The Ageing Process Affects the Antioxidant Defences and the Poly (ADPribosyl)ation Activity in Cistus Incanus L. Leaves

 ,
,  ,
,  ,
, 
 ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Chlorophyll a Fluorescence Measurement
2.3. Isolation of Nuclei and Western Blotting Analysis
2.4. Poly (ADP-Ribose) Polymerase (PARP) Assay Activity
2.5. Determination of Total Soluble and Fat-Soluble Antioxidant Capacity
2.6. Antioxidant Enzyme Analysis
2.7. Statistical Analysis
3. Results
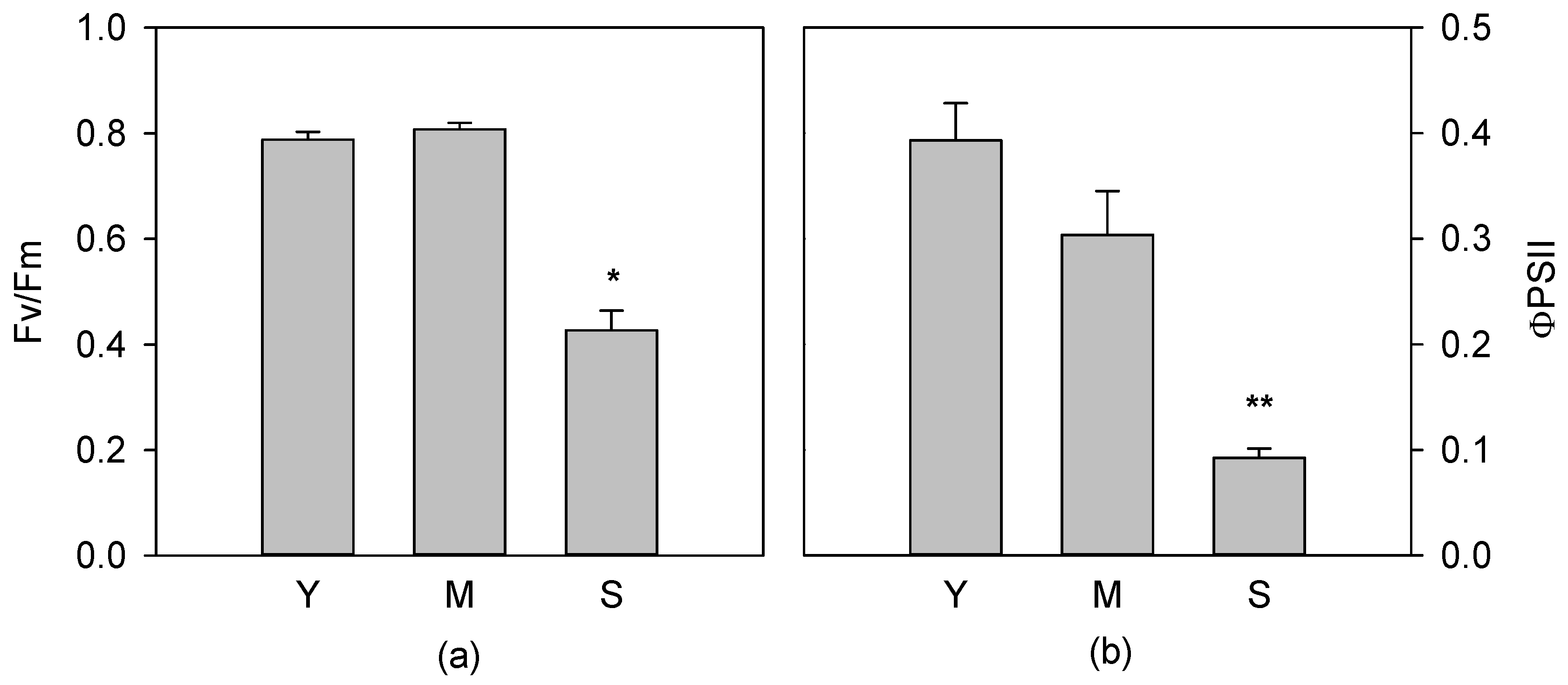
3.1. Photochemical Apparatus Efficiency
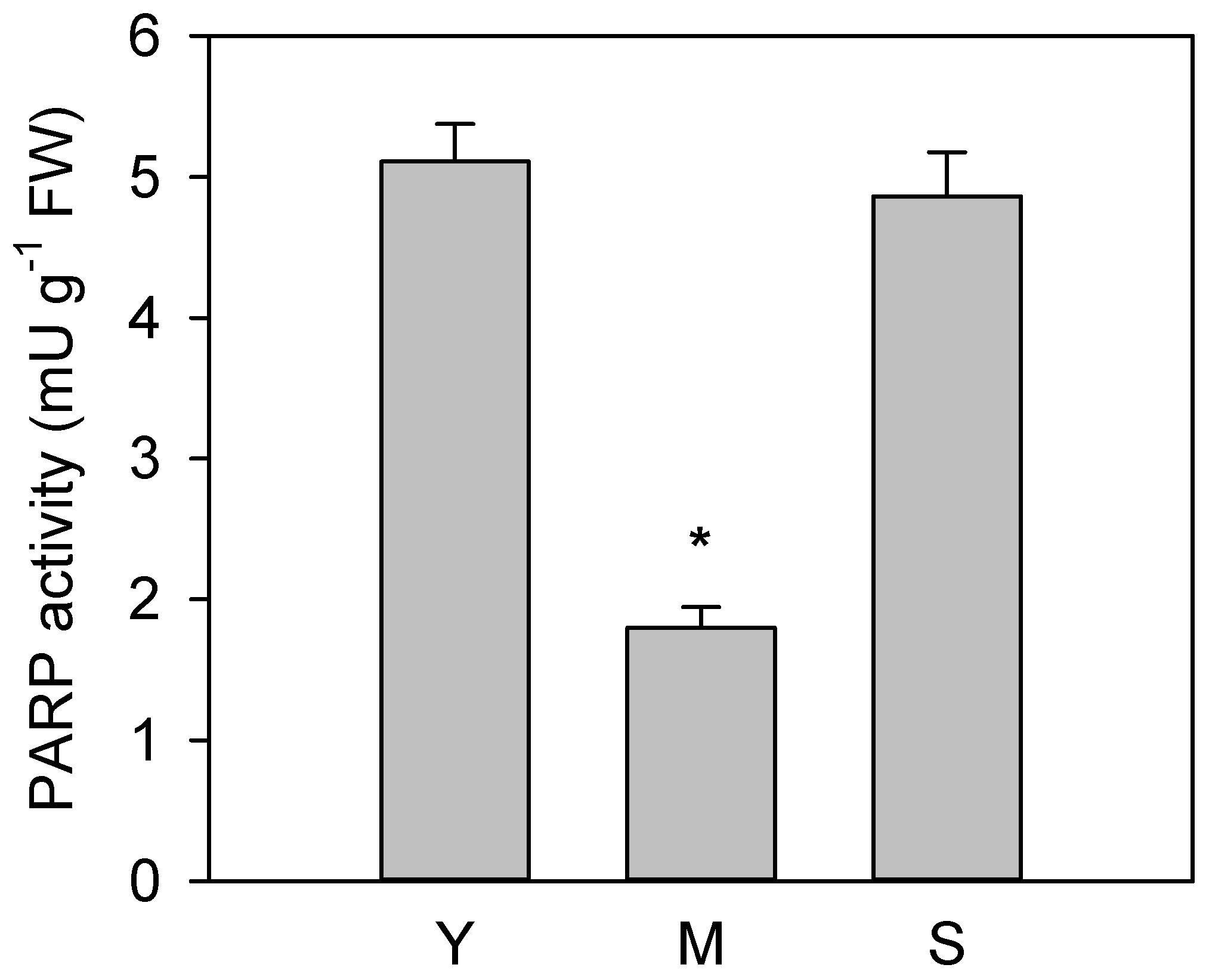
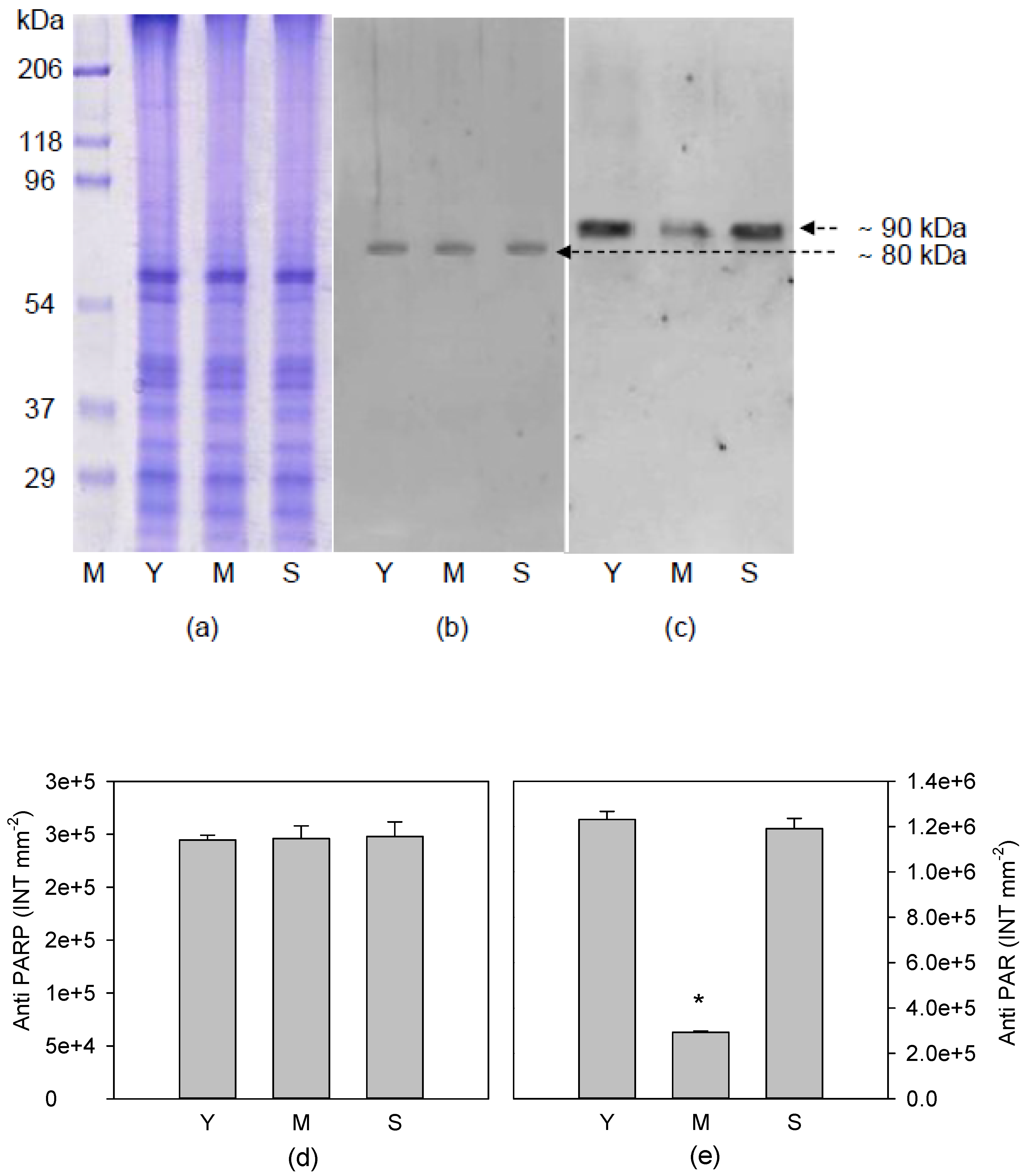
3.2. Poly(ADP)ribosylation Characterization
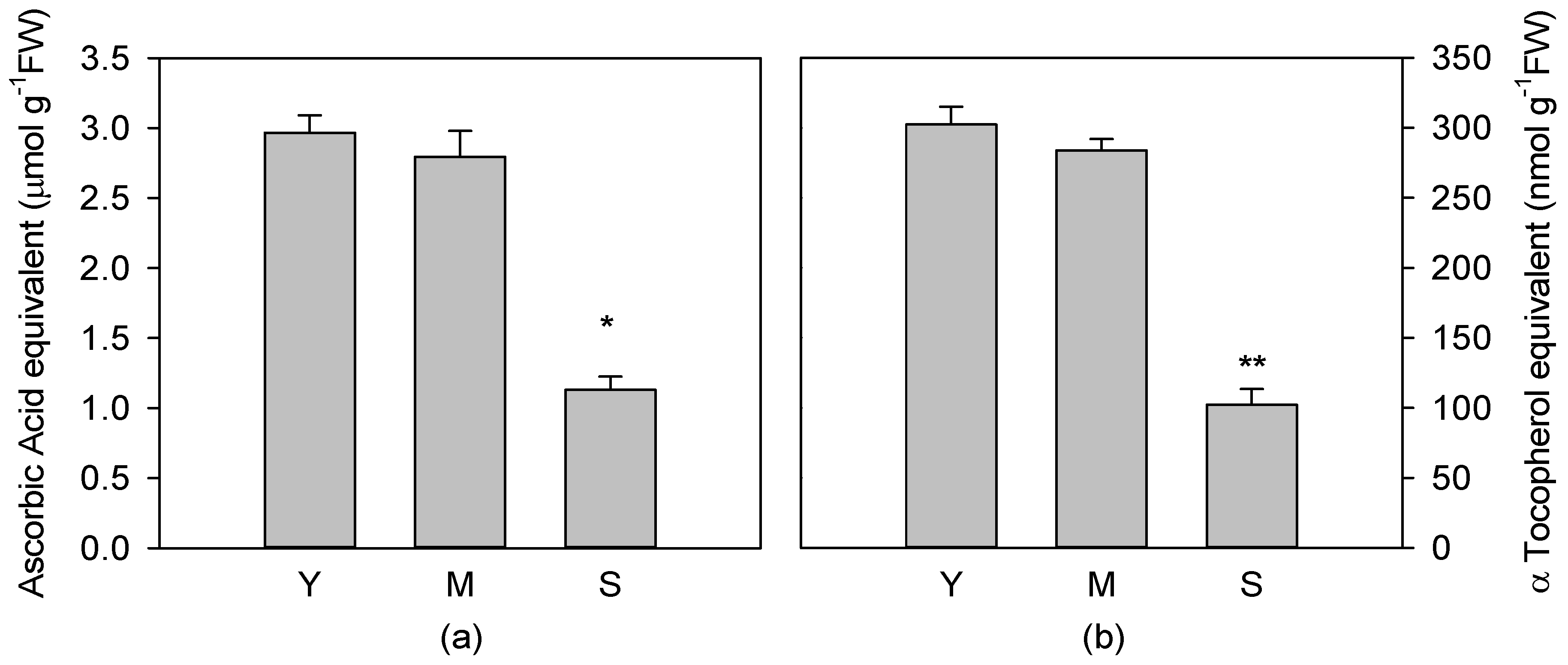
3.3. Total Soluble and Fat Soluble Antioxidant Capacity
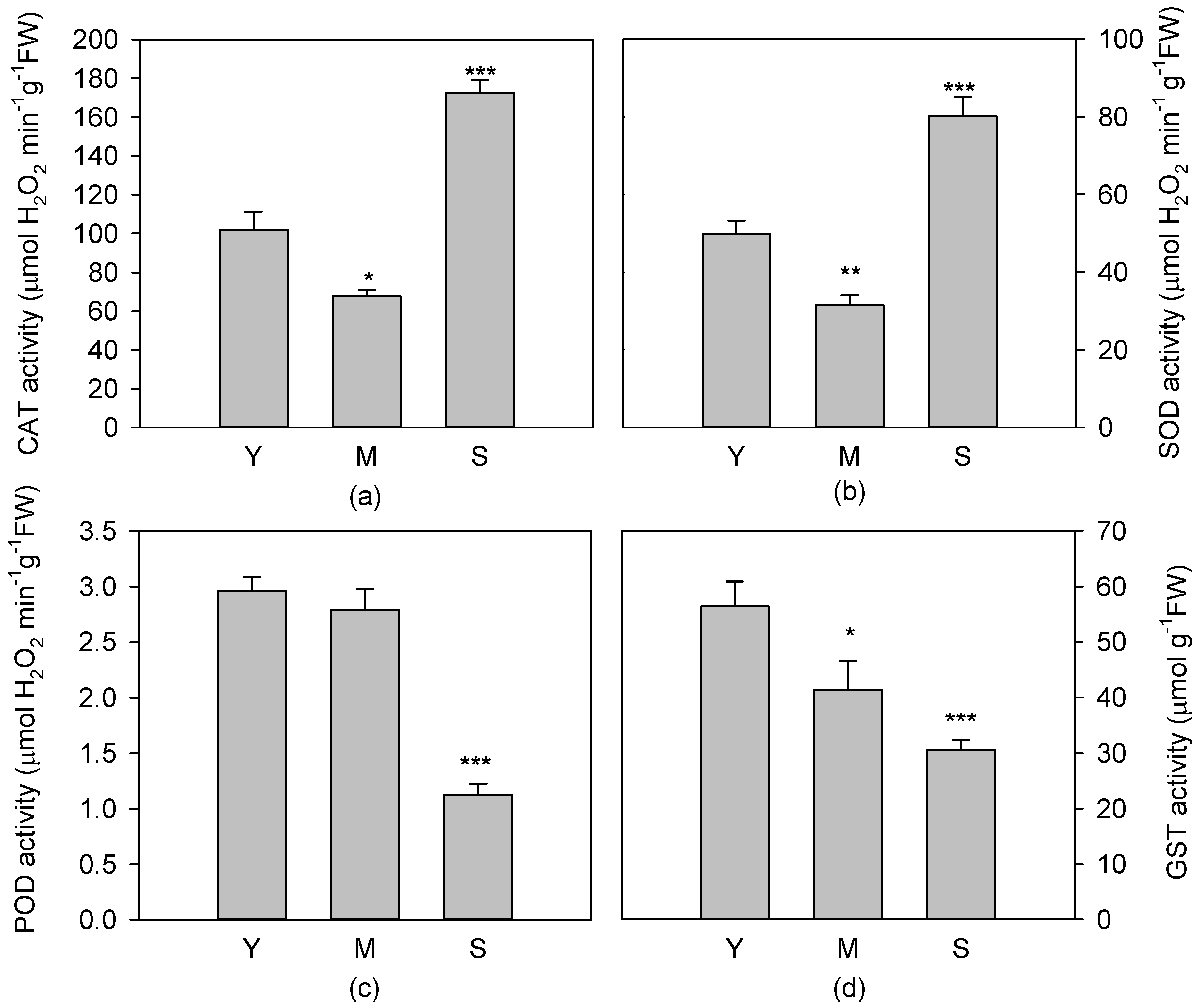
3.4. Activity of Antioxidant Enzymes
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Noodén, L.D.; Guiamet, J.J. Genetic control of senescence and aging in plants. In Handbook of the Biology of Aging, 4th ed.; Schneider, E.L., Rowe, J.W., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 94–118. [Google Scholar]
- Dangl, J.L.; Dietrich, R.A.; Thomas, H. Cell death and senescence. In Biochemistry and Molecular Biology of Plants; Buchanan, B., Gruissem, W., Jones, R., Eds.; ASPP Press: Rockville, MD, USA, 2000; pp. 1044–1100. [Google Scholar]
- Troen, B.R. The biology of aging. Mt. Sinai J. Med. 2003, 70, 3–22. [Google Scholar] [PubMed]
- Day, R.E.; Rogers, P.J.; Dawes, I.W.; Higgins, V.J. Molecular analysis of maltotriose transport and utilization by Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2002, 68, 5326–5335. [Google Scholar] [CrossRef] [PubMed]
