Timing of Treatment with the Flavonoid 7,8-DHF Critically Impacts on Its Effects on Learning and Memory in the Ts65Dn Mouse




Abstract
:1. Introduction
2. Methods
2.1. Colony
2.2. Experimental Protocol
2.2.1. Experiment 1
2.2.2. Experiment 2
2.3. Behavioral Testing
2.4. DCX Immunohistochemistry
2.5. Western Blotting
2.6. Statistical Analysis
3. Results
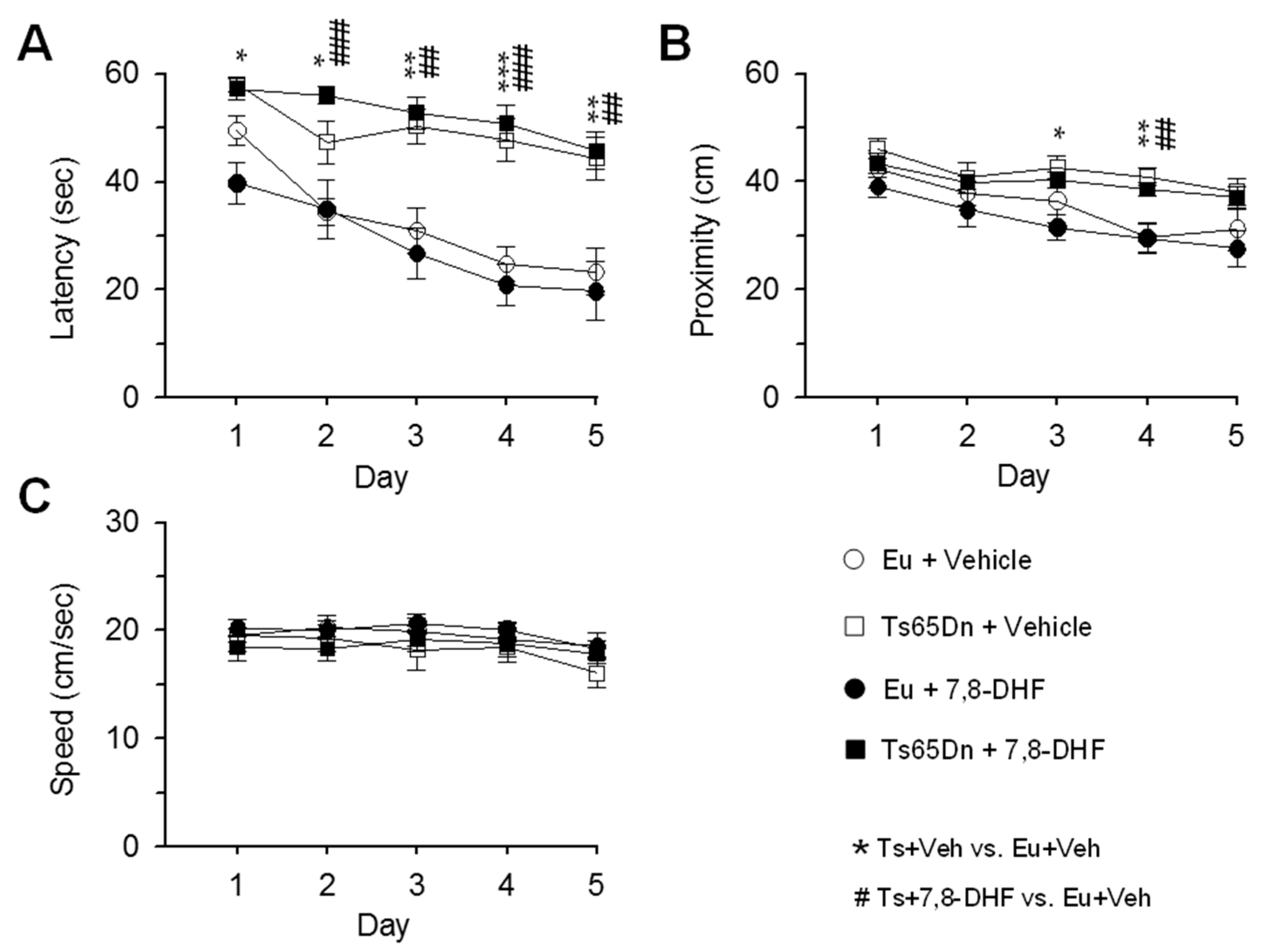
3.1. Long-Term Effects of Neonatal Treatment with 7,8-DHF on Hippocampus-Dependent Learning and Memory
3.2. Effects of Adult Treatment with 7,8-DHF on Hippocampus-Dependent Learning and Memory
3.3. Effects of Adult Treatment with 7,8-DHF on Neurogenesis
3.4. Effects of Adult Treatment with 7,8-DHF on the TrkB Receptor
4. Discussion
4.1. Neonatal Treatment with 7,8-DHF Fails to Induce Long-Lasting Effects on Hippocampus-Dependent Learning and Memory in Ts65Dn Mice
4.2. Adult Therapy with 7,8-DHF Does Not Replicate the Effects of Therapy in Young Ts65Dn Mice
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bartesaghi, R.; Guidi, S.; Ciani, E. Is it possible to improve neurodevelopmental abnormalities in Down syndrome? Rev. Neurosci. 2011, 22, 419–455. [Google Scholar] [CrossRef] [PubMed]
- Dierssen, M. Down syndrome: The brain in trisomic mode. Nat. Rev. Neurosci. 2012, 13, 844–858. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Jing, Y.; Cost, G.J.; Chiang, J.C.; Kolpa, H.J.; Cotton, A.M.; Carone, D.M.; Carone, B.R.; Shivak, D.A.; Guschin, D.Y.; et al. Translating dosage compensation to trisomy 21. Nature 2013, 500, 296–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardiner, K.J. Molecular basis of pharmacotherapies for cognition in Down syndrome. Trends Pharmacol. Sci. 2010, 31, 66–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stagni, F.; Giacomini, A.; Guidi, S.; Ciani, E.; Bartesaghi, R. Timing of therapies for Down syndrome: The sooner, the better. Front. Behav. Neurosci. 2015, 9, 265. [Google Scholar] [CrossRef] [PubMed]
- Moosavi, F.; Hosseini, R.; Saso, L.; Firuzi, O. Modulation of neurotrophic signaling pathways by polyphenols. Drug Des. Dev. Ther. 2016, 10, 23–42. [Google Scholar]
- Spencer, J.P. Flavonoids: Modulators of brain function? Br. J. Nutr. 2008, 99 (E Suppl. 1), ES60–ES77. [Google Scholar] [CrossRef]
- Williams, R.J.; Spencer, J.P. Flavonoids, cognition, and dementia: Actions, mechanisms, and potential therapeutic utility for Alzheimer disease. Free Radic. Biol. Med. 2012, 52, 35–45. [Google Scholar] [CrossRef]
- Rendeiro, C.; Rhodes, J.S.; Spencer, J.P.E. The mechanisms of action of flavonoids in the brain: Direct versus indirect effects. Neurochem. Int. 2015, 89, 126–139. [Google Scholar] [CrossRef]
