers, milk analyzed at 1, 4 and 13 weeks [18,34] |
| 86–244 | Chlorella supplementation for 6 months, from gestational week 16–20 until day of delivery. Milk collected at 1–6 days [28] | |
| Isoflavones | 70.7 ± 19.2 | Soy beverage with 55 mg isoflavones daily for 2–4 days [24] |
| 12.0 | Soy drink, 12 mg isoflavones daily for 6 days [25] | |
| Epicat-Gluc-4 * | 0.0–36.4 | Free living mothers, milk collected at 1–30 days [23] |
| Epicat-Sulf-3 * | 0.0–14.5 | |
| MetEpicat-Sulf-3 * | 0.0–23.7 | |
| Caffeine ** | 0.06–0.77 | Milk of habitual coffee and chocolate mothers [34] |
| Theobromine | 0.08–0.50 ** | |
| Paraxanthine | 0.15–1.68 ** | |
| Theophylline | 0.10–0.66 ** |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsopmo, A. Phytochemicals in Human Milk and Their Potential Antioxidative Protection. Antioxidants 2018, 7, 32. https://doi.org/10.3390/antiox7020032
Tsopmo A. Phytochemicals in Human Milk and Their Potential Antioxidative Protection. Antioxidants. 2018; 7(2):32. https://doi.org/10.3390/antiox7020032
Chicago/Turabian StyleTsopmo, Apollinaire. 2018. "Phytochemicals in Human Milk and Their Potential Antioxidative Protection" Antioxidants 7, no. 2: 32. https://doi.org/10.3390/antiox7020032
APA StyleTsopmo, A. (2018). Phytochemicals in Human Milk and Their Potential Antioxidative Protection. Antioxidants, 7(2), 32. https://doi.org/10.3390/antiox7020032