- Woo, H.R.; Kim, H.J.; Nam, H.G.; Lim, P.O. Plant leaf senescence and death–regulation by multiple layers of control and implications for aging in general. J. Cell Sci. 2013, 126, 4823–4833. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Balazadeh, S.; Tohge, T.; Erban, A.; Giavalisco, P.; Kopka, J.; Mueller-Roeber, B.; Fernie, A.R.; Hoefgen, R. Comprehensive dissection of spatiotemporal metabolic shifts in primary, secondary, and lipid metabolism during developmental senescence in Arabidopsis. Plant Physiol. 2013, 162, 1290–1310. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, D.R.; Garcia Martinez, P.; Michel, X.; Narbonne, J.F.; O’Hara, S.; Ribera, D.; Winston, G.W. Oxyradical production as a pollution mediated mechanism of toxicity in the common mussel Mytilusedulis L. and other mollusks. Funct. Ecol. 1990, 4, 415–424. [Google Scholar] [CrossRef]
- Arena, C.; De Micco, V.; De Maio, A. Growth alteration and leaf biochemical responses in Phaseolus vulgaris exposed to different doses of ionising radiation. Plant Biol. 2014, 16, 194–202. [Google Scholar] [CrossRef]
- Svilar, D.; Goellner, E.M.; Almeida, K.H.; Sobol, R.W. Base excision repair and lesion-dependent sub pathways for repair of oxidative DNA damage. Antioxid. Redox Signal. 2011, 14, 2491–2507. [Google Scholar] [CrossRef]
- Barnes, D.E.; Lindahl, T. Repair and genetic consequences of endogenous DNA base damage in mammalian cells. Annu. Rev. Genet. 2004, 38, 445–476. [Google Scholar] [CrossRef]
- Kannan, K.; Jain, S.K. Oxidative stress and apoptosis. Pathophysiology 2000, 7, 153–163. [Google Scholar] [CrossRef]
- Choi, K.; Kim, J.; Kim, G.W.; Choi, C. Oxidative stress-induced necrotic cell death via mitochondira-dependent burst of reactive oxygen species. Curr. Neurovasc. Res. 2009, 6, 213–222. [Google Scholar] [CrossRef]
- Filomeni, G.; De Zio, D.; Cecconi, F. Oxidative stress and autophagy: The clash between damage and metabolic needs. Cell Death Differ. 2015, 22, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, D.V.; Wiley, C.D.; Velarde, M.C. Mitochondrial effectors of cellular senescence: Beyond the free radical theory of aging. Aging Cell 2015, 14, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Van der Oost, R.; Beyer, J.; Vermeulen, N.P.E. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Gruber, J.; Fong, S.; Chen, C.B.; Yoong, S.; Pastorin, G.; Schaffer, S.; Cheah, I.; Halliwell, B. Mitochondria-Targeted antioxidants and metabolic modulators as pharmacological interventions to slow ageing. Biotechnol. Adv. 2013, 31, 563–592. [Google Scholar] [CrossRef]
- Bodnar, A.G. Marine invertebrates as models for aging research. Exp. Gerontol. 2009, 44, 477–484. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef]
- Dizdaroglu, M.; Jaruga, P.; Birincioglu, M.; Rodriguez, H. Free radical-induced damage to DNA: Mechanisms and measurement. Free Radic. Biol. Med. 2002, 32, 1102–1115. [Google Scholar] [CrossRef]
- Bjelland, S.; Seeberg, E. Mutagenicity, toxicity and repair of DNA base damage induced by oxidation. Mutat. Res. 2003, 531, 37–80. [Google Scholar] [CrossRef]
- Maslov, A.Y.; Vijg, J. Genome instability, cancer and aging. Biochim. Biophysic. Acta 2009, 1790, 963–969. [Google Scholar] [CrossRef] [Green Version]
- David, S.S.; O’Shea, V.L.; Kundu, S. Base-excision repair of oxidative DNA damage. Nature 2007, 447, 941–950. [Google Scholar] [CrossRef] [Green Version]
- Berquist, B.R.; Wilson, D.M. Pathways for repairing and tolerating the spectrum of oxidative DNA lesions. Cancer Lett. 2012, 327, 61–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sancar, A.; Lindsey-Boltz, L.A.; Ünsal-Kaçmaz, K.; Linn, S. Molecular mechanisms of mammalian DNA repair and the DNA damage checkpoints. Annu. Rev. Biochem. 2004, 73, 39–85. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, V.; Amé, J.C.; Dollé, P.; Schultz, I.; Rinaldi, B.; Fraulob, V.; Ménissier-de Murcia, J.; de Murcia, G. Poly (ADP-ribose) polymerase-2 (PARP-2) is required for efficient base excision DNA repair in association with PARP-1 and XRCC1. J. Biol. Chem. 2002, 277, 23028–23036. [Google Scholar] [CrossRef] [PubMed]
- Bürkle, A. Poly(ADP-ribosyl)ation: A post translation protein modification linked with genome protection and mammalian longevity. Biogerontology 2000, 1, 41–46. [Google Scholar] [CrossRef]