- Toiber, D.; Azkona, G.; Ben-Ari, S.; Toran, N.; Soreq, H.; Dierssen, M. Engineering DYRK1A overdosage yields Down syndrome-characteristic cortical splicing aberrations. Neurobiol. Dis. 2010, 40, 348–359. [Google Scholar] [CrossRef]
- Guedj, F.; Sebrie, C.; Rivals, I.; Ledru, A.; Paly, E.; Bizot, J.C.; Smith, D.; Rubin, E.; Gillet, B.; Arbones, M.; et al. Green tea polyphenols rescue of brain defects induced by overexpression of DYRK1A. PLoS ONE 2009, 4, e4606. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, P.; Ciani, E.; Guidi, S.; Trazzi, S.; Felice, D.; Grossi, G.; Fernandez, M.; Giuliani, A.; Calza, L.; Bartesaghi, R. Early pharmacotherapy restores neurogenesis and cognitive performance in the Ts65Dn mouse model for Down syndrome. J. Neurosci. 2010, 30, 8769–8779. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, Y.; Berry, T.L.; Nelson, M.; Hunter, C.L.; Fukuhara, K.; Imai, H.; Ito, S.; Granholm-Bentley, A.C.; Kaplan, A.P.; Mutoh, T. Stimulated neuronal expression of brain-derived neurotrophic factor by Neurotropin. Mol. Cell. Neurosci. 2010, 45, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Begenisic, T.; Sansevero, G.; Baroncelli, L.; Cioni, G.; Sale, A. Early environmental therapy rescues brain development in a mouse model of Down syndrome. Neurobiol. Dis. 2015, 82, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Bimonte-Nelson, H.A.; Hunter, C.L.; Nelson, M.E.; Granholm, A.C. Frontal cortex BDNF levels correlate with working memory in an animal model of Down syndrome. Behav. Brain Res. 2003, 139, 47–57. [Google Scholar] [CrossRef]
- Jang, S.W.; Liu, X.; Yepes, M.; Shepherd, K.R.; Miller, G.W.; Liu, Y.; Wilson, W.D.; Xiao, G.; Blanchi, B.; Sun, Y.E.; et al. A selective TrkB agonist with potent neurotrophic activities by 7,8-dihydroxyflavone. Proc. Natl. Acad. Sci. USA 2010, 107, 2687–2692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stagni, F.; Giacomini, A.; Guidi, S.; Emili, M.; Uguagliati, B.; Salvalai, M.E.; Bortolotto, V.; Grilli, M.; Rimondini, R.; Bartesaghi, R. A flavonoid agonist of the TrkB receptor for BDNF improves hippocampal neurogenesis and hippocampus-dependent memory in the Ts65Dn mouse model of DS. Exp. Neurol. 2017, 298, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Reinholdt, L.G.; Ding, Y.; Gilbert, G.J.; Czechanski, A.; Solzak, J.P.; Roper, R.J.; Johnson, M.T.; Donahue, L.R.; Lutz, C.; Davisson, M.T. Molecular characterization of the translocation breakpoints in the Down syndrome mouse model Ts65Dn. Mamm. Genome 2011, 22, 685–691. [Google Scholar] [CrossRef] [Green Version]
- Giacomini, A.; Stagni, F.; Trazzi, S.; Guidi, S.; Emili, M.; Brigham, E.; Ciani, E.; Bartesaghi, R. Inhibition of APP gamma-secretase restores Sonic Hedgehog signaling and neurogenesis in the Ts65Dn mouse model of Down syndrome. Neurobiol. Dis. 2015, 82, 385–396. [Google Scholar] [CrossRef] [Green Version]
