Serratia marcescens BM1 Enhances Cadmium Stress Tolerance and Phytoremediation Potential of Soybean Through Modulation of Osmolytes, Leaf Gas Exchange, Antioxidant Machinery, and Stress-Responsive Genes Expression

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Investigation of Cadmium Tolerance of Serratia marcescens BM1 Strain
2.2. Inoculation and Growth of Soybean Plants
2.3. Morphological Parameters of Plant Root and Shoot
2.4. Measurement of Phosphorus, Nitrogen, and Cadmium Uptake
2.5. Measurements of Chlorophyll Content, Leaf Relative Water Content, and Gas-Exchange Attributes
2.6. Measurement of Total Soluble Sugars, Soluble Protein, Proline and Glycine Betaine Levels
2.7. Determination of Total Flavonoids and Phenols Contents
2.8. Estimation of Hydrogen Peroxide and Malondialdehyde Levels
2.9. Determination of Leaf Antioxidant Capacity
2.10. Estimation of Antioxidant Enzymes
2.11. Expression Analysis of Stress-Related Genes
2.12. Statistical Analysis
3. Results and Discussion
3.1. Cadmium Tolerance of Serratia marcescens BM1
3.2. Serratia marcescens BM1 Enhances Growth and Biomass of Soybean Plants Under Cadmium Stress
3.3. Serratia marcescens BM1 Modulates the Contents of Cadmium, Nitrogen and Phosphorous in Cadmium-Stressed Soybean Plants
3.4. Serratia marcescens BM1 Induces Leaf Gas Exchange Attributes, Leaf Relative Water Content and Biosynthesis of Chlorophyll, Sugars, Proteins, Osmolytes, Flavonoids, and Phenolics Under Cadmium Stress
3.5. Serratia marcescens BM1 Inoculation Reduces the Contents of H2O2 and MDA Under Cadmium Stress
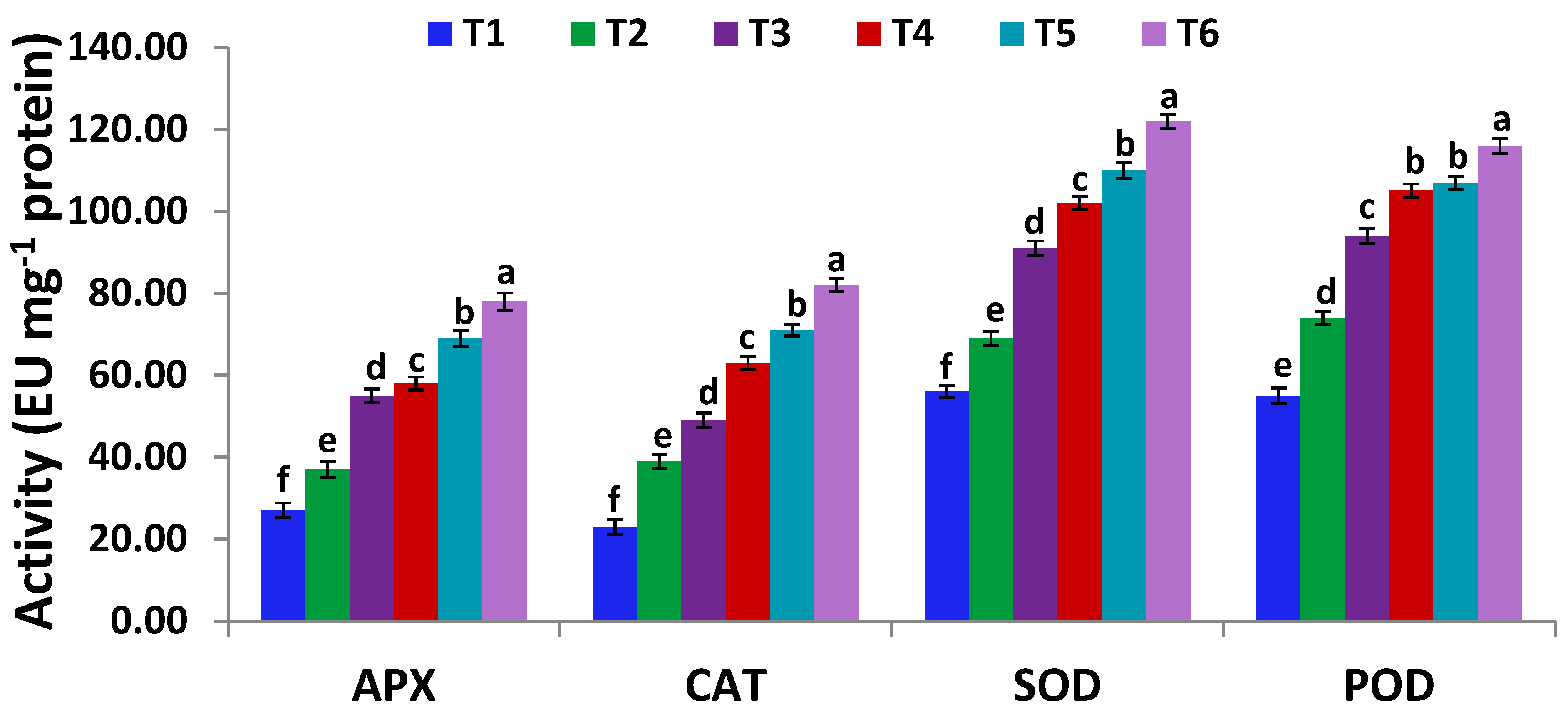
3.6. Serratia marcescens BM1 Inoculation Modulates Antioxidant Capacity and Antioxidative Enzyme Activities Under Cadmium Stress
3.7. Serratia marcescens BM1 Induces the Expression Levels of Stress-Related Genes Under Cadmium Stress
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wani, W.; Masoodi, K.Z.; Zaid, A.; Wani, S.H.; Shah, F.; Meena, V.S.; Wani, S.A.; Mosa, K.A. Engineering plants for heavy metal stress tolerance. Rend. Lincei 2018, 29, 709–723. [Google Scholar] [CrossRef]
- Abou-Kassem, D.E.; Mahrose, K.M.; Alagawany, M. The role of vitamin E or clay in growing Japanese quail fed diets polluted by cadmium at various levels. Animal 2016, 10, 508–519. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Rizwan, M.; Zaid, A.; Arif, M.S.; Yasmeen, T.; Hussain, A.; Shahid, M.R.; Bukhari, S.A.H.; Hussain, S.; Abbasi, G.H. 5-Aminolevulinic Acid-Induced Heavy Metal Stress Tolerance and Underlying Mechanisms in Plants. J. Plant Growth Regul. 2018, 37, 1423–1436. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Zia-ur-Rehman, M.; Maqbool, A. A critical review on the effects of zinc at toxic levels of cadmium in plants. Environ. Sci. Pollut. Res. 2019, 26, 6279–6289. [Google Scholar] [CrossRef]
- Per, T.S.; Khan, N.A.; Masood, A.; Fatma, M. Methyl jasmonate alleviates cadmium-induced photosynthetic damages through increased S-assimilation and glutathione production in mustard. Front. Plant Sci. 2016, 7, 1933. [Google Scholar] [CrossRef] [Green Version]