- D’Amours, D.; Desnoyers, S.; D’Silva, I.; Poirier, G.G. Poly (ADP-ribosyl)ation reactions in the regulation of nuclear functions. Biochem. J. 1999, 342, 249–268. [Google Scholar] [CrossRef]
- Kim, M.Y.; Zhang, T.; Kraus, W.L. Poly (ADP-ribosyl)ation by PARP-1: ‘PAR-laying’ NAD+ into a nuclear signal. Genes Dev. 2005, 19, 1951–1967. [Google Scholar] [CrossRef]
- Amé, J.C.; Rolli, V.; Schreiber, V.; Niedergang, C.; Apiou, F.; Decker, P.; Muller, S.; Höger, T.; Ménissier-de Murcia, J.; de Murcia, G. PARP-2, A novel mammalian DNA damage-dependent poly (ADP-ribose) polymerase. J. Biol. Chem. 1999, 274, 17860–17868. [Google Scholar] [CrossRef]
- Shieh, W.M.; Amé, J.C.; Wilson, M.V.; Wang, Z.Q.; Koh, D.W.; Jacobson, M.K.; Jacobson, E.L. Poly (ADP-ribose) polymerase null mouse cells synthesize ADP-ribose polymers. J. Biol. Chem. 1998, 273, 30069–30072. [Google Scholar] [CrossRef]
- Hooten, N.; Fitzpatrick, M.; Kompaniez, K.; Jacob, K.D.; Moore, B.R.; Nagle, J.; Barnes, J.; Lohani, A.; Evans, M.K. Coordination of DNA repair by NEIL1 and PARP-1: A possible link to aging. Aging 2012, 4, 674–685. [Google Scholar] [CrossRef]
- Grube, K.; Bürkle, A. Poly (ADP-ribose) polymerase activity in mononuclear leukocytes of 13 mammalian species correlates with species-specific life span. Proc. Natl. Acad. Sci. USA 1992, 89, 11759–11763. [Google Scholar] [CrossRef]
- Kapahi, P.; Boulton, M.E.; Kirkwood, T.B. Positive correlation between mammalian life span and cellular resistance to stress. Free Radic. Biol. Med. 1999, 26, 495–500. [Google Scholar] [CrossRef]
- Beneke, S.; Alvarez-Gonzalez, R.; Bürkle, A. Comparative characterisation of poly (ADP-ribose) polymerase-1 from two mammalian species with different life span. Exp. Gerontol. 2000, 35, 989–1002. [Google Scholar] [CrossRef]
- Doucet-Chabeaud, G.; Godon, C.; Brutesco, C.; De Murcia, G.; Kazmaier, M. Ionising radiation induces the expression of PARP-1 and PARP-2 genes in Arabidopsis. Mol. Genet. Genomics 2001, 265, 954–963. [Google Scholar] [CrossRef]
