Induction of Antioxidant Protein HO-1 Through Nrf2-ARE Signaling Due to Pteryxin in Peucedanum Japonicum Thunb in RAW264.7 Macrophage Cells

Abstract
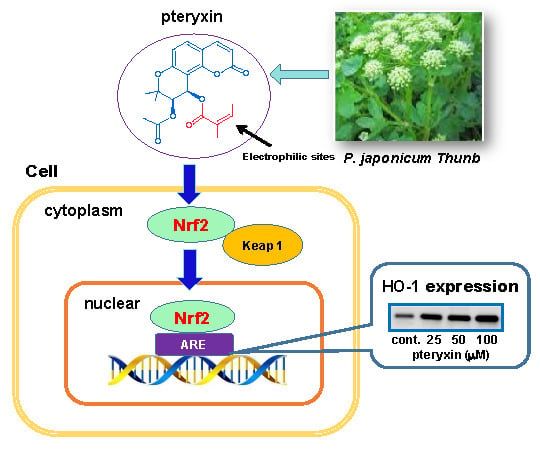
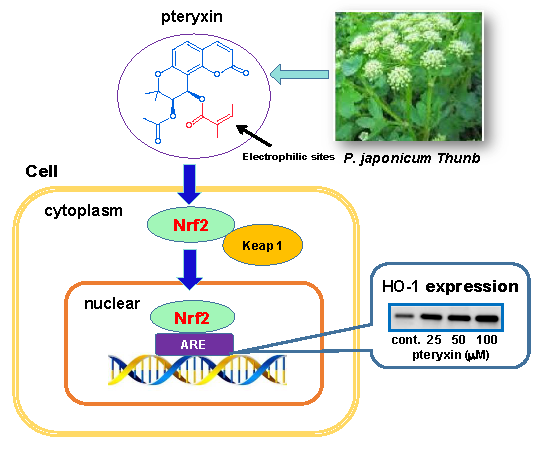
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Isolation of Pteryxin
2.3. Analysis of Pteryxin
2.4. Cell Culture
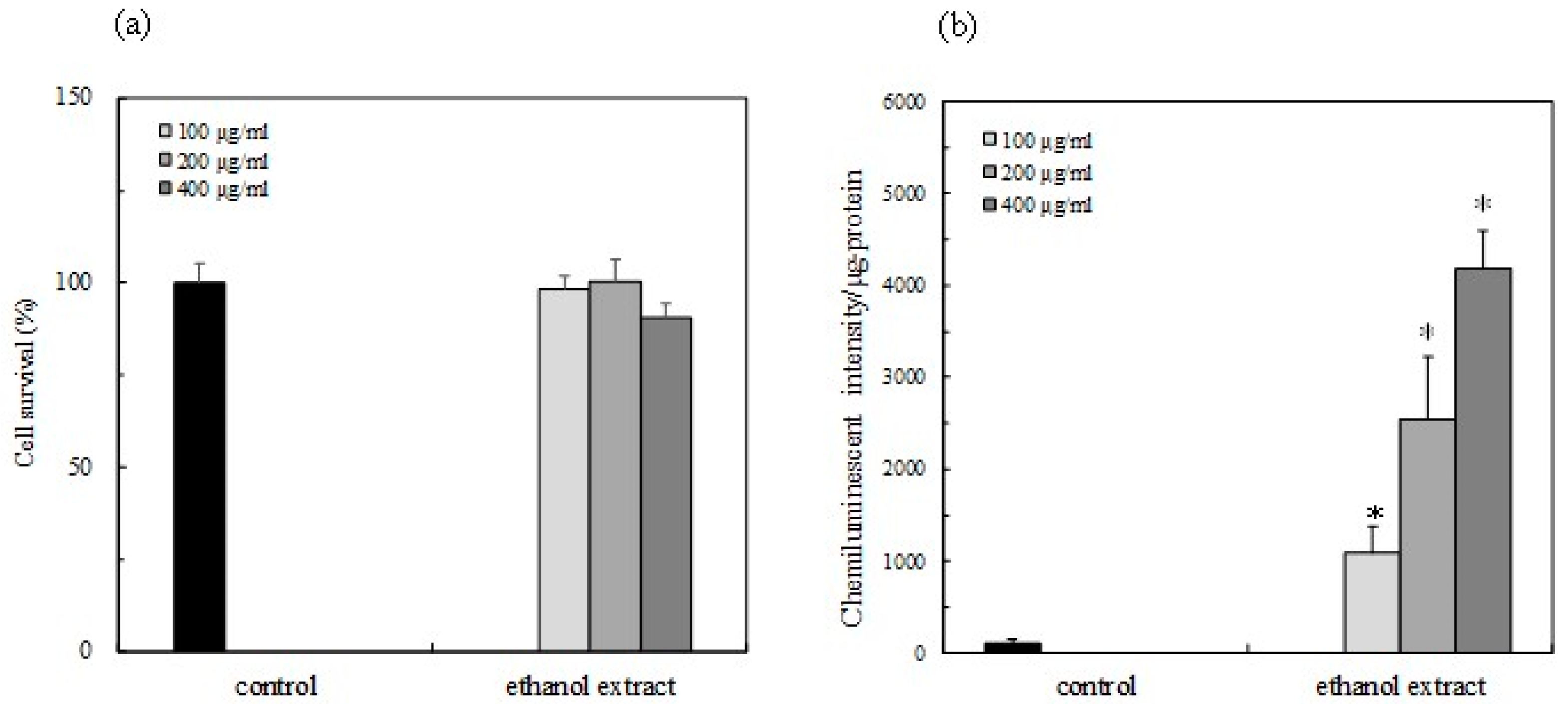
2.5. Cell Survaival
2.6. Activity of Nrf2-ARE Signaling
2.7. Nuclear Extraction and Determination of Nrf2
2.8. Protein Expression by Western Blot Analysis
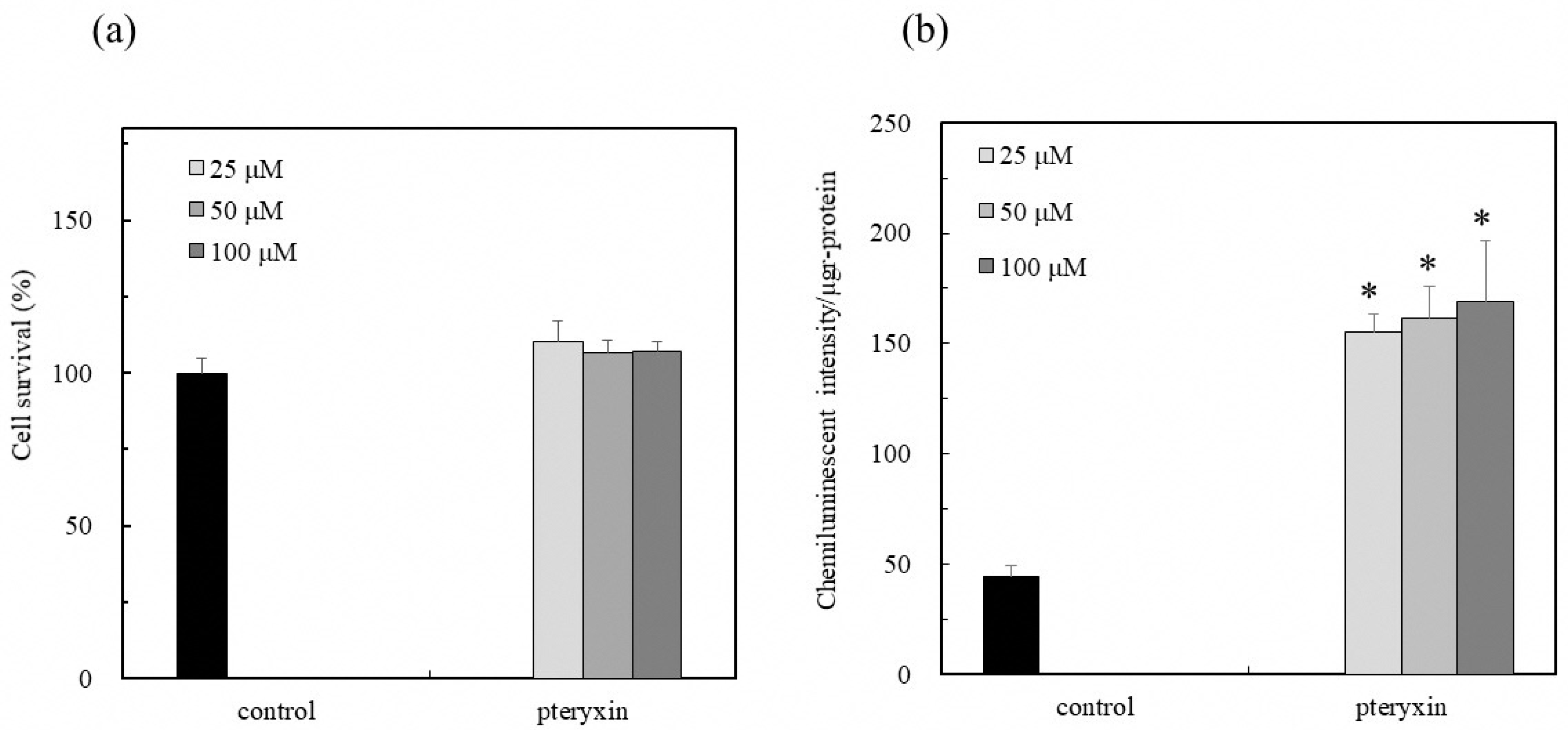
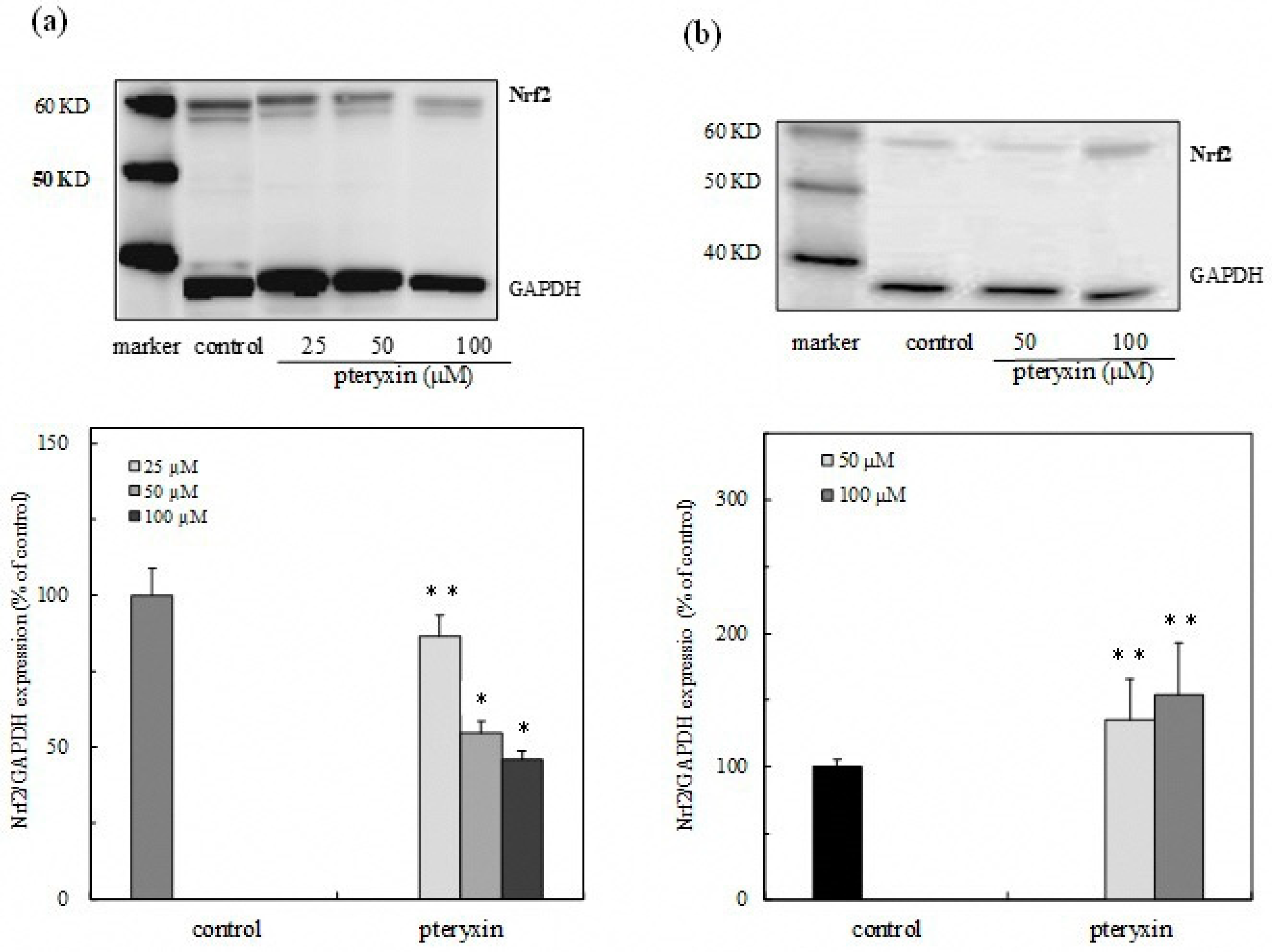
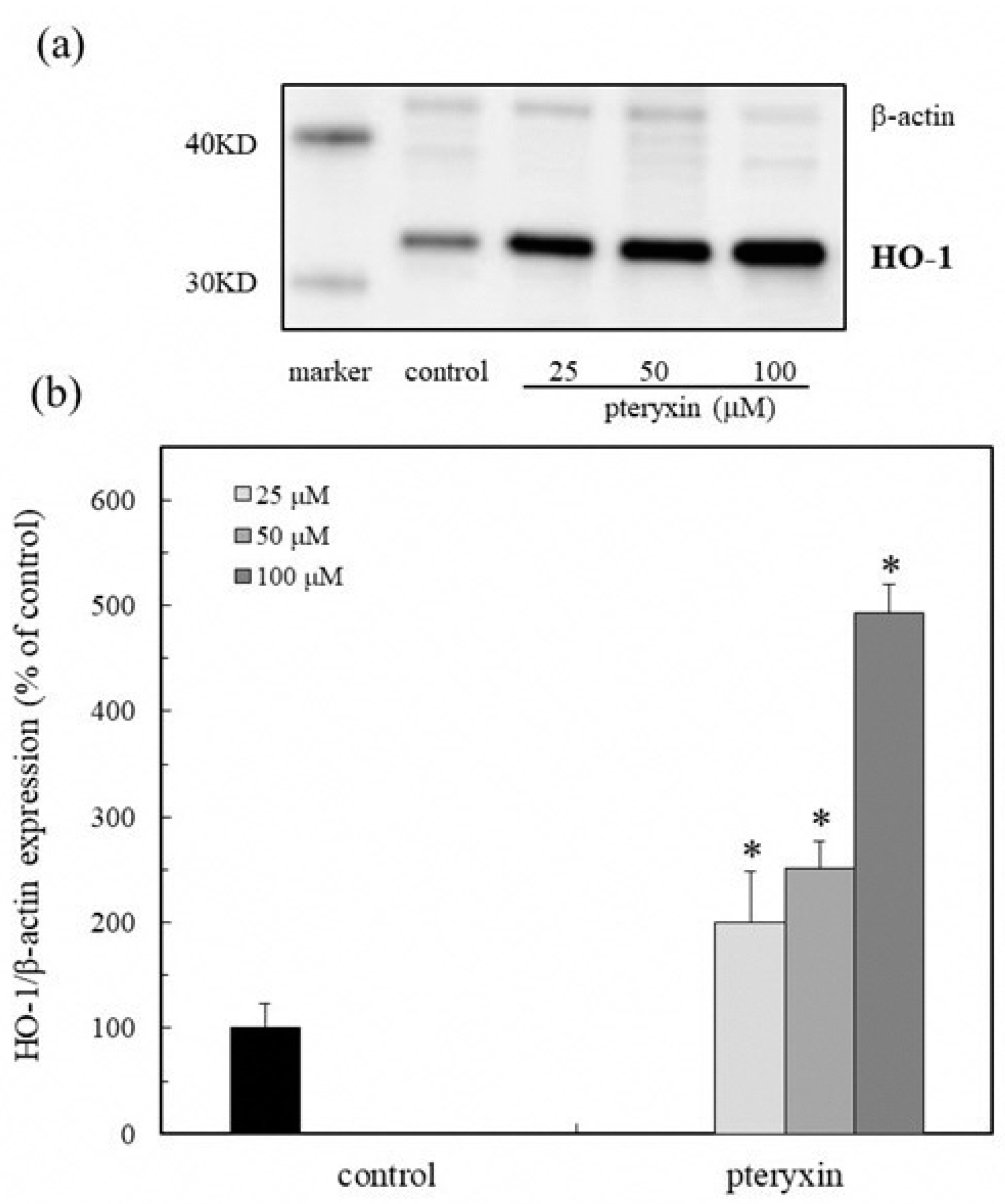
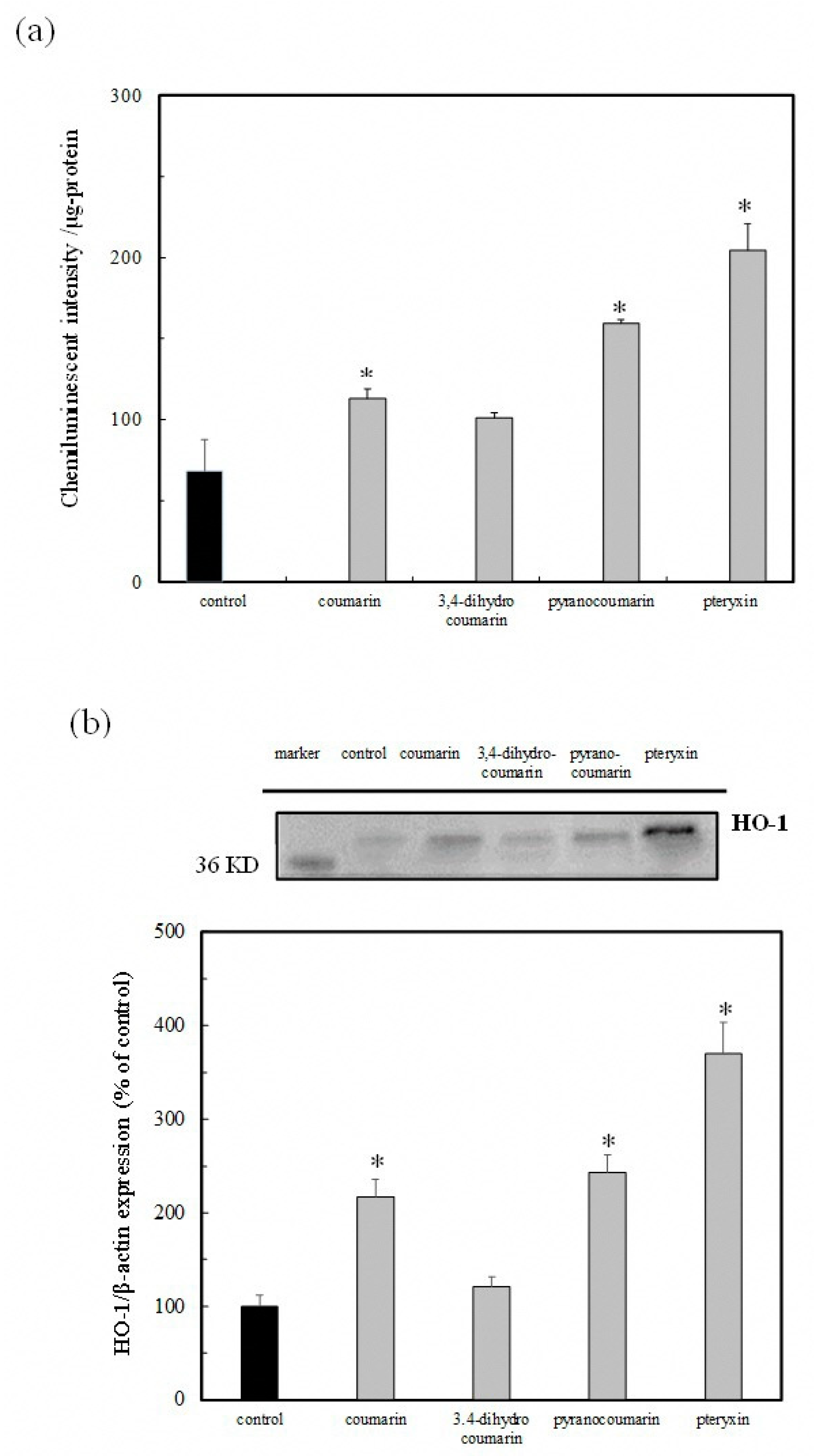
3. Results
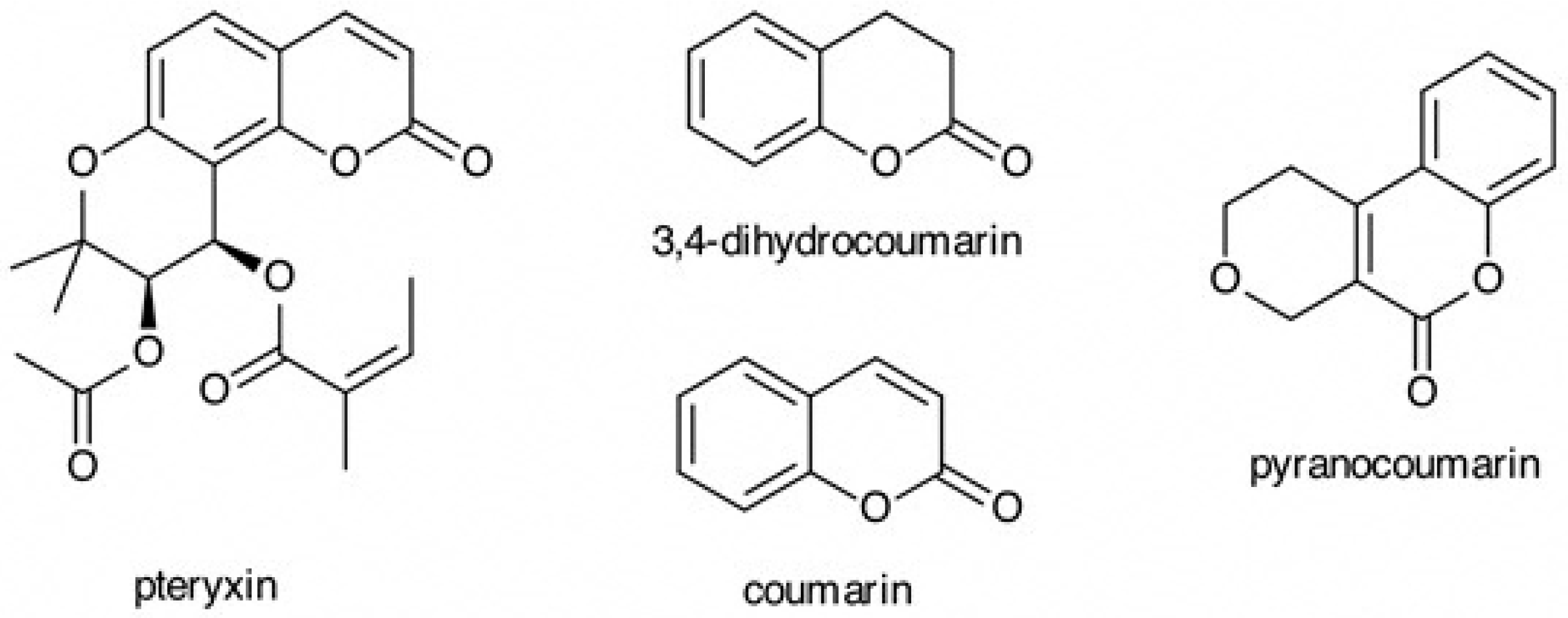