- Zaid, A.; Mohammad, F. Methyl Jasmonate and Nitrogen Interact to Alleviate Cadmium Stress in Mentha arvensis by Regulating Physio-Biochemical Damages and ROS Detoxification. J. Plant Growth Regul. 2018, 37, 1331–1348. [Google Scholar] [CrossRef]
- Elkelish, A.A.; Alhaithloul, H.A.S.; Qari, S.H.; Soliman, M.H.; Hasanuzzaman, M. Pretreatment with Trichoderma harzianum alleviates waterlogging-induced growth alterations in tomato seedlings by modulating physiological, biochemical, and molecular mechanisms. Environ. Exp. Bot. 2019, 171, 103946. [Google Scholar] [CrossRef]
- El-Esawi, M.; Arthaut, L.; Jourdan, N.; d’Harlingue, A.; Martino, C.; Ahmad, M. Blue-light induced biosynthesis of ROS contributes to the signaling mechanism of Arabidopsis cryptochrome. Sci. Rep. 2017, 7, 13875. [Google Scholar] [CrossRef]
- Consentino, L.; Lambert, S.; Martino, C.; Jourdan, N.; Bouchet, P.-E.; Witczak, J.; Castello, P.; El-Esawi, M.; Corbineau, F.; d’Harlingue, A.; et al. Blue-light dependent reactive oxygen species formation by Arabidopsis cryptochrome may define a novel evolutionarily conserved signalling mechanism. New Phytol. 2015, 206, 1450–1462. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Ali, H.M.; Alayafi, A.A.; Witczak, J.; Ahmad, M. Genetic variation and alleviation of salinity stress in barley. Molecules 2018, 23, 2488. [Google Scholar] [CrossRef] [Green Version]
- El-Esawi, M.A.; Alayafi, A.A. Overexpression of rice Rab7 gene improves drought and heat tolerance and increases grain yield in rice (Oryza sativa L.). Genes 2019, 10, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Esawi, M.A.; Alayafi, A.A. Overexpression of StDREB2 Transcription Factor Enhances Drought Stress Tolerance in Cotton (Gossypium barbadense L.). Genes 2019, 10, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soliman, M.; Alhaithloul, H.A.; Hakeem, K.R.; Alharbi, B.M.; El-Esawi, M.; Elkelish, A. Exogenous Nitric Oxide Mitigates Nickel-Induced Oxidative Damage in Eggplant by Upregulating Antioxidants, Osmolyte Metabolism, and Glyoxalase Systems. Plants 2019, 8, 562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Ahmad, M. Overexpression of AtWRKY30 Transcription Factor Enhances Heat and Drought Stress Tolerance in Wheat (Triticum aestivum L.). Genes 2019, 10, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Esawi, M.A.; Elkelish, A.; Elansary, H.O.; Ali, H.M.; Elshikh, M.; Witczak, J.; Ahmad, M. Genetic transformation and hairy root induction enhance the antioxidant potential of Lactuca serriola L. Oxid. Med. Cell. Longev. 2017, 2017, 5604746. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Qin, C.; Begum, N.; Maodong, Q.; Dong, X.X.; El-Esawi, M.; El-Sheikh, M.A.; Alatar, A.A.; Zhang, L. Nitrogen availability prevents oxidative effects of salinity on wheat growth and photosynthesis by up-regulating the antioxidants and osmolytes metabolism, and secondary metabolite accumulation. BMC Plant Biol. 2019, 19, 479. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Wang, Y.; Li, G.Z.; Hao, L. Salicylic acid-altering Arabidopsis plant response to cadmium exposure: Underlying mechanisms affecting antioxidation and photosynthesis-related processes. Ecotoxicol. Environ. Saf. 2019, 169, 645–653. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Hussain, A.; Ali, Q.; Shakoor, M.B.; Zia-ur-Rehman, M.; Farid, M.; Asma, M. Effect of zinc-lysine on growth, yield and cadmium uptake in wheat (Triticum aestivum L.) and health risk assessment. Chemosphere 2017, 187, 35–42. [Google Scholar] [CrossRef]