- Babiychuk, E.; Cottrill, P.B.; Storozhenko, S.; Fuangthong, M.; Chen, Y.; O’Farrell, M.K.; Van Montagu, M.; Inze, D.; Kushnir, S. Higher plants possess two structurally different poly (ADP-ribose) polymerases. Plant J. 1998, 15, 635–645. [Google Scholar] [CrossRef]
- Briggs, A.G.; Bent, A.F. Poly (ADP-ribosyl) ation in plants. Trends Plant Sci. 2011, 16, 372–380. [Google Scholar] [CrossRef]
- Bianchi, A.R.; De Maio, A. Synthesis and degradation of poly (ADP-ribose) in plants. Front. Biosci. 2014, 19, 1436–1444. [Google Scholar] [CrossRef]
- Lamb, R.S.; Citarelli, M.; Teotia, S. Functions of the poly (ADP-ribose) polymerase superfamily in plants. Cell. Mol. Life Sci. 2012, 69, 175–189. [Google Scholar] [CrossRef]
- Chen, I.; Haehnel, U.; Altschmied, L.; Schubert, I.; Puchta, H. The transcriptional response of Arabidopsis to genotoxic stress—A high-density colony array study (HDCA). Plant J. 2003, 35, 771–786. [Google Scholar] [CrossRef]
- Arena, C.; Mistretta, C.; Di Natale, E.; FaraoneMennella, M.R.; Virzo De Santo, A.; De Maio, A. Characterization and role of poly(ADP-ribosyl)ation in the Mediterranean species Cistus incanus L. under different temperature conditions. Plant Physiol. Biochem. 2011, 49, 435–440. [Google Scholar] [CrossRef]
- Adams-Phillips, L.; Briggs, A.G.; Bent, A.F. Disruption of poly (ADP-ribosyl)ation mechanisms alters responses of Arabidopsis to biotic stress. Plant Physiol. 2010, 152, 267–280. [Google Scholar] [CrossRef]
- Arena, C.; De Micco, V.; De Maio, A.; Mistretta, C.; Aronne, G.; Vitale, L. Winter and summer leaves of Cistus incanus: Differences in leaf morphofunctional traits, photosynthetic energy partitioning, and poly(ADP-ribose) polymerase (PARP) activity. Botany 2013, 91, 805–813. [Google Scholar] [CrossRef]
- Hunt, L.; Holdsworth, M.J.; Gray, J.E. Nicotinamidase activity is important for germination. Plant J. 2007, 51, 341–351. [Google Scholar] [CrossRef]
