The Energy Costs of Prematurity and the Neonatal Intensive Care Unit (NICU) Experience


Abstract
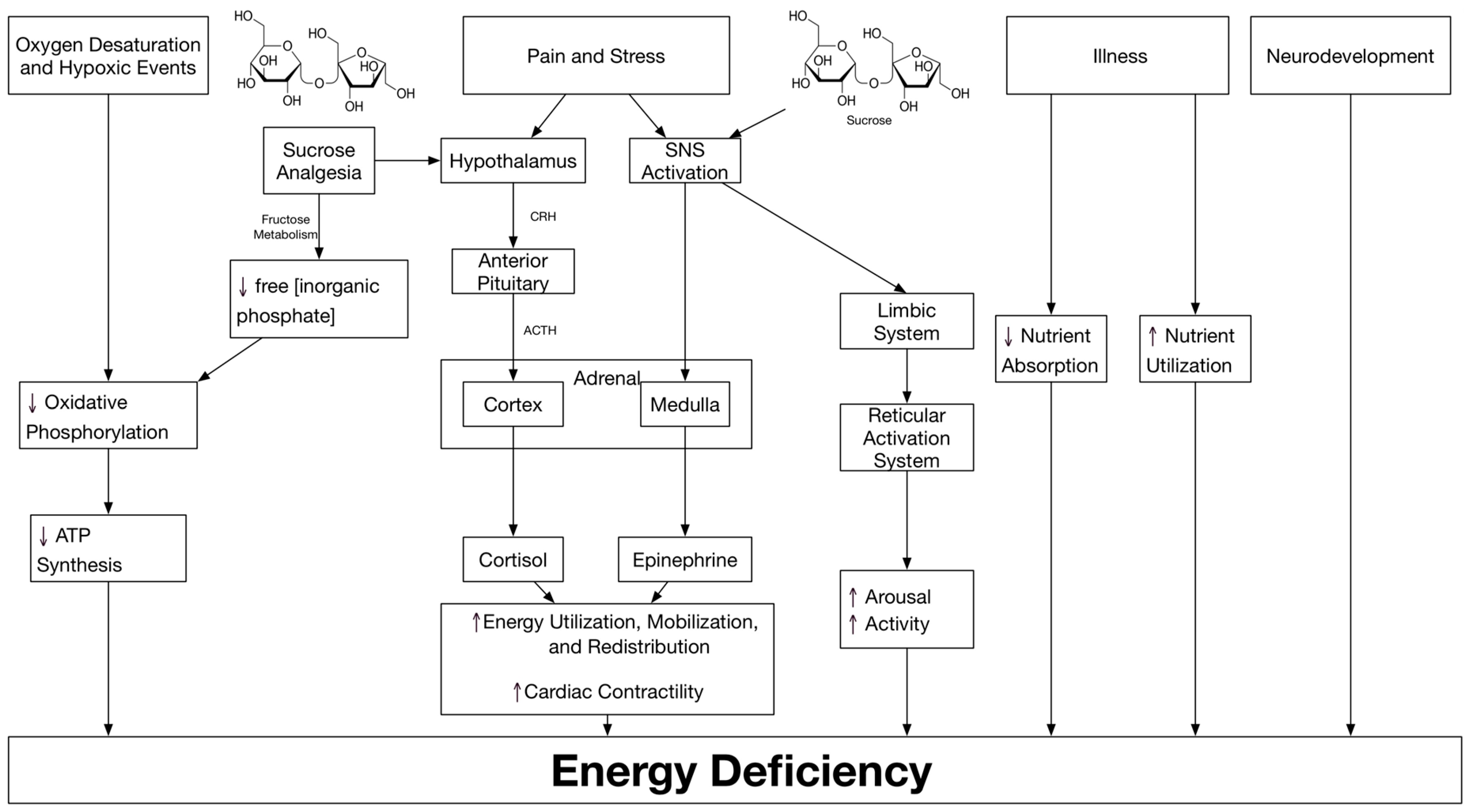
:1. Introduction
2. Premature Infants Are in an Energy-Deficit State
3. Energy Deficiency Affects Long-Term Outcomes
4. Prematurity and Chronic Stress, Energy Deficiency and Neuroplasticity
5. Sucrose and Stress Relief
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dinerstein, A.; Nieto, R.M.; Solana, C.L.; Perez, G.P.; Otheguy, L.E.; Larguia, A.M. Early and aggressive nutritional strategy (parenteral and enteral) decreases postnatal growth failure in very low birth weight infants. J. Perinatol. 2006, 26, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Ramel, S.E.; Brown, L.D.; Georgieff, M.K. The Impact of Neonatal Illness on Nutritional Requirements—One Size Does Not Fit All. Curr. Pediatr. Rep. 2014, 2, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Ramel, S.E.; Demerath, E.W.; Gray, H.L.; Younge, N.; Boys, C.; Georgieff, M.K. The relationship of poor linear growth velocity with neonatal illness and two-year neurodevelopment in preterm infants. Neonatology 2012, 102, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Ehrenkranz, R.A.; Dusick, A.M.; Vohr, B.R.; Wright, L.L.; Wrage, L.A.; Poole, W.K. Growth in the neonatal intensive care unit influences neurodevelopmental and growth outcomes of extremely low birth weight infants. Pediatrics 2006, 117, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Lapillonne, A. Feeding the preterm infant after discharge. World Rev. Nutr. Diet. 2014, 110, 264–277. [Google Scholar] [CrossRef] [PubMed]
- Vargas, J.; Junco, M.; Gomez, C.; Lajud, N. Early Life Stress Increases Metabolic Risk, HPA Axis Reactivity and Depressive-Like Behavior When Combined with Postweaning Social Isolation in Rats. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Stevens, B.; Yamada, J.; Ohlsson, A.; Haliburton, S.; Shorkey, A. Sucrose for analgesia (pain relief) in newborn infants undergoing painful procedures. Cochrane Database Syst. Rev. 2016, 7. [Google Scholar] [CrossRef]
- Carbajal, R.; Rousset, A.; Danan, C.; Coquery, S.; Nolent, P.; Ducrocq, S.; Saizou, C.; Lapillonne, A.; Granier, M.; Durand, P.; et al. Epidemiology and treatment of painful procedures in neonates in intensive care units. JAMA 2008, 300, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Angeles, D.M.; Ashwal, S.; Wycliffe, N.D.; Ebner, C.; Fayard, E.; Sowers, L.; Holshouser, B.A. Relationship between opioid therapy, tissue-damaging procedures and brain metabolites as measured by proton MRS in asphyxiated term neonates. Pediatr. Res. 2007, 61, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Asmerom, Y.; Slater, L.; Boskovic, D.S.; Bahjri, K.; Holden, M.S.; Phillips, R.; Deming, D.; Ashwal, S.; Fayard, E.; Angeles, D.M. Oral sucrose for heel lance increases adenosine triphosphate use and oxidative stress in preterm neonates. J. Pediatr. 2013, 163, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Holsti, L.; Grunau, R.E. Considerations for using sucrose to reduce procedural pain in preterm infants. Pediatrics 2010, 125, 1042–1047. [Google Scholar] [CrossRef] [PubMed]