- Stagni, F.; Giacomini, A.; Emili, M.; Trazzi, S.; Guidi, S.; Sassi, M.; Ciani, E.; Rimondini, R.; Bartesaghi, R. Short- and long-term effects of neonatal pharmacotherapy with epigallocatechin-3-gallate on hippocampal development in the Ts65Dn mouse model of Down syndrome. Neuroscience 2016, 333, 277–301. [Google Scholar] [CrossRef]
- Giacomini, A.; Stagni, F.; Emili, M.; Guidi, S.; Salvalai, M.E.; Grilli, M.; Vidal-Sanchez, V.; Martinez-Cue, C.; Bartesaghi, R. Treatment with corn oil improves neurogenesis and cognitive performance in the Ts65Dn mouse model of Down syndrome. Brain Res. Bull. 2018, 140, 378–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guidi, S.; Stagni, F.; Bianchi, P.; Ciani, E.; Ragazzi, E.; Trazzi, S.; Grossi, G.; Mangano, C.; Calza, L.; Bartesaghi, R. Early pharmacotherapy with fluoxetine rescues dendritic pathology in the Ts65Dn mouse model of Down syndrome. Brain Pathol. 2013, 23, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Trazzi, S.; Mitrugno, V.M.; Valli, E.; Fuchs, C.; Rizzi, S.; Guidi, S.; Perini, G.; Bartesaghi, R.; Ciani, E. APP-dependent up-regulation of Ptch1 underlies proliferation impairment of neural precursors in Down syndrome. Hum. Mol. Genet. 2011, 20, 1560–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couillard-Despres, S.; Winner, B.; Schaubeck, S.; Aigner, R.; Vroemen, M.; Weidner, N.; Bogdahn, U.; Winkler, J.; Kuhn, H.G.; Aigner, L. Doublecortin expression levels in adult brain reflect neurogenesis. Eur. J. Neurosci. 2005, 21, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chan, C.-B.; Jang, S.-W.; Pradoldej, S.; Huang, J.; He, K.; Phun, L.H.; France, S.; Xiao, G.; Jia, Y.; et al. A synthetic 7,8-dihydroxyflavone derivative promotes neurogenesis and exhibits potent antidepressant effect. J. Med. Chem. 2010, 53, 8274–8286. [Google Scholar] [CrossRef]
- Gao, L.; Tian, M.; Zhao, H.Y.; Xu, Q.Q.; Huang, Y.M.; Si, Q.C.; Tian, Q.; Wu, Q.M.; Hu, X.M.; Sun, L.B.; et al. TrkB activation by 7,8-dihydroxyflavone increases synapse AMPA subunits and ameliorates spatial memory deficits in a mouse model of Alzheimer’s disease. J. Neurochem. 2016, 136, 620–636. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, X.; Schroeder, J.P.; Chan, C.-B.; Song, M.; Yu, S.P.; Weinshenker, D.; Ye, K. 7,8-dihydroxyflavone prevents synaptic loss and memory deficits in a mouse model of Alzheimer’s disease. Neuropsychopharmacology 2014, 39, 638–650. [Google Scholar] [CrossRef] [PubMed]
- Altman, J.; Bayer, S. Postnatal development of the hippocampal dentate gyrus under normal and experimental conditions. In The Hippocampus; Isaacson, R.L., Pribram, K.H., Eds.; Plenum Press: New York, NY, USA; London, UK, 1975; Volume 1, pp. 95–122. [Google Scholar]