- De Micco, V.; Aronne, G. Seasonal dimorphism in wood anatomy of the Mediterranean Cistus incanus L. subsp. incanus. Trees 2009, 23, 981–989. [Google Scholar] [CrossRef]
- De Micco, V.; Arena, C.; Vitale, L.; Aronne, G.; Virzo De Santo, A. Mediterranean Cistus incanus winter leaves under natural outdoor and warmer indoor conditions. Botany 2011, 89, 677–688. [Google Scholar] [CrossRef]
- Vitale, L.; Magliulo, V.; Arena, C. Morphological and physiological modifications of Cistus salvifolius L. winter leaves to rise of winter temperatures. Plant Biosyst. 2014, 148, 1093–1101. [Google Scholar] [CrossRef]
- Van Kooten, O.; Snell, J.F.H. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 90, 87–92. [Google Scholar] [CrossRef]
- De Maio, A.; Natale, E.; Rotondo, S.; Di Cosmo, A.; Faraone Mennella, M.R. Vault-Poly-ADP-ribose polymerase in the Octopus vulgaris brain: A regulatory factor of actin polymerization dynamic. Comp. Biochem. Phys. B Biochem. Mol. Biol. 2013, 166, 40–47. [Google Scholar] [CrossRef]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef]
- Wong, D.W.S.; Whitaker, J.R. Catalase. In Handbook of Food Enzymology; Whitaker, J.R., Voragen, A.G.J., Wong, D.W.S., Eds.; Marcel Dekker Inc.: New York, NY, USA, 2003; pp. 389–401. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalases and peroxidases. Methods Enzymol. 1955, 2, 764–817. [Google Scholar] [CrossRef]
- Verma, S.; Dubey, R.S. Lead toxicity induces lipid peroxidation and alters the activities of antioxidant enzymes in growing rice plants. Plant Sci. 2003, 164, 645–655. [Google Scholar] [CrossRef]
- Yuan, Z.W.; Jiang, T.J. Peroxidase. In Handbook of Food Enzymology; Whitaker, J.R., Voragen, A.G.J., Wong, D.W.S., Eds.; Marcel Dekker Inc.: New York, NY, USA, 2003; pp. 389–401. [Google Scholar] [CrossRef]
- Sun, M.; Zigman, S. An improved spectrophotometric assay for superoxide dismutase based on epinephrine autoxidation. Anal. Biochem. 1977, 90, 81–89. [Google Scholar] [CrossRef]