- Luo, H.; Li, H.; Zhang, X.; Fu, J. Antioxidant responses and gene expression in perennial ryegrass (Lolium perenne L.) under cadmium stress. Ecotoxicology 2011, 20, 770–778. [Google Scholar] [CrossRef]
- Khanna, K.; Jamwal, V.L.; Kohli, S.K.; Gandhi, S.G.; Ohri, P.; Bhardwaj, R.; Abd-Allah, E.F.; Hashem, A.; Ahmad, P. Plant growth promoting rhizobacteria induced Cd tolerance in Lycopersicon esculentum through altered antioxidative defense expression. Chemosphere 2019, 217, 463–474. [Google Scholar] [CrossRef]
- Mnasri, M.; Ghabriche, R.; Fourati, E.; Zaier, H.; Sabally, K.; Barrington, S.; Ghnaya, T. Cd and Ni transport and accumulation in the halophyte Sesuvium portulacastrum: Implication of organic acids in these processes. Front. Plant Sci. 2015, 6, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, A.J.; Nascimento, C.W.A.; Gouveia-Neto, A.S. Assessment of cadmium phytotoxicity alleviation by silicon using chlorophyll a fluorescence. Photosynthetica 2016, 55, 648–654. [Google Scholar] [CrossRef]
- Sakouhi, L.; Rahoui, S.; Gharsallah, C.; Munemasa, S.; El Ferjani, E.; Murata, Y.; Chaoui, A. Effects of calcium and EGTA on thiol homeostasis and defense-related enzymes in Cd-exposed chickpea roots. Acta Physiol. Plant. 2018, 40, 20. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; ur Rehman, M.Z.; Rinklebe, J.; Tsang, D.C.; Bashir, A.; Maqbool, A.; Tack, F.M.G.; Ok, Y.S. Cadmium phytoremediation potential of Brassica crop species: A review. Sci. Total Environ. 2018, 631, 1175–1191. [Google Scholar] [CrossRef]
- Ma, Y.; Rajkumar, M.; Vicente, J.A.F.; Freitas, H. Inoculation of Ni-resistant plant growth promoting bacterium Psychrobacter sp. strain SRS8 for the improvement of nickel phytoextraction by energy crops. Int. J. Phytoremed. 2010, 13, 126–139. [Google Scholar] [CrossRef]
- Shi, P.; Zhu, K.; Zhang, Y.; Chai, T. Growth and Cadmium Accumulation of Solanum nigrum L. Seedling were Enhanced by Heavy Metal-Tolerant Strains of Pseudomonas aeruginosa. Water Air Soil Pollut. 2016, 227, 459. [Google Scholar] [CrossRef]
- Mesa-Marín, J.; Del-Saz, N.F.; Rodríguez-Llorente, I.D.; Redondo-Gómez, S.; Pajuelo, E.; Ribas-Carbó, M.; Mateos-Naranjo, E. PGPR Reduce Root Respiration and Oxidative Stress Enhancing Spartina maritima Root Growth and Heavy Metal Rhizoaccumulation. Front. Plant Sci. 2018, 9, 1500. [Google Scholar] [CrossRef] [Green Version]
- Madhaiyan, M.; Poonguzhali, S.; Sa, T. Metal tolerating methylotrophic bacteria reduces nickel and cadmium toxicity and promotes plant growth of tomato (Lycopersicon esculentum L.). Chemosphere 2007, 69, 220–228. [Google Scholar] [CrossRef]
- Wei, T.; Lv, X.; Jia, H.; Hua, L.; Xu, H.; Zhou, R.; Zhao, J.; Ren, X.; Guo, J. Effects of salicylic acid, Fe (II) and plant growth-promoting bacteria on Cd accumulation and toxicity alleviation of Cd tolerant and sensitive tomato genotypes. J. Environ. Manag. 2018, 214, 164–171. [Google Scholar] [CrossRef]
