Assessment of In Vitro Bioactivities of Polysaccharides Isolated from Hericium Novae-Zealandiae


 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Polysaccharide Extraction and Purification
2.3. Protein Content in Polysaccharides
2.4. Anti-Proliferation Assay
2.5. Gene Expression
2.6. AChE Inhibition
2.7. Determination of Antioxidant Capacity
2.7.1. DPPH Assay
2.7.2. FRAP
2.7.3. ABTS
2.8. Statistical Analysis
3. Results and Discussion
3.1. Polysaccharide Yield and Protein Content in Polysaccharides
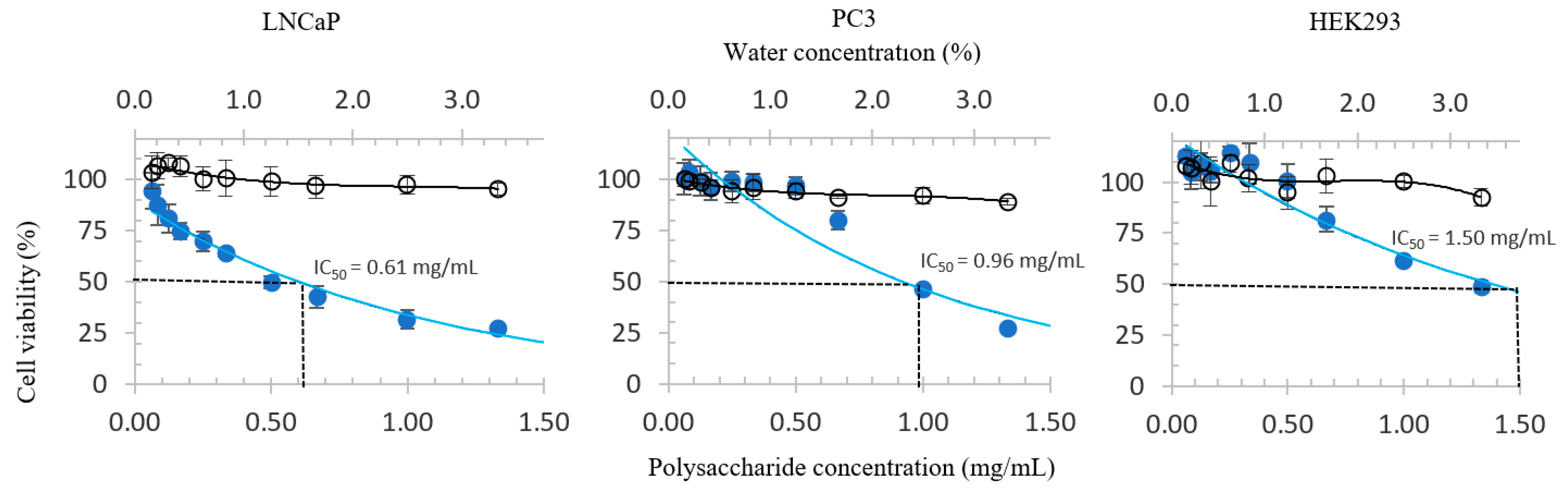
3.2. Proliferative Inhibition of PCa and HEK293 Cells
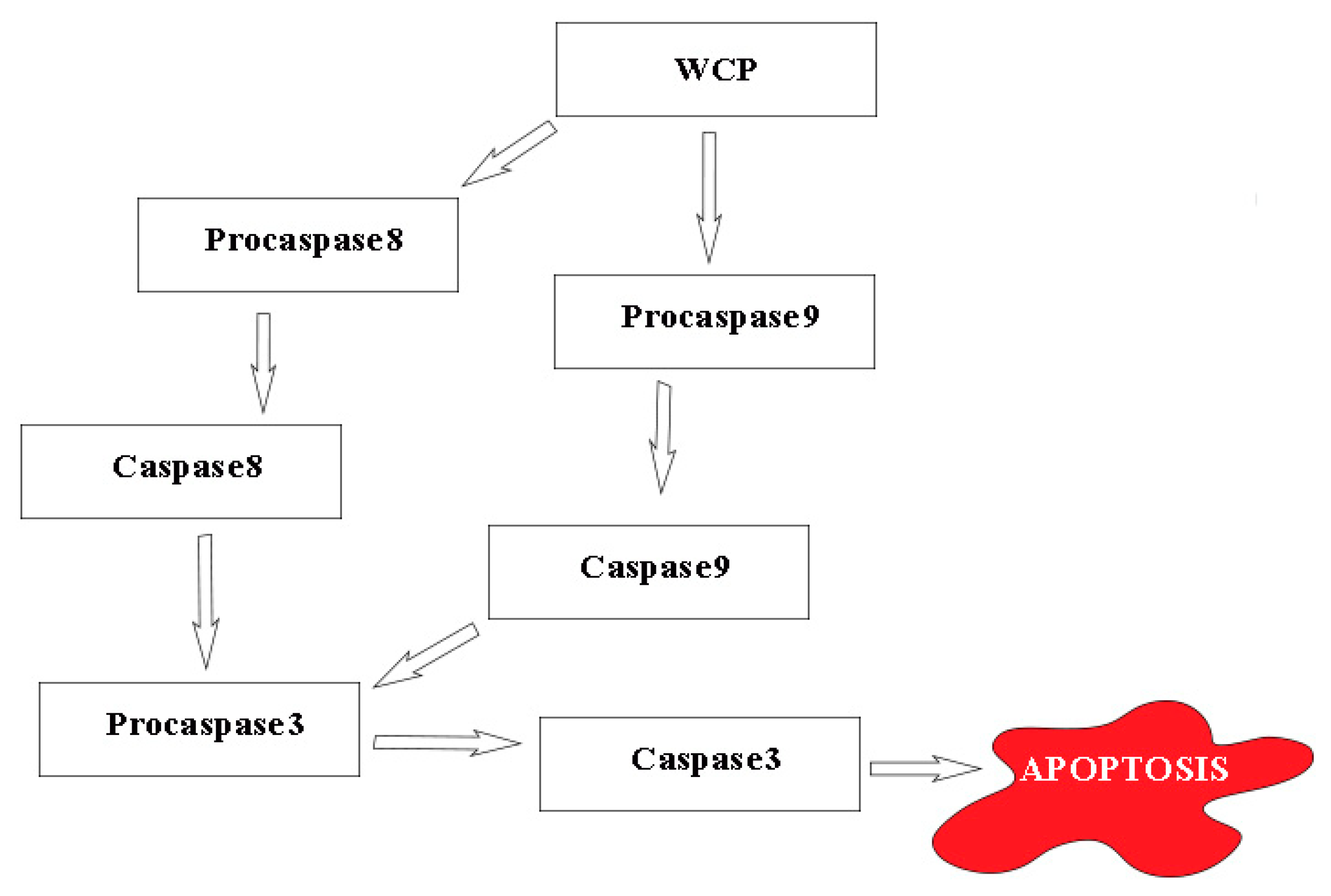
3.3. RT-qPCR
3.3.1. LNCaP
3.3.2. PC3
3.4. AChE Inhibition
3.5. Antioxidant Activities
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moradali, M.-F.; Mostafavi, H.; Ghods, S.; Hedjaroude, G.-A. Immunomodulating and anticancer agents in the realm of macromycetes fungi (macrofungi). Int. Immunopharmacol. 2007, 7, 701–724. [Google Scholar] [CrossRef] [PubMed]
- De Silva, D.D.; Rapior, S.; Fons, F.; Bahkali, A.H.; Hyde, K.D. Medicinal mushrooms in supportive cancer therapies: An approach to anti-cancer effects and putative mechanisms of action. Fungal Divers. 2012, 55, 1–35. [Google Scholar] [CrossRef]
- Rathore, H.; Prasad, S.; Sharma, S. Mushroom nutraceuticals for improved nutrition and better human health: A review. PharmaNutrition 2017, 5, 35–46. [Google Scholar] [CrossRef]
- Guo, C.; Choi, M.W.; Cheung, P.C.K. Mushroom and immunity. Curr. Top. Nutraceutical Res. 2012, 10, 31–42. [Google Scholar]