- Khopde, S.M.; Priyadarsini, K.I.; Mohan, H.; Gawandi, V.B.; Satav, J.G.; Yakhmi, J.V.; Banavaliker, M.M.; Biyani, M.K.; Mittal, J.P. Characterizing the antioxidant activity of amla (Phyllanthusemblica) extract. Curr. Sci. 2001, 81, 185–190. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases the first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar]
- Arena, C.; De Maio, A.; De Nicola, F.; Santorufo, L.; Vitale, L.; Maisto, G. Assessment of eco-physiological performance of Quercus ilex L. leaves in urban area by an integrated approach. Water Air Soil Pollut. 2014, 225, 1824. [Google Scholar] [CrossRef]
- Burkle, A. DNA repair and PARP in aging. Free Radic. Res. 2006, 40, 1295–1302. [Google Scholar] [CrossRef]
- Burkle, A.; Diefenbach, J.; Brabeck, C.; Beneke, S. Ageing and PARP. Pharmacol. Res. 2005, 52, 93–99. [Google Scholar] [CrossRef]
- Piskunova, T.S.; Yurova, M.N.; Ovsyannikov, A.I.; Semenchenko, A.V.; Zabezhinski, M.A.; Popovich, I.G.; Wang, Z.Q.; Anisimov, V.N. Deficiency in poly(ADP-ribose) polymerase-1 (PARP-1) accelerates aging and spontaneous carcinogenesis in mice. Curr. Gerontol. Geriatr. Res. 2008, 754190. [Google Scholar] [CrossRef]
- Ding, S.; Wang, L.; Yang, Z.; Lu, Q.; Wen, X.; Lu, C. Decreased glutathione reductase 2 leads to early leaf senescence in Arabidopsis. J. Integr. Plant Biol. 2016, 58, 29–47. [Google Scholar] [CrossRef]
- Xie, X.; He, Z.; Chen, N.; Tang, Z.; Wang, Q.; Cai, Y. The roles of environmental factors in regulation of oxidative stress in plant. Hindaw iBioMed. Res. Int. 2019, 19, 9732325. [Google Scholar] [CrossRef]
- Munne’-Bosch, S.; Alegre, L. The function of tocopherols and tocotrienols in plants. Crit. Rev. Plant Sci. 2002, 21, 31–57. [Google Scholar] [CrossRef]
- Krishnakumar, R.; Gamble, M.J.; Frizzell, K.M.; Berrocal, J.G.; Kininis, M.; Kraus, W.L. Reciprocal binding of PARP-1 and histone H1 at promoters specifies transcriptional outcomes. Science 2008, 319, 819–821. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Kunkowska, A.B.; Schippers, J.H.M. Role of reactive oxygen species during cell expansion in leaves. Plant Physiol. 2016, 172, 2098–2106. [Google Scholar] [CrossRef]