- Sessitsch, A.; Kuffner, M.; Kidd, P.; Vangronsveld, J.; Wenzel, W.W.; Fallmann, K.; Puschenreiter, M. The role of plant-associated bacteria in the mobilization and phytoextraction of trace elements in contaminated soils. Soil Biol. Biochem. 2013, 60, 182–194. [Google Scholar] [CrossRef] [Green Version]
- Kao, P.H.; Huang, C.C.; Hseu, Z.Y. Response of microbial activities to heavy metals in a neutral loamy soil treated with biosolid. Chemosphere 2006, 64, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, G.; Farrag, K.; Soler-Rovira, P.; Ferrara, M.; Nigro, F.; Senesi, N. The effect of compost and Bacillus licheniformis on the phytoextraction of Cr, Cu, Pb and Zn by three brassicaceae species from contaminated soils in the Apulia region, Southern Italy. Geoderma 2012, 170, 322–330. [Google Scholar] [CrossRef]
- Malekzadeh, E.; Alikhani, H.A.; Savaghebi-Firoozabadi, G.R.; Zarei, M. Bioremediation of cadmium-contaminated soil through cultivation of maize inoculated with plant growth–promoting rhizobacteria. Bioremed. J. 2012, 16, 204–211. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Hashem, A.; Abd_Allah, E.F. Bacillus: A biological tool for crop improvement through bio-molecular changes in adverse environments. Front. Physiol. 2017, 8, 667. [Google Scholar] [CrossRef] [PubMed]
- Gupta, C.; Dubey, R.; Maheshwari, D. Plant growth enhancement and suppression of Macrophomina phaseolina causing charcoal rot of peanut by fluorescent Pseudomonas. Biol. Fertil. Soils 2002, 35, 399–405. [Google Scholar]
- Arkhipova, T.N.; Veselov, S.U.; Melentiev, A.I.; Martynenko, E.V.; Kudoyarova, G.R. Ability of bacterium Bacillus subtilis to produce cytokinins and to influence the growth and endogenous hormone content of lettuce plants. Plant Soil 2005, 272, 201–209. [Google Scholar] [CrossRef]
- Gururani, M.A.; Upadhyaya, C.P.; Baskar, V.; Venkatesh, J.; Nookaraju, A.; Park, S.W. Plant growth-promoting rhizobacteria enhance abiotic stress tolerance in Solanum tuberosum through inducing changes in the expression of ROS-scavenging enzymes and improved photosynthetic performance. J. Plant Growth Regul. 2013, 32, 245–258. [Google Scholar] [CrossRef]
- Chandra, P.; Tripathi, P.; Chandra, A. Isolation and molecular characterization of plant growth-promoting Bacillus spp. and their impact on sugarcane (Saccharum spp. hybrids) growth and tolerance towards drought stress. Acta Physiol. Plant. 2018, 40, 199. [Google Scholar] [CrossRef]
- Cristani, M.; Naccari, C.; Nostro, A.; Pizzimenti, A.; Trombetta, D.; Pizzimenti, F. Possible use of Serratia marcescens in toxic metal biosorption (removal). Environ. Sci. Pollut. Res. 2012, 19, 161–168. [Google Scholar] [CrossRef]
- Khan, A.R.; Park, G.S.; Asaf, S.; Hong, S.J.; Jung, B.K.; Shin, J.H. Complete genome analysis of Serratia marcescens RSC-14: A plant growth-promoting bacterium that alleviates cadmium stress in host plants. PLoS ONE 2017, 12, e0171534. [Google Scholar] [CrossRef] [Green Version]