- Stevens, B.; Yamada, J.; Beyene, J.; Gibbins, S.; Petryshen, P.; Stinson, J.; Narciso, J. Consistent management of repeated procedural pain with sucrose in preterm neonates: Is it effective and safe for repeated use over time? Clin. J. Pain 2005, 21, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Taddio, A.; Shah, V.; Atenafu, E.; Katz, J. Influence of repeated painful procedures and sucrose analgesia on the development of hyperalgesia in newborn infants. Pain 2009, 144, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Muralidhara, D.V.; Shetty, P.S. Sucrose feeding stimulates basal metabolism & nonshivering thermogenesis in undernourished rats. Indian J. Med. Res. 1990, 92, 447–451. [Google Scholar] [PubMed]
- Laugero, K.D. A new perspective on glucocorticoid feedback: Relation to stress, carbohydrate feeding and feeling better. J. Neuroendocrinol. 2001, 13, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Laugero, K.D. Reinterpretation of basal glucocorticoid feedback: Implications to behavioral and metabolic disease. Vitam. Horm. 2004, 69, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Goran, M.I.; Dumke, K.; Bouret, S.G.; Kayser, B.; Walker, R.W.; Blumberg, B. The obesogenic effect of high fructose exposure during early development. Nat. Rev. Endocrinol. 2013, 9, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Tappy, L.; Egli, L.; Lecoultre, V.; Schneider, P. Effects of fructose-containing caloric sweeteners on resting energy expenditure and energy efficiency: A review of human trials. Nutr. Metab. 2013, 10, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldenberg, R.L.; Culhane, J.F.; Iams, J.D.; Romero, R. Epidemiology and causes of preterm birth. Lancet 2008, 371, 75–84. [Google Scholar] [CrossRef]
- Tchirikov, M.; Zhumadilov, Z.S.; Bapayeva, G.; Bergner, M.; Entezami, M. The effect of intraumbilical fetal nutrition via a subcutaneously implanted port system on amino acid concentration by severe IUGR human fetuses. J. Perinat. Med. 2017, 45, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Denne, S.C. Protein and energy requirements in preterm infants. Semin. Neonatol. SN 2001, 6, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Fairchild, K.; Mohr, M.; Paget-Brown, A.; Tabacaru, C.; Lake, D.; Delos, J.; Moorman, J.R.; Kattwinkel, J. Clinical associations of immature breathing in preterm infants: Part 1—Central apnea. Pediatr. Res. 2016, 80, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Plank, M.S.; Boskovic, D.S.; Sowers, L.C.; Angeles, D.M. Biochemical markers of neonatal hypoxia. Pediatr. Health 2008, 2, 485–501. [Google Scholar] [CrossRef]
- Michiels, C. Physiological and pathological responses to hypoxia. Am. J. Pathol. 2004, 164, 1875–1882. [Google Scholar] [CrossRef]
- Slater, L.; Asmerom, Y.; Boskovic, D.S.; Bahjri, K.; Plank, M.S.; Angeles, K.R.; Phillips, R.; Deming, D.; Ashwal, S.; Hougland, K.; et al. Procedural pain and oxidative stress in premature neonates. J. Pain Off. J. Am. Pain Soc. 2012, 13, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Holsti, L.; Grunau, R.E.; Oberlander, T.F.; Whitfield, M.F.; Weinberg, J. Body movements: An important additional factor in discriminating pain from stress in preterm infants. Clin. J. Pain 2005, 21, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Flatters, S.J. The Contribution of Mitochondria to Sensory Processing and Pain. Prog. Mol. Biol. Transl. Sci. 2015, 131, 119–146. [Google Scholar] [PubMed]