- Rodrıguez, A.A.; Grunberg, K.A.; Taleisnik, E.L. Reactive oxygen species in the elongation zone of maize leaves are necessary for leaf extension. Plant Physiol. 2002, 129, 1627–1632. [Google Scholar] [CrossRef]
- Gapper, C.; Dolan, L. Control of plant development by reactive oxygen species. Plant Physiol. 2006, 141, 341–345. [Google Scholar] [CrossRef]
- Kunert, K.J.; Ederer, M. Leaf aging and lipid peroxidation: The role of the antioxidants vitamin C and E. Physiol. Plant. 1985, 65, 85–88. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arena, C.; Vitale, L.; Bianchi, A.R.; Mistretta, C.; Vitale, E.; Parisi, C.; Guerriero, G.; Magliulo, V.; De Maio, A. The Ageing Process Affects the Antioxidant Defences and the Poly (ADPribosyl)ation Activity in Cistus Incanus L. Leaves. Antioxidants 2019, 8, 528. https://doi.org/10.3390/antiox8110528
Arena C, Vitale L, Bianchi AR, Mistretta C, Vitale E, Parisi C, Guerriero G, Magliulo V, De Maio A. The Ageing Process Affects the Antioxidant Defences and the Poly (ADPribosyl)ation Activity in Cistus Incanus L. Leaves. Antioxidants. 2019; 8(11):528. https://doi.org/10.3390/antiox8110528
Chicago/Turabian StyleArena, Carmen, Luca Vitale, Anna Rita Bianchi, Carmela Mistretta, Ermenegilda Vitale, Costantino Parisi, Giulia Guerriero, Vincenzo Magliulo, and Anna De Maio. 2019. "The Ageing Process Affects the Antioxidant Defences and the Poly (ADPribosyl)ation Activity in Cistus Incanus L. Leaves" Antioxidants 8, no. 11: 528. https://doi.org/10.3390/antiox8110528
APA StyleArena, C., Vitale, L., Bianchi, A. R., Mistretta, C., Vitale, E., Parisi, C., Guerriero, G., Magliulo, V., & De Maio, A. (2019). The Ageing Process Affects the Antioxidant Defences and the Poly (ADPribosyl)ation Activity in Cistus Incanus L. Leaves. Antioxidants, 8(11), 528. https://doi.org/10.3390/antiox8110528