- Altman, J.; Bayer, S.A. Migration and distribution of two populations of hippocampal granule cell precursors during the perinatal and postnatal periods. J. Comp. Neurol. 1990, 301, 365–381. [Google Scholar] [CrossRef]
- Altman, J.; Bayer, S.A. Mosaic organization of the hippocampal neuroepithelium and the multiple germinal sources of dentate granule cells. J. Comp. Neurol. 1990, 301, 325–342. [Google Scholar] [CrossRef]
- Workman, A.D.; Charvet, C.J.; Clancy, B.; Darlington, R.B.; Finlay, B.L. Modeling transformations of neurodevelopmental sequences across mammalian species. J. Neurosci. 2013, 33, 7368–7383. [Google Scholar] [CrossRef]
- Stagni, F.; Raspanti, A.; Giacomini, A.; Guidi, S.; Emili, M.; Ciani, E.; Giuliani, A.; Bighinati, A.; Calza, L.; Magistretti, J.; et al. Long-term effect of neonatal inhibition of APP gamma-secretase on hippocampal development in the Ts65Dn mouse model of Down syndrome. Neurobiol. Dis. 2017, 103, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Guidi, S.; Stagni, F.; Bianchi, P.; Ciani, E.; Giacomini, A.; De Franceschi, M.; Moldrich, R.; Kurniawan, N.; Mardon, K.; Giuliani, A.; et al. Prenatal pharmacotherapy rescues brain development in a Down’s syndrome mouse model. Brain 2014, 137, 380–401. [Google Scholar] [CrossRef] [PubMed]
- Velazquez, R.; Ash, J.A.; Powers, B.E.; Kelley, C.M.; Strawderman, M.; Luscher, Z.I.; Ginsberg, S.D.; Mufson, E.J.; Strupp, B.J. Maternal choline supplementation improves spatial learning and adult hippocampal neurogenesis in the Ts65Dn mouse model of Down syndrome. Neurobiol. Dis. 2013, 58, 92–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, J.; Chen, M.; Gandhy, S.U.; Strawderman, M.; Levitsky, D.A.; Maclean, K.N.; Strupp, B.J. Perinatal choline supplementation improves cognitive functioning and emotion regulation in the Ts65Dn mouse model of Down syndrome. Behav. Neurosci. 2010, 124, 346–361. [Google Scholar] [CrossRef] [PubMed]
- Nakano-Kobayashi, A.; Awaya, T.; Kii, I.; Sumida, Y.; Okuno, Y.; Yoshida, S.; Sumida, T.; Inoue, H.; Hosoya, T.; Hagiwara, M. Prenatal neurogenesis induction therapy normalizes brain structure and function in Down syndrome mice. Proc. Natl. Acad. Sci. USA 2017, 114, 10268–10273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stagni, F.; Giacomini, A.; Guidi, S.; Ciani, E.; Ragazzi, E.; Filonzi, M.; De Iasio, R.; Rimondini, R.; Bartesaghi, R. Long-term effects of neonatal treatment with fluoxetine on cognitive performance in Ts65Dn mice. Neurobiol. Dis. 2015, 74, 204–218. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chua, K.W.; Chua, C.C.; Yu, H.; Pei, A.; Chua, B.H.; Hamdy, R.C.; Xu, X.; Liu, C.F. Antioxidant activity of 7,8-dihydroxyflavone provides neuroprotection against glutamate-induced toxicity. Neurosci. Lett. 2011, 499, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zhu, S.; Wang, B.; Chen, L.; Li, R.; Yao, W.; Qu, Z. Antioxidant action of 7,8-dihydroxyflavone protects PC12 cells against 6-hydroxydopamine-induced cytotoxicity. Neurochem. Int. 2014, 64, 18–23. [Google Scholar] [CrossRef]