- Fuller, R.J.M.; Buchanan, P.K.; Roberts, M. Maori knowledge of fungi/Mātauranga o ngā harore. In The Fungi of New Zealand/Ngā Harore o Aotearoa Volume 1. Introduction to Fungi of New Zealand; McKenzie, E.H.C., Ed.; Hong Kong Fungal Diversity Press: Hong Kong, China, 2004; pp. 81–118. [Google Scholar]
- Wu, F.; Zhou, C.; Zhou, D.; Ou, S.; Zhang, X.; Huang, H. Structure characterization of a novel polysaccharide from: Hericium erinaceus fruiting bodies and its immunomodulatory activities. Food Funct. 2018, 9, 294–306. [Google Scholar] [CrossRef]
- Kuo, H.C.; Kuo, Y.R.; Lee, K.F.; Hsieh, M.C.; Huang, C.Y.; Hsieh, Y.Y.; Lee, K.C.; Kuo, H.L.; Lee, L.Y.; Chen, W.P.; et al. A Comparative Proteomic Analysis of Erinacine A’s Inhibition of Gastric Cancer Cell Viability and Invasiveness. Cell. Physiol. Biochem. 2017, 43, 195–208. [Google Scholar] [CrossRef]
- Lee, S.R.; Jung, K.; Noh, H.J.; Park, Y.J.; Lee, H.L.; Lee, K.R.; Kang, K.S.; Kim, K.H. A new cerebroside from the fruiting bodies of Hericium erinaceus and its applicability to cancer treatment. Biorgan. Med. Chem. Lett. 2015, 25, 5712–5715. [Google Scholar] [CrossRef]
- Kim, S.P.; Nam, S.H.; Friedman, M. Erratum: Hericium erinaceus (Lion’s mane) mushroom extracts inhibit metastasis of cancer cells to the lung in CT-26 colon cancer-transplanted mice (Journal of the Agricultural Food Chemistry (2013) 61: 20 (4898–4904). J. Agric. Food Chem. 2014, 62, 528. [Google Scholar] [CrossRef]