- Steinhorn, D.M.; Green, T.P. Severity of illness correlates with alterations in energy metabolism in the pediatric intensive care unit. Crit. Care Med. 1991, 19, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Cerra, F.B.; Siegel, J.H.; Coleman, B.; Border, J.R.; McMenamy, R.R. Septic autocannibalism. A failure of exogenous nutritional support. Ann. Surg. 1980, 192, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Mehta, N.M.; Duggan, C.P. Nutritional Deficiencies during Critical Illness. Pediatr. Clin. N. Am. 2009, 56, 1143–1160. [Google Scholar] [CrossRef] [PubMed]
- Dao, D.T.; Anez-Bustillos, L.; Cho, B.S.; Li, Z.; Puder, M.; Gura, K.M. Assessment of Micronutrient Status in Critically Ill Children: Challenges and Opportunities. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- De Albuquerque Wilasco, M.I.; Uribe-Cruz, C.; Santetti, D.; Fries, G.R.; Dornelles, C.T.L.; da Silveira, T.R. IL-6, TNF-α, IL-10 and nutritional status in pediatric patients with biliary atresia. J. Pediatr. (Rio J.) 2017, 93, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Harris, M.C.; Costarino, A.T.; Sullivan, J.S.; Dulkerian, S.; McCawley, L.; Corcoran, L.; Butler, S.; Kilpatrick, L. Cytokine elevations in critically ill infants with sepsis and necrotizing enterocolitis. J. Pediatr. 1994, 124, 105–111. [Google Scholar] [CrossRef]
- Harris, J.J.; Jolivet, R.; Attwell, D. Synaptic energy use and supply. Neuron 2012, 75, 762–777. [Google Scholar] [CrossRef] [PubMed]
- Brummelte, S.; Grunau, R.E.; Chau, V.; Poskitt, K.J.; Brant, R.; Vinall, J.; Gover, A.; Synnes, A.R.; Miller, S.P. Procedural pain and brain development in premature newborns. Ann. Neurol. 2012, 71, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.P.; Ferriero, D.M. From selective vulnerability to connectivity: Insights from newborn brain imaging. Trends Neurosci. 2009, 32, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Kuzawa, C.W. Adipose tissue in human infancy and childhood: An evolutionary perspective. Am. J. Phys. Anthropol. 1998, 27, 177–209. [Google Scholar] [CrossRef]
- Harris, J.J.; Reynell, C.; Attwell, D. The physiology of developmental changes in BOLD functional imaging signals. Dev. Cogn. Neurosci. 2011, 1, 199–216. [Google Scholar] [CrossRef] [PubMed]
- Hay, W.W.; Brown, L.D.; Denne, S.C. Energy requirements, protein-energy metabolism and balance and carbohydrates in preterm infants. World Rev. Nutr. Diet. 2014, 110, 64–81. [Google Scholar] [CrossRef] [PubMed]
- Neu, J. Gastrointestinal development and meeting the nutritional needs of premature infants. Am. J. Clin. Nutr. 2007, 85, 629S–634S. [Google Scholar] [CrossRef] [PubMed]
- Burrin, D.G.; Stoll, B. Key nutrients and growth factors for the neonatal gastrointestinal tract. Clin. Perinatol. 2002, 29, 65–96. [Google Scholar] [CrossRef]