- Valenti, D.; de Bari, L.; de Rasmo, D.; Signorile, A.; Henrion-Caude, A.; Contestabile, A.; Vacca, R.A. The polyphenols resveratrol and epigallocatechin-3-gallate restore the severe impairment of mitochondria in hippocampal progenitor cells from a Down syndrome mouse model. Biochim. Biophys. Acta 2016, 1862, 1093–1104. [Google Scholar] [CrossRef]
- Marongiu, D.; Imbrosci, B.; Mittmann, T. Modulatory effects of the novel TrkB receptor agonist 7,8-dihydroxyflavone on synaptic transmission and intrinsic neuronal excitability in mouse visual cortex In Vitro. Eur. J. Pharmacol. 2013, 709, 64–71. [Google Scholar] [CrossRef]
- Kleschevnikov, A.M.; Belichenko, P.V.; Gall, J.; George, L.; Nosheny, R.; Maloney, M.T.; Salehi, A.; Mobley, W.C. Increased efficiency of the GABAA and GABAB receptor-mediated neurotransmission in the Ts65Dn mouse model of Down syndrome. Neurobiol. Dis. 2012, 45, 683–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Chan, C.B.; Ye, K. 7,8-dihydroxyflavone, a small molecular TrkB agonist, is useful for treating various BDNF-implicated human disorders. Transl. Neurodegener. 2016, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Devi, L.; Ohno, M. 7,8-dihydroxyflavone, a small-molecule TrkB agonist, reverses memory deficits and BACE1 elevation in a mouse model of Alzheimer’s disease. Neuropsychopharmacology 2012, 37, 434–444. [Google Scholar] [CrossRef]
- Andero, R.; Daviu, N.; Escorihuela, R.M.; Nadal, R.; Armario, A. 7,8-dihydroxyflavone, a TrkB receptor agonist, blocks long-term spatial memory impairment caused by immobilization stress in rats. Hippocampus 2012, 22, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Andero, R.; Heldt, S.A.; Ye, K.; Liu, X.; Armario, A.; Ressler, K.J. Effect of 7,8-dihydroxyflavone, a small-molecule TrkB agonist, on emotional learning. Am. J. Psychiatry 2011, 168, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Lv, F.; Li, L.; Yu, H.; Dong, M.; Fu, Q. 7,8-dihydroxyflavone rescues spatial memory and synaptic plasticity in cognitively impaired aged rats. J. Neurochem. 2012, 122, 800–811. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Peng, Q.; Liu, X.; Jin, J.; Hou, Z.; Zhang, J.; Mori, S.; Ross, C.A.; Ye, K.; Duan, W. Small-molecule TrkB receptor agonists improve motor function and extend survival in a mouse model of Huntington’s disease. Hum. Mol. Genet. 2013, 22, 2462–2470. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, O.T.; Aytan, N.; Carreras, I.; Choi, J.K.; Kowall, N.W.; Jenkins, B.G.; Dedeoglu, A. 7,8-Dihydroxyflavone improves motor performance and enhances lower motor neuronal survival in a mouse model of amyotrophic lateral sclerosis. Neurosci. Lett. 2014, 566, 286–291. [Google Scholar] [CrossRef] [Green Version]