3.1. Determination of Pteryxin
3.2. Nrf2-ARE Signaling
3.3. Nrf2 Expression in Cytoplasm and Nuclei
3.4. HO-1 Expression
3.5. Nrf2-ARE Signaling Activity and HO-1 Protein Expression by Coumarin Derivatives
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sarkhail, P. Traditional uses, phytochemistry and pharmacological properties of the genus. J. Ethnopharmacol. 2014, 156, 235–270. [Google Scholar] [CrossRef] [PubMed]
- Hisamoto, M.; Kikuzaki, H.; Ohigashi, H.; Nakatani, N. Antioxidant compounds from the leaves of Peucedanum japonicum Thunb. J. Agric. Food Chem. 2003, 51, 5255–5261. [Google Scholar] [CrossRef] [PubMed]
- Hisamoto, M.; Kikuzaki, H.; Ohigashi, H.; Nakatani, N. Constituents of the leaves of Peucedanum japonicum Thunb. and their biological activity. J. Agric. Food Chem. 2004, 52, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Nukitrangsan, N.; Okabe, T.; Toda, T.; Inafuku, M.; Iwasaki, H.; Oku, H. Anti-obesity activity of Peucedanum japonicum Thunb extract in obese diabetic animal model C57BL/6JHam Slc-ob/ob Mice. Int. J. Life Sci. Med. Res. 2012, 2, 28–34. [Google Scholar] [CrossRef]
- Nugara, R.N.; Inafuku, M.; Iwasaki, H.; Oku, H. Partially purified Peucedanum japonicum Thunb extracts exert anti-obesity effects in vitro. Nutrition 2014, 30, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Nugara, R.N.; Inafuku, M.; Takara, K.; Iwasaki, H.; Oku, H. Pteryxin: A coumarin in Peucedanum japonicum Thunb leaves exerts antiobesity activity through modulation of adipogenic gene network. Nutrition 2014, 30, 1177–1184. [Google Scholar] [CrossRef]