- Jacobi, S.K.; Odle, J. Nutritional Factors Influencing Intestinal Health of the Neonate. Adv. Nutr. Int. Rev. J. 2012, 3, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Stoll, B.; Horst, D.A.; Cui, L.; Chang, X.; Ellis, K.J.; Hadsell, D.L.; Suryawan, A.; Kurundkar, A.; Maheshwari, A.; Davis, T.A.; et al. Chronic Parenteral Nutrition Induces Hepatic Inflammation, Steatosis and Insulin Resistance in Neonatal Pigs. J. Nutr. 2010, 140, 2193–2200. [Google Scholar] [CrossRef] [PubMed]
- Tappenden, K.A. Mechanisms of enteral nutrient-enhanced intestinal adaptation. Gastroenterology 2006, 130, S93–S99. [Google Scholar] [CrossRef] [PubMed]
- Reali, A.; Greco, F.; Marongiu, G.; Deidda, F.; Atzeni, S.; Campus, R.; Dessì, A.; Fanos, V. Individualized fortification of breast milk in 41 Extremely Low Birth Weight (ELBW) preterm infants. Clin. Chim. Acta 2015, 451, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.M.; Jeroudi, M.A.; Baier, R.J.; Dhanireddy, R.; Krouskop, R.W. Aggressive early total parental nutrition in low-birth-weight infants. J. Perinatol. Off. J. Calif. Perinat. Assoc. 2004, 24, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Shalak, L.; Perlman, J.M. Hypoxic-ischemic brain injury in the term infant-current concepts. Early Hum. Dev. 2004, 80, 125–141. [Google Scholar] [CrossRef] [PubMed]
- Georgieff, M.K.; Brunette, K.E.; Tran, P.V. Early life nutrition and neural plasticity. Dev. Psychopathol. 2015, 27, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Hensch, T.K. Critical period regulation. Annu. Rev. Neurosci. 2004, 27, 549–579. [Google Scholar] [CrossRef] [PubMed]
- Ramel, S.E.; Georgieff, M.K. Preterm nutrition and the brain. World Rev. Nutr. Diet. 2014, 110, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Stephens, B.E.; Walden, R.V.; Gargus, R.A.; Tucker, R.; McKinley, L.; Mance, M.; Nye, J.; Vohr, B.R. First-week protein and energy intakes are associated with 18-month developmental outcomes in extremely low birth weight infants. Pediatrics 2009, 123, 1337–1343. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, E.B.; Morley, R.; Lucas, A. Early diet and general cognitive outcome at adolescence in children born at or below 30 weeks gestation. J. Pediatr. 2009, 155, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Fenton, T.R.; Anderson, D.; Groh-Wargo, S.; Hoyos, A.; Ehrenkranz, R.A.; Senterre, T. An Attempt to Standardize the Calculation of Growth Velocity of Preterm Infants-Evaluation of Practical Bedside Methods. J. Pediatr. 2017. [Google Scholar] [CrossRef] [PubMed]
- Holden, M.S.; Hopper, A.; Slater, L.; Asmerom, Y.; Esiaba, I.; Boskovic, D.S.; Angeles, D.M. Urinary Hypoxanthine as a Measure of Increased ATP Utilization in Late Preterm Infants. ICAN Infant Child Adolesc. Nutr. 2014. [Google Scholar] [CrossRef] [PubMed]