- Queiroz, P.S.; Ruas, F.A.D.; Barboza, N.R.; de Castro Borges, W.; Guerra-Sá, R. Alterations in the proteomic composition of Serratia marcescens in response to manganese (II). BMC Biotechnol. 2018, 18, 83. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Wirth, S.; Jabborova, D.; Räsänen, L.A.; Liao, H. Coordination between Bradyrhizobium and Pseudomonas alleviates salt stress in soybean through altering root system architecture. J. Plant Interact. 2017, 12, 100–107. [Google Scholar] [CrossRef] [Green Version]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alamri, S.A.; Ali, H.M.; Alayafi, A.A. Bacillus firmus (SW5) augments salt tolerance in soybean (Glycine max L.) by modulating root system architecture, antioxidant defense systems and stress-responsive genes expression. Plant Physiol. Biochem. 2018, 132, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Elkelish, A.A.; Alnusaire, T.S.; Soliman, M.H. Calcium availability regulates antioxidant system, physio-biochemical activities and alleviates salinity stress mediated oxidative damage in soybean seedlings. J. Appl. Bot. Food Qual. 2019, 92, 258–266. [Google Scholar]
- Abd El-Azeem, S.A.M. Studies on Plant Growth Promoting Rhizosphere Microorganisms. Ph.D. Thesis, Faculty of Agriculture Suez Canal University, Ismailia, Egypt, 2006. [Google Scholar]
- Abd El-Azeem, S.A.M.; Mehana, T.A.; Shabayek, A.A. Some plant-growth-promoting traits of rhizobacteria isolated from Suez Canal region, Egypt. Afr. Crop Sci. Conf. Proc. 2007, 8, 1517–1525. [Google Scholar]
- Bremner, J.M. Total Nitrogen. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties (Methods of Soil); American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1965; pp. 1149–1178. [Google Scholar]
- Murphy, J.; Riley, J. A modified single solution method for the determination of phosphate in natural waters. Anal. Chem. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzym. 1987, 148, 350–382. [Google Scholar]
- Holá, D.; Benešová, M.; Honnerová, J.; Hnilička, F.; Rothová, O.; Kočová, M.; Hniličková, H. The evaluation of photosynthetic parameters in maize inbred lines subjected to water deficiency: Can these parameters be used for the prediction of performance of hybrid progeny? Photosynthetica 2010, 48, 545–558. [Google Scholar] [CrossRef]
- Dey, P.M. Oligosaccharides. In Methods in Plant Biochemistry, Carbohydrates; Dey, P.M., Ed.; Academic Press: London, UK, 1990; Volume 2, pp. 189–218. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bates, L.; Waldren, P.P.; Teare, J.D. Rapid determination of free proline of water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Grieve, C.; Grattan, S. Rapid assay for determination of water-soluble quaternary ammonium compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Zieslin, N.; Ben-Zaken, R. Peroxidase activity and presence of phenolic substances in peduncles of rose flowers. Plant Physiol. Biochem. 1993, 31, 333–339. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Ueno, H.; Yamakura, S.; Arastoo, R.S.; Oshima, T.; Kokubo, K. Systematic evaluation and mechanistic investigation of antioxidant activity of fullerenols using β carotene bleaching assay. Nanomaterials 2014, 7, 802596. [Google Scholar]