- Parrini, M.; Ghezzi, D.; Deidda, G.; Medrihan, L.; Castroflorio, E.; Alberti, M.; Baldelli, P.; Cancedda, L.; Contestabile, A. Aerobic exercise and a BDNF-mimetic therapy rescue learning and memory in a mouse model of Down syndrome. Sci. Rep. 2017, 7, 16825. [Google Scholar] [CrossRef]
- Liu, X.; Obianyo, O.; Chan, C.B.; Huang, J.; Xue, S.; Yang, J.J.; Zeng, F.; Goodman, M.; Ye, K. Biochemical and biophysical investigation of the brain-derived neurotrophic factor mimetic 7,8-dihydroxyflavone in the binding and activation of the TrkB receptor. J. Biol. Chem. 2014, 289, 27571–27584. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| - | Latency (s) | |||||
| D1 | D2 | D3 | D4 | D5 | ||
| Eu + Veh | Eu + 7,8-DHF | 0.007 | 0.789 | 0.177 | 0.207 | 0.236 |
| Ts65Dn + Veh | 0.035 | 0.044 | 0.007 | <0.001 | 0.008 | |
| Ts65Dn + 7,8-DHF | 0.066 | 0.001 | 0.003 | <0.001 | 0.006 | |
| Eu + 7,8-DHF | Ts65Dn + Veh | <0.001 | 0.024 | <0.001 | <0.001 | <0.001 |
| Ts65Dn + 7,8-DHF | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Ts65Dn + Veh | Ts65Dn + 7,8-DHF | 0.823 | 0.129 | 0.664 | 0.575 | 0.829 |
| - | Proximity (cm) | |||||
| D1 | D2 | D3 | D4 | D5 | ||
| Eu + Veh | Eu + 7,8-DHF | 0.565 | 0.647 | 0.110 | 0.471 | 0.209 |
| Ts65Dn + Veh | 0.062 | 0.204 | 0.045 | 0.003 | 0.098 | |
| Ts65Dn + 7,8-DHF | 0.270 | 0.169 | 0.100 | 0.006 | 0.146 | |
| Eu + 7,8-DHF | Ts65Dn + Veh | 0.017 | 0.090 | 0.001 | <0.001 | 0.006 |
| Ts65Dn + 7,8-DHF | 0.103 | 0.074 | 0.003 | 0.001 | 0.010 | |
| Ts65Dn + Veh | Ts65Dn + 7,8-DHF | 0.465 | 0.887 | 0.743 | 0.840 | 0.872 |
| - | Swim Speed (cm/s) | |||||
| D1 | D2 | D3 | D4 | D5 | ||
| Eu + Veh | Eu + 7,8-DHF | 0.632 | 0.953 | 0.689 | 0.485 | 0.976 |
| Ts65Dn + Veh | 0.974 | 0.668 | 0.335 | 0.680 | 0.117 | |
| Ts65Dn + 7,8-DHF | 0.443 | 0.292 | 0.681 | 0.830 | 0.711 | |
| Eu + 7,8-DHF | Ts65Dn + Veh | 0.617 | 0.627 | 0.179 | 0.277 | 0.123 |
| Ts65Dn + 7,8-DHF | 0.225 | 0.267 | 0.431 | 0.382 | 0.732 | |
| Ts65Dn + Veh | Ts65Dn + 7,8-DHF | 0.472 | 0.529 | 0.603 | 0.855 | 0.253 |
| - | Latency (s) | ||||||||
| D1 | D2 | D3 | D4 | D5 | D6 | D7 | D8 | ||
| Eu + Veh | Eu + 7,8-DHF | 0.296 | 0.135 | 0.170 | 0.281 | 0.078 | 0.051 | 0.293 | 0.209 |
| Ts65Dn + Veh | 0.017 | 0.006 | 0.001 | 0.003 | 0.022 | 0.060 | 0.079 | 0.282 | |
| Ts65Dn + 7,8-DHF | 0.018 | 0.005 | 0.083 | 0.002 | 0.108 | 0.073 | 0.174 | 0.194 | |