- Esiaba, I.; Angeles, D.M.; Holden, M.S.; Tan, J.B.C.; Asmerom, Y.; Gollin, G.; Boskovic, D.S. Urinary Allantoin Is Elevated in Severe Intraventricular Hemorrhage in the Preterm Newborn. Transl. Stroke Res. 2016, 7, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Sammallahti, S.; Kajantie, E.; Matinolli, H.-M.; Pyhälä, R.; Lahti, J.; Heinonen, K.; Lahti, M.; Pesonen, A.-K.; Eriksson, J.G.; Hovi, P.; et al. Nutrition after preterm birth and adult neurocognitive outcomes. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Connelly, A.; Bland, R.M.; Reilly, J.J. Health impact of catch-up growth in low-birth weight infants: Systematic review, evidence appraisal and meta-analysis. Matern. Child. Nutr. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, M.S.; Valentine, C.J. Neonatal Body Composition: Measuring Lean Mass as a Tool to Guide Nutrition Management in the Neonate. Nutr. Clin. Pract. Off. Publ. Am. Soc. Parenter. Enter. Nutr. 2015, 30, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Raaijmakers, A.; Allegaert, K. Catch-Up Growth in Former Preterm Neonates: No Time to Waste. Nutrients 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Jaquet, D.; Deghmoun, S.; Chevenne, D.; Collin, D.; Czernichow, P.; Lévy-Marchal, C. Dynamic change in adiposity from fetal to postnatal life is involved in the metabolic syndrome associated with reduced fetal growth. Diabetologia 2005, 48, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Hay, W.W. Aggressive Nutrition of the Preterm Infant. Curr. Pediatr. Rep. 2013, 1. [Google Scholar] [CrossRef] [PubMed]
- Victora, C.G.; Barros, F.C.; Horta, B.L.; Martorell, R. Short-term benefits of catch-up growth for small-for-gestational-age infants. Int. J. Epidemiol. 2001, 30, 1325–1330. [Google Scholar] [CrossRef] [PubMed]
- Jain, V.; Singhal, A. Catch up growth in low birth weight infants: Striking a healthy balance. Rev. Endocr. Metab. Disord. 2012, 13, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Ramel, S.E.; Gray, H.L.; Ode, K.L.; Younge, N.; Georgieff, M.K.; Demerath, E.W. Body composition changes in preterm infants following hospital discharge: Comparison with term infants. J. Pediatr. Gastroenterol. Nutr. 2011, 53, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Olhager, E.; Törnqvist, C. Body composition in late preterm infants in the first 10 days of life and at full term. Acta Paediatr. 2014, 103, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.J.; Wootton, S.A.; Leaf, A.A.; Jackson, A.A. Preterm birth and body composition at term equivalent age: A systematic review and meta-analysis. Pediatrics 2012, 130, e640–e649. [Google Scholar] [CrossRef] [PubMed]
- Franco, L.P.; Morais, C.C.; Cominetti, C. Normal-weight obesity syndrome: Diagnosis, prevalence and clinical implications. Nutr. Rev. 2016, 74, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Rigo, J.; de Curtis, M.; Pieltain, C. Nutritional assessment in preterm infants with special reference to body composition. Semin. Neonatol. 2001, 6, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Griffin, I.J. Nutritional assessment in preterm infants. Nestlé Nutr. Workshop Ser. Paediatr. Programme 2007, 59, 177–192. [Google Scholar] [CrossRef]
- Lapillonne, A.; Griffin, I.J. Feeding preterm infants today for later metabolic and cardiovascular outcomes. J. Pediatr. 2013, 162, S7–S16. [Google Scholar] [CrossRef] [PubMed]
- Embleton, N.D.; Korada, M.; Wood, C.L.; Pearce, M.S.; Swamy, R.; Cheetham, T.D. Catch-up growth and metabolic outcomes in adolescents born preterm. Arch. Dis. Child. 2016. [Google Scholar] [CrossRef] [PubMed]