- Pyrzynska, K.; Pękal, A. Application of free radical diphenylpicrylhydrazyl (DPPH) to estimate the antioxidant capacity of food samples. Anal. Methods 2013, 5, 4288–4295. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Aebi, H. Catalase in vitro. Methods Enzym. 1984, 105, 121–126. [Google Scholar]
- Kono, Y. Generation of superoxide radical during autoxidation of hydroxylamine and an assay for superoxide dismutase. Arch. Biochem. Biophys. 1978, 186, 189–195. [Google Scholar] [CrossRef]
- Putter, J.; Becker, R. Methods of Enzymatic Analysis; Academic Press: New York, NY, USA, 1974; p. 685. [Google Scholar]
- Zhou, Y.; Huang, J.L.; Zhang, X.L.; Zhu, L.M.; Wang, X.F.; Guo, N.; Zhao, J.M.; Xing, H. Overexpression of Chalcone Isomerase (CHI) Increases Resistance Against Phytophthora sojae in Soybean. J. Plant Biol. 2018, 61, 309–319. [Google Scholar] [CrossRef]
- Wei, W.; Huang, J.; Hao, Y.J.; Zou, H.F.; Wang, H.W.; Zhao, J.Y.; Liu, X.Y.; Zhang, W.K.; Ma, B.; Zhang, J.S.; et al. Soybean GmPHD-type transcription regulators improve stress tolerance in transgenic Arabidopsis plants. PLoS ONE 2009, 4, e7209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaishnav, A.; Kumari, S.; Jain, S.; Varma, A.; Tuteja, N.; Choudhary, D.K. PGPR-mediated expression of salt tolerance gene in soybean through volatiles under sodium nitroprusside. J. Basic Microbiol. 2016, 56, 1274–1288. [Google Scholar] [CrossRef] [PubMed]
- Sirhindi, G.; Mir, M.A.; Abd-Allah, E.F.; Ahmad, P.; Gucel, S. Jasmonic Acid Modulates the Physio-Biochemical Attributes, Antioxidant Enzyme Activity, and Gene Expression in Glycine max under Nickel Toxicity. Front. Plant Sci. 2016, 7, 591. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Elansary, H.O.; El-Shanhorey, N.A.; Abdel-Hamid, A.M.E.; Ali, H.M.; Elshikh, M.S. Salicylic Acid-Regulated Antioxidant Mechanisms and Gene Expression Enhance Rosemary Performance under Saline Conditions. Front. Physiol. 2017, 8, 716. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, H.J.; Pak, J.H.; Cho, H.S.; Choi, H.K.; Jung, H.W.; Lee, D.H.; Chung, Y.-S. Overexpression of AtSZF2 from Arabidopsis showed enhanced tolerance to salt stress in soybean. Plant Breed. Biotechnol. 2017, 5, 1–15. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Selvakumar, G.; Mohan, M.; Kundu, S.; Gupta, A.D.; Joshi, P.; Nazim, S.; Gupta, H.S. Cold tolerance and plant growth promotion potential of Serratia marcescens strain SRM (MTCC 8708) isolated from flowers of summer squash (Cucurbita pepo). Lett. Appl. Microbiol. 2008, 46, 171–175. [Google Scholar] [CrossRef]
- Devi, K.A.; Pandey, P.; Sharma, G.D. Plant Growth-Promoting Endophyte Serratia marcescens AL2-16 Enhances the Growth of Achyranthes aspera L., a Medicinal Plant. HAYATI J. Biosci. 2016, 23, 173–180. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alzahrani, S.M.; Ali, H.M.; Alayafi, A.A.; Ahmad, M. Serratia liquefaciens KM4 Improves Salt Stress Tolerance in Maize by Regulating Redox Potential, Ion Homeostasis, Leaf Gas Exchange and Stress-Related Gene Expression. Int. J. Mol. Sci. 2018, 19, 3310. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P.; Jha, P.N. The multifarious PGPR Serratia marcescens CDP-13 augments induced systemic resistance and enhanced salinity tolerance of wheat (Triticum aestivum L.). PLoS ONE 2016, 11, e0155026. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.R.; Ullah, I.; Khan, A.L.; Park, G.S.; Waqas, M.; Hong, S.J.; Jung, B.K.; Kwak, Y.; Lee, I.J.; Shin, J.H. Improvement in phytoremediation potential of Solanum nigrum under cadmium contamination through endophytic-assisted Serratia sp. RSC-14 inoculation. Environ. Sci. Pollut. Res. 2015, 22, 14032–14042. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, U.; Chakraborty, B.N.; Chakraborty, A.P. Influence of Serratia marcescens TRS-1 on growth promotion and induction of resistance in Camellia sinensis against Fomes lamaoensis. J. Plant Interact. 2010, 5, 261–272. [Google Scholar] [CrossRef]
- Wan, Y.; Luo, S.; Chen, J.; Xiao, X.; Chen, L.; Zeng, G.; Liu, C.; He, Y. Effect of endophyte-infection on growth parameters and Cd-induced phytotoxicity of Cd-hyperaccumulator Solanum nigrum L. Chemosphere 2012, 89, 743–750. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Alayafi, A.A. Azospirillum lipoferum FK1 confers improved salt tolerance in chickpea (Cicer arietinum L.) by modulating osmolytes, antioxidant machinery and stress-related genes expression. Environ. Exp. Bot. 2019, 159, 55–56. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CdCl2 (µM) | BM1 | Root Length (cm) | Root Fresh Weight (g Plant−1) | Root Dry Weight (g Plant−1) | Shoot Length (cm) | Shoot Fresh Weight (g Plant−1) | Shoot Dry Weight (g Plant−1) | Root DW/Shoot DW |
|---|---|---|---|---|---|---|---|---|
| 0 | −BM1 (T1) | 15.6 ± 1.11b | 1.37 ± 0.07b | 0.15 ± 0.03b | 25.2 ± 1.4b | 1.83 ± 0.15b | 0.37 ± 0.14b | 0.41 ± 0.05b |
| +BM1 (T2) | 17.3 ± 1.25a | 1.51 ± 0.08a | 0.18 ± 0.06a | 26.9 ± 1.6a | 1.98 ± 0.12a | 0.42 ± 0.12a | 0.43 ± 0.04a | |
| 150 | −BM1 (T3) | 11.3 ± 1.11d | 0.98 ± 0.06d | 0.10 ± 0.05d | 17.4± 1.3d | 1.24 ± 0.11d | 0.25 ± 0.15d | 0.40 ± 0.05b |
| +BM1 (T4) | 12.9 ± 1.21c | 1.18 ± 0.09c | 0.12 ± 0.04c | 20.6 ± 1.5c | 1.41 ± 0.12c | 0.29 ± 0.11c | 0.42 ± 0.03a | |
| 300 | −BM1 (T5) | 8.2 ± 1.07f | 0.61 ± 0.08f | 0.06 ± 0.05f | 12.4 ± 1.6f | 0.77 ± 0.15f | 0.16 ± 0.13f | 0.37 ± 0.05d |
| +BM1 (T6) | 9.6 ± 1.10e | 0.77 ± 0.06e | 0.07 ± 0.04e | 15.8 ± 1.8e | 0.92 ± 0.12e | 0.18 ± 0.11e | 0.39 ± 0.03c |
| CdCl2 (µM) | BM1 | Cd Content (mg g−1 DW) | N Content (mg g−1 DW) | P Content (mg g−1 DW) | Pn (μmol m2 s−1) | E (mmol m2 s−1) | gs (mol m2 s−1) |
|---|---|---|---|---|---|---|---|
| 0 | −BM1 (T1) | 0.01 ± 0.01e | 0.88 ± 0.12b | 0.40 ± 0.08b | 17.03 ±1.24b | 1.84 ± 0.07b | 0.11 ± 0.03b |
| +BM1 (T2) | 0.01 ± 0.01e | 0.93 ± 0.11a | 0.44 ± 0.11a | 18.92 ±1.15a | 2.03 ± 0.05a | 0.13 ± 0.02a | |
| 150 | −BM1 (T3) | 0.19 ± 0.09c | 0.72 ± 0.09d | 0.29 ± 0.09d | 11.12 ±1.21d | 1.49 ± 0.04d | 0.06 ± 0.03d |
| +BM1 (T4) | 0.15 ± 0.05d | 0.81 ± 0.07c | 0.36 ± 0.10c | 13.25 ±1.23c | 1.58 ± 0.07c | 0.08 ± 0.02c | |
| 300 | −BM1 (T5) | 0.2 8± 0.11a | 0.58 ± 0.08f | 0.22 ± 0.11e | 8.91 ± 1.05f | 1.33 ± 0.07f | 0.03 ± 0.01f |
| +BM1 (T6) | 0.23 ± 0.10b | 0.65 ± 0.07e | 0.30 ± 0.12d | 10.33 ±1.07e | 1.42 ± 0.08e | 0.05 ± 0.03e |