| Eu + 7,8-DHF | Ts65Dn + Veh | 0.283 | 0.331 | <0.001 | 0.001 | 0.001 | 0.001 | 0.015 | 0.037 |
| Ts65Dn + 7,8-DHF | 0.217 | 0.204 | 0.008 | <0.001 | 0.004 | 0.002 | 0.038 | 0.029 | |
| Ts65Dn + Veh | Ts65Dn + 7,8-DHF | 0.767 | 0.659 | 0.144 | 0.585 | 0.668 | 0.882 | 0.852 | 0.722 |
| - | Proximity (cm) | ||||||||
| D1 | D2 | D3 | D4 | D5 | D6 | D7 | D8 | ||
| Eu+Veh | Eu + 7,8-DHF | 0.338 | 0.082 | 0.005 | 0.016 | 0.004 | 0.018 | 0.012 | 0.006 |
| Ts65Dn + Veh | 0.025 | 0.097 | 0.424 | 0.389 | 0.539 | 0.538 | 0.345 | 0.840 | |
| Ts65Dn + 7,8-DHF | 0.882 | 0.660 | 0.349 | 0.541 | 0.834 | 0.640 | 0.747 | 0.883 | |
| Eu + 7,8-DHF | Ts65Dn + Veh | 0.006 | 0.003 | 0.001 | 0.003 | 0.001 | 0.006 | 0.002 | 0.005 |
| Ts65Dn + 7,8-DHF | 0.476 | 0.058 | 0.080 | 0.009 | 0.016 | 0.087 | 0.050 | 0.020 | |
| Ts65Dn + Veh | Ts65Dn + 7,8-DHF | 0.041 | 0.316 | 0.117 | 0.889 | 0.470 | 0.332 | 0.268 | 0.754 |
| - | Swim speed (cm/s) | ||||||||
| D1 | D2 | D3 | D4 | D5 | D6 | D7 | D8 | ||
| Eu + Veh | Eu + 7,8-DHF | 0.531 | 0.954 | 0.789 | 0.473 | 0.453 | 0.152 | 0.730 | 0.343 |
| Ts65Dn + Veh | 0.620 | 0.292 | 0.821 | 0.749 | 0.815 | 0.929 | 0.654 | 0.689 | |
| Ts65Dn + 7,8-DHF | 0.018 | 0.104 | 0.150 | 0.913 | 0.616 | 0.119 | 0.484 | 0.071 | |
| Eu + 7,8-DHF | Ts65Dn + Veh | 0.477 | 0.401 | 0.947 | 0.670 | 0.356 | 0.144 | 0.476 | 0.559 |
| Ts65Dn + 7,8-DHF | 0.180 | 0.166 | 0.298 | 0.468 | 0.827 | 0.908 | 0.754 | 0.435 | |
| Ts65Dn + Veh | Ts65Dn + 7,8-DHF | 0.432 | 0.480 | 0.225 | 0.706 | 0.494 | 0.113 | 0.292 | 0.152 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giacomini, A.; Stagni, F.; Emili, M.; Uguagliati, B.; Rimondini, R.; Bartesaghi, R.; Guidi, S. Timing of Treatment with the Flavonoid 7,8-DHF Critically Impacts on Its Effects on Learning and Memory in the Ts65Dn Mouse. Antioxidants 2019, 8, 163. https://doi.org/10.3390/antiox8060163
Giacomini A, Stagni F, Emili M, Uguagliati B, Rimondini R, Bartesaghi R, Guidi S. Timing of Treatment with the Flavonoid 7,8-DHF Critically Impacts on Its Effects on Learning and Memory in the Ts65Dn Mouse. Antioxidants. 2019; 8(6):163. https://doi.org/10.3390/antiox8060163
Chicago/Turabian StyleGiacomini, Andrea, Fiorenza Stagni, Marco Emili, Beatrice Uguagliati, Roberto Rimondini, Renata Bartesaghi, and Sandra Guidi. 2019. "Timing of Treatment with the Flavonoid 7,8-DHF Critically Impacts on Its Effects on Learning and Memory in the Ts65Dn Mouse" Antioxidants 8, no. 6: 163. https://doi.org/10.3390/antiox8060163
APA StyleGiacomini, A., Stagni, F., Emili, M., Uguagliati, B., Rimondini, R., Bartesaghi, R., & Guidi, S. (2019). Timing of Treatment with the Flavonoid 7,8-DHF Critically Impacts on Its Effects on Learning and Memory in the Ts65Dn Mouse. Antioxidants, 8(6), 163. https://doi.org/10.3390/antiox8060163