- Peng, N.-H.; Bachman, J.; Chen, C.-H.; Huang, L.-C.; Lin, H.-C.; Li, T.-C. Energy expenditure in preterm infants during periods of environmental stress in the neonatal intensive care unit. Jpn. J. Nurs. Sci. JJNS 2014, 11, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Stevens, B.; Yamada, J.; Lee, G.Y.; Ohlsson, A. Sucrose for analgesia in newborn infants undergoing painful procedures. Cochrane Database Syst. Rev. 2013, 1. [Google Scholar] [CrossRef]
- Atkinson, L.; Jamieson, B.; Khoury, J.; Ludmer, J.; Gonzalez, A. Stress Physiology in Infancy and Early Childhood: Cortisol Flexibility, Attunement and Coordination. J. Neuroendocrinol. 2016, 28. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Stress, adaptation and disease. Allostasis and allostatic load. Ann. N. Y. Acad. Sci. 1998, 840, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, E.F.; Watterberg, K.L. Relative adrenal insufficiency in the preterm and term infant. J. Perinatol. Off. J. Calif. Perinat. Assoc. 2009, 29, S44–S49. [Google Scholar] [CrossRef] [PubMed]
- Heckmann, M.; Hartmann, M.F.; Kampschulte, B.; Gack, H.; Bödeker, R.-H.; Gortner, L.; Wudy, S.A. Cortisol production rates in preterm infants in relation to growth and illness: A noninvasive prospective study using gas chromatography-mass spectrometry. J. Clin. Endocrinol. Metab. 2005, 90, 5737–5742. [Google Scholar] [CrossRef] [PubMed]
- Ng, P.C. Effect of stress on the hypothalamic-pituitary-adrenal axis in the fetus and newborn. J. Pediatr. 2011, 158, e41–e43. [Google Scholar] [CrossRef] [PubMed]
- Boonen, E.; Berghe, G.V. Novel insights in the HPA-axis during critical illness. Acta Clin. Belg. 2014, 69, 397–406. [Google Scholar] [CrossRef]
- Peeters, B.; Boonen, E.; Langouche, L.; Van den Berghe, G. The HPA axis response to critical illness: New study results with diagnostic and therapeutic implications. Mol. Cell. Endocrinol. 2015, 408, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Stephens, M.A.C.; Wand, G. Stress and the HPA Axis. Alcohol Res. Curr. Rev. 2012, 34, 468–483. [Google Scholar]
- McEwen, B.S.; Gianaros, P.J. Central role of the brain in stress and adaptation: Links to socioeconomic status, health and disease. Ann. N. Y. Acad. Sci. 2010, 1186, 190–222. [Google Scholar] [CrossRef] [PubMed]
- Simmons, P.S.; Miles, J.M.; Gerich, J.E.; Haymond, M.W. Increased proteolysis. An effect of increases in plasma cortisol within the physiologic range. J. Clin. Investig. 1984, 73, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Haley, D.W.; Weinberg, J.; Grunau, R.E. Cortisol, contingency learning and memory in preterm and full-term infants. Psychoneuroendocrinology 2006, 31, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Quesada, A.A.; Tristão, R.M.; Pratesi, R.; Wolf, O.T. Hyper-responsiveness to acute stress, emotional problems and poorer memory in former preterm children. Stress Amst. Neth. 2014, 17, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Wadsby, M.; Nelson, N.; Ingemansson, F.; Samuelsson, S.; Leijon, I. Behaviour problems and cortisol levels in very-low-birth-weight children. Nord. J. Psychiatry 2014, 68, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Juruena, M.F. Early-life stress and HPA axis trigger recurrent adulthood depression. Epilepsy Behav. 2014, 38, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Raine, A. Prefrontal Structural and Functional Brain Imaging findings in Antisocial, Violent and Psychopathic Individuals: A Meta-Analysis. Psychiatry Res. 2009, 174, 81–88. [Google Scholar] [CrossRef] [PubMed]