| CdCl2 (µM) | BM1 | Chlorophyll (mg g−1 FW) | RWC (%) | Sugars (µg g−1 FW) | Proteins (mg g−1 FW) | Proline (µg g−1 FW) | GB (µmol g−1 FW) |
|---|---|---|---|---|---|---|---|
| 0 | −BM1 (T1) | 2.02 ± 0.08b | 91 ± 0.53b | 2.41 ± 0.14b | 1.11 ± 0.14b | 20.5 ± 0.29f | 2.36 ± 0.04f |
| +BM1 (T2) | 2.14 ± 0.07a | 93 ± 0.48a | 2.63 ± 0.16a | 1.23 ± 0.13a | 35.1 ± 0.24e | 3.22 ± 0.07e | |
| 150 | −BM1 (T3) | 1.59 ± 0.09d | 55 ± 0.51d | 2.17 ± 0.14d | 0.88 ± 0.10d | 55.2 ± 0.41d | 4.17 ± 0.06d |
| +BM1 (T4) | 1.68 ± 0.08c | 68 ± 0.46c | 2.29 ± 0.15c | 1.05 ± 0.14c | 77.3 ± 0.40c | 5.13 ± 0.05c | |
| 300 | −BM1 (T5) | 1.41 ± 0.09f | 42 ± 0.61e | 1.93 ± 0.17e | 0.67 ± 0.11f | 81.7 ± 0.52b | 6.06 ± 0.07b |
| +BM1 (T6) | 1.49 ± 0.07e | 54 ± 0.45d | 2.12 ± 0.13d | 0.79 ± 0.14e | 97.4 ± 0.55a | 7.88 ± 0.06a |
| CdCl2 (µM) | BM1 | Total Flavonoids | Total Phenolics | H2O2 (µmol g−1 FW) | MDA (µmol g−1 FW) | DPPH (IC50, μg mL−1) | β-Carotene Linoleic Acid (IC50, μg mL−1) |
|---|---|---|---|---|---|---|---|
| 0 | −BM1 (T1) | 7.21 ± 0.28b | 17.8 ± 0.24b | 0.27 ± 0.05d | 20 ± 1.71e | 0.53 ± 0.03a | 0.51 ± 0.05a |
| +BM1 (T2) | 9.11 ± 0.30a | 19.1 ± 0.23a | 0.18 ± 0.06e | 16 ± 1.66f | 0.48 ± 0.05b | 0.47 ± 0.04b | |
| 150 | −BM1 (T3) | 5.05 ± 0.22d | 13.6 ± 0.22d | 0.72 ± 0.04b | 51 ± 2.27b | 0.46 ± 0.04c | 0.44 ± 0.05c |
| +BM1 (T4) | 5.59 ± 0.27c | 14.9 ± 0.24c | 0.61 ± 0.07c | 42 ± 2.24d | 0.42 ± 0.03d | 0.41 ± 0.06d | |
| 300 | −BM1 (T5) | 2.9 ± 0.23f | 10.5 ± 0.31e | 0.85 ± 0.04a | 61 ± 2.43a | 0.38 ± 0.05e | 0.37 ± 0.05e |
| +BM1 (T6) | 4.1 ± 0.26e | 13.1 ± 0.29d | 0.60 ± 0.05c | 47 ± 2.11c | 0.35 ± 0.04f | 0.34 ± 0.05f |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Esawi, M.A.; Elkelish, A.; Soliman, M.; Elansary, H.O.; Zaid, A.; Wani, S.H. Serratia marcescens BM1 Enhances Cadmium Stress Tolerance and Phytoremediation Potential of Soybean Through Modulation of Osmolytes, Leaf Gas Exchange, Antioxidant Machinery, and Stress-Responsive Genes Expression. Antioxidants 2020, 9, 43. https://doi.org/10.3390/antiox9010043
El-Esawi MA, Elkelish A, Soliman M, Elansary HO, Zaid A, Wani SH. Serratia marcescens BM1 Enhances Cadmium Stress Tolerance and Phytoremediation Potential of Soybean Through Modulation of Osmolytes, Leaf Gas Exchange, Antioxidant Machinery, and Stress-Responsive Genes Expression. Antioxidants. 2020; 9(1):43. https://doi.org/10.3390/antiox9010043
Chicago/Turabian StyleEl-Esawi, Mohamed A., Amr Elkelish, Mona Soliman, Hosam O. Elansary, Abbu Zaid, and Shabir H. Wani. 2020. "Serratia marcescens BM1 Enhances Cadmium Stress Tolerance and Phytoremediation Potential of Soybean Through Modulation of Osmolytes, Leaf Gas Exchange, Antioxidant Machinery, and Stress-Responsive Genes Expression" Antioxidants 9, no. 1: 43. https://doi.org/10.3390/antiox9010043
APA StyleEl-Esawi, M. A., Elkelish, A., Soliman, M., Elansary, H. O., Zaid, A., & Wani, S. H. (2020). Serratia marcescens BM1 Enhances Cadmium Stress Tolerance and Phytoremediation Potential of Soybean Through Modulation of Osmolytes, Leaf Gas Exchange, Antioxidant Machinery, and Stress-Responsive Genes Expression. Antioxidants, 9(1), 43. https://doi.org/10.3390/antiox9010043