- Okabe, T.; Toda, T.; Nukitrangsan, N.; Inafuku, M.; Iwasaki, H.; Oku, H. Peucedanum japonicum Thunb inhibits high-fat diet induced obesity in mice. Phytother. Res. 2011, 25, 870–877. [Google Scholar] [CrossRef]
- Choi, R.Y.; Nam, S.J.; Ham, J.R.; Lee, H.I.; Yee, S.T.; Kang, K.Y.; Lee, M.K. Anti-adipogenic and anti-diabetic effects of cis-3, 4-diisovalerylkhellactone isolated from Peucedanum japonicum Thunb leaves in vitro. Bioorganic Med. Chem. Lett. 2016, 26, 4655–4660. [Google Scholar] [CrossRef] [PubMed]
- Taira, N.; Nugara, R.N.; Inafuku, M.; Takara, K.; Ogi, T.; Ichiba, Y.; Iwasaki, H.; Okabe, T.; Oku, H.J. In vivo and in vitro anti-obesity activities of dihydropyranocoumarins derivatives from Peucedanum japonicum Thunb. J. Funct. Foods. 2017, 29, 19–28. [Google Scholar] [CrossRef]
- Kaspar, J.W.; Niture, S.K.; Jaiswal, A.K. Nrf2:INrf2 (Keap1) signaling in oxidative stress. Free Radic. Biol. Med. 2009, 47, 1304–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, H.-A.; Lee, I.-K. The Role of Nrf2: Adipocyte differentiation, obesity, and insulin resistance. Oxid. Med. Cell. Longev. 2013, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Kong, A.N. Dietary chemopreventive compounds and ARE/EpRE signaling. Free Radic. Biol. Med. 2004, 36, 1505–1516. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.-J.; Kundu, J.K.; Na, H.-K. Nrf2 as a master redox switch in turning on the cellular signaling involved in the induction of cytoprotective genes by some chemopreventive phytochemicals. Planta