- McKlveen, J.M.; Morano, R.L.; Fitzgerald, M.; Zoubovsky, S.; Cassella, S.N.; Scheimann, J.R.; Ghosal, S.; Mahbod, P.; Packard, B.A.; Myers, B.; et al. Chronic Stress Increases Prefrontal Inhibition: A Mechanism for Stress-Induced Prefrontal Dysfunction. Biol. Psychiatry 2016. [Google Scholar] [CrossRef] [PubMed]
- Reser, J.E. Chronic stress, cortical plasticity and neuroecology. Behav. Processes 2016, 129, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Boyer, K.; Johnston, C.; Walker, C.-D.; Filion, F.; Sherrard, A. Does sucrose analgesia promote physiologic stability in preterm neonates? Biol. Neonate 2004, 85, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Stang, H.J.; Snellman, L.W.; Condon, L.M.; Conroy, M.M.; Liebo, R.; Brodersen, L.; Gunnar, M.R. Beyond dorsal penile nerve block: A more humane circumcision. Pediatrics 1997, 100, E3. [Google Scholar] [CrossRef] [PubMed]
- Tryon, M.S.; Stanhope, K.L.; Epel, E.S.; Mason, A.E.; Brown, R.; Medici, V.; Havel, P.J.; Laugero, K.D. Excessive Sugar Consumption May Be a Difficult Habit to Break: A View From the Brain and Body. J. Clin. Endocrinol. Metab. 2015. [Google Scholar] [CrossRef] [PubMed]
- Banga, S.; Datta, V.; Rehan, H.S.; Bhakhri, B.K. Effect of Sucrose Analgesia, for Repeated Painful Procedures, on Short-term Neurobehavioral Outcome of Preterm Neonates: A Randomized Controlled Trial. J. Trop. Pediatr. 2016, 62, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, S.; Ranger, M.; Chau, C.M.Y.; Ellegood, J.; Lerch, J.P.; Holsti, L.; Goldowitz, D.; Grunau, R.E. Repeated exposure to sucrose for procedural pain in mouse pups leads to long-term widespread brain alterations. Pain 2017, 158, 1586–1598. [Google Scholar] [CrossRef] [PubMed]
- Ulrich-Lai, Y.M.; Ostrander, M.M.; Thomas, I.M.; Packard, B.A.; Furay, A.R.; Dolgas, C.M.; Van Hooren, D.C.; Figueiredo, H.F.; Mueller, N.K.; Choi, D.C.; et al. Daily limited access to sweetened drink attenuates hypothalamic-pituitary-adrenocortical axis stress responses. Endocrinology 2007, 148, 1823–1834. [Google Scholar] [CrossRef] [PubMed]
- Ulrich-Lai, Y.M.; Christiansen, A.M.; Ostrander, M.M.; Jones, A.A.; Jones, K.R.; Choi, D.C.; Krause, E.G.; Evanson, N.K.; Furay, A.R.; Davis, J.F.; et al. Pleasurable behaviors reduce stress via brain reward pathways. Proc. Natl. Acad. Sci. USA 2010, 107, 20529–20534. [Google Scholar] [CrossRef] [PubMed]
- Slater, R.; Cornelissen, L.; Fabrizi, L.; Patten, D.; Yoxen, J.; Worley, A.; Boyd, S.; Meek, J.; Fitzgerald, M. Oral sucrose as an analgesic drug for procedural pain in newborn infants: A randomised controlled trial. Lancet 2010, 376, 1225–1232. [Google Scholar] [CrossRef]
- Ulrich-Lai, Y.M.; Ostrander, M.M.; Herman, J.P. HPA axis dampening by limited sucrose intake: Reward frequency vs. caloric consumption. Physiol. Behav. 2011, 103, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Blass, E.M.; Ciaramitaro, V. A new look at some old mechanisms in human newborns: Taste and tactile determinants of state, affect and action. Monogr. Soc. Res. Child Dev. 1994, 59, 1–81. [Google Scholar] [CrossRef]
- Fitzgerald, M. What do we really know about newborn infant pain? Exp. Physiol. 2015, 100, 1451–1457. [Google Scholar] [CrossRef] [PubMed]
- Foo, H.; Mason, P. Ingestion analgesia occurs when a bad taste turns good. Behav. Neurosci. 2011, 125, 956–961. [Google Scholar] [CrossRef] [PubMed]
- Takamata, A.; Mack, G.W.; Gillen, C.M.; Nadel, E.R. Sodium appetite, thirst and body fluid regulation in humans during rehydration without sodium replacement. Am. J. Physiol. 1994, 266, R1493–R1502. [Google Scholar] [CrossRef] [PubMed]
- Tappy, L.; Lê, K.-A. Metabolic effects of fructose and the worldwide increase in obesity. Physiol. Rev. 2010, 90, 23–46. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Population | Parameter Measured | Effect | Sucrose Dose | Control | References |
|---|---|---|---|---|---|
| Premature Infant | ATP Utilization | Increased | 2 mL for neonates >2 kg 1.5 mL for neonates 1.5–2 kg 0.5 mL for neonates that were <1.5 kg | Placebo (Sterile Water) | [10] |
| Oxidative Stress | Increased | ||||
| Premature Infant | Salivary Cortisol | No Significant Difference | 0.1–0.3 mL per Painful Procedure | Placebo (Sterile Water) | [91] |
| Premature Infant | Plasma Cortisol | No Significant Difference | Pacifier Dipped in Sucrose | Water | [92] |
| Adult Human Females | Salivary Cortisol | Decreased | 3 Servings of Study Beverageper Day | Aspartame | [93] |
| Regional Brain Responses | Increased in Left Hippocampus | ||||
| Premature Infant | Neurodevelopmental Assessment | No Significant Difference | 0.5 mL per Painful Procedure | Placebo (Sterile Water) | [94] |
| Mouse Pups | CNS White Matter Regions | Decreased | 0.1–0.2 g Sucrose per kg Body Weight | Vehicle (Sterile Water) | [95] |
| CNS Gray Matter Regions | Decreased | ||||
| Adult Rat | Plasma ACTH | Decreased | Up to 4 mL Twice a Day | Saccharin and/or Water | [96,97] |
| Corticosterone | Decreased | ||||
| CRH mRNA | Decreased |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, J.B.C.; Boskovic, D.S.; Angeles, D.M. The Energy Costs of Prematurity and the Neonatal Intensive Care Unit (NICU) Experience. Antioxidants 2018, 7, 37. https://doi.org/10.3390/antiox7030037
Tan JBC, Boskovic DS, Angeles DM. The Energy Costs of Prematurity and the Neonatal Intensive Care Unit (NICU) Experience. Antioxidants. 2018; 7(3):37. https://doi.org/10.3390/antiox7030037
Chicago/Turabian StyleTan, John B. C., Danilo S. Boskovic, and Danilyn M. Angeles. 2018. "The Energy Costs of Prematurity and the Neonatal Intensive Care Unit (NICU) Experience" Antioxidants 7, no. 3: 37. https://doi.org/10.3390/antiox7030037
APA StyleTan, J. B. C., Boskovic, D. S., & Angeles, D. M. (2018). The Energy Costs of Prematurity and the Neonatal Intensive Care Unit (NICU) Experience. Antioxidants, 7(3), 37. https://doi.org/10.3390/antiox7030037