Med. 2008, 74, 1526–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taira, J.; Sonamoto, M.; Uehara, M. Dual biological functions of a cytoprotective effect and apoptosis induction by bioavailable marine carotenoid fucoxanthinol through modulation of the Nrf2 activation in RAW264.7 macrophage cells. Mar. Drugs 2017, 15, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taira, J.; Miyazato, H.; Ueda, K. Marine peroxy sesquiterpenoids induce apoptosis by modulation of Nrf2-ARE signaling in HCT116 colon cancer cells. Mar. Drugs 2018, 16, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taira, J.; Nanbu, H.; Ueda, K. Nitric oxide-scavenging compounds in Agrimonia pilosa Ledeb on LPS-induced RAW264.7 macrophages. Food Chem. 2009, 115, 1221–1227. [Google Scholar] [CrossRef]
- Yagishita, Y.; Uruno, A.; Fukutomi, T.; Sugiyama, F.; Takahashi, S.; Yamamoto, M. Nrf2 improves leptin and insulin resistance provoked by hypothalamic oxidative stress. Cell Rep. 2017, 18, 2030–2044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayama, N.; Iwamoto, N.; Sumi, D.; Shinkai, Y.; Tanaka-Kagawa, T.; Jinno, H.; Kumagai, Y. Peroxiredoxin 6 is a molecular target for 1, 2-naphthoquinone, an atmospheric electrophile in human pulmonary epithelial A549 cells. J. Toxicol. Sci. 2011, 36, 817–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taira, J.; Ogi, T. Induction of Antioxidant Protein HO-1 Through Nrf2-ARE Signaling Due to Pteryxin in Peucedanum Japonicum Thunb in RAW264.7 Macrophage Cells. Antioxidants 2019, 8, 621. https://doi.org/10.3390/antiox8120621
Taira J, Ogi T. Induction of Antioxidant Protein HO-1 Through Nrf2-ARE Signaling Due to Pteryxin in Peucedanum Japonicum Thunb in RAW264.7 Macrophage Cells. Antioxidants. 2019; 8(12):621. https://doi.org/10.3390/antiox8120621
Chicago/Turabian StyleTaira, Junsei, and Takayuki Ogi. 2019. "Induction of Antioxidant Protein HO-1 Through Nrf2-ARE Signaling Due to Pteryxin in Peucedanum Japonicum Thunb in RAW264.7 Macrophage Cells" Antioxidants 8, no. 12: 621. https://doi.org/10.3390/antiox8120621
APA StyleTaira, J., & Ogi, T. (2019). Induction of Antioxidant Protein HO-1 Through Nrf2-ARE Signaling Due to Pteryxin in Peucedanum Japonicum Thunb in RAW264.7 Macrophage Cells. Antioxidants, 8(12), 621. https://doi.org/10.3390/antiox8120621