- Jang, H.J.; Kim, J.E.; Jeong, K.H.; Lim, S.C.; Kim, S.Y.; Cho, K.O. The Neuroprotective Effect of Hericium erinaceus Extracts in Mouse Hippocampus after Pilocarpine-Induced Status Epilepticus. Int. J. Mol. Sci. 2019, 20, 859. [Google Scholar] [CrossRef]
- Wang, X.Y.; Yin, J.Y.; Zhao, M.M.; Liu, S.Y.; Nie, S.P.; Xie, M.Y. Gastroprotective activity of polysaccharide from Hericium erinaceus against ethanol-induced gastric mucosal lesion and pylorus ligation-induced gastric ulcer, and its antioxidant activities. Carbohydr. Polym. 2018, 186, 100–109. [Google Scholar] [CrossRef]
- Noh, H.J.; Yoon, J.Y.; Kim, G.S.; Lee, S.E.; Lee, D.Y.; Choi, J.H.; Kim, S.Y.; Kang, K.S.; Cho, J.Y.; Kim, K.H. Benzyl alcohol derivatives from the mushroom Hericium erinaceum attenuate LPS-stimulated inflammatory response through the regulation of NF-κB and AP-1 activity. Immunopharmacol. Immunotoxicol. 2014, 36, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Tania, M.; Liu, R.; Rahman, M.M. Hericium erinaceus: An edible mushroom with medicinal values. J. Complement. Integr. Med. 2013, 10, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Buchanan, P.; Quek, S.Y. Development and validation of an HPLC-DAD-MS method for determination of four nucleoside compounds in the New Zealand native mushroom Hericium sp. Food Chem. 2019, 278, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, Y.; Yang, S.; Zhao, D.; Wang, M. A polysaccharide from cultured mycelium of Hericium erinaceus relieves ulcerative colitis by counteracting oxidative stress and improving mitochondrial function. Int. J. Biol. Macromol. 2019, 125, 572–579. [Google Scholar] [CrossRef]
- Li, Q.Z.; Wu, D.; Zhou, S.; Liu, Y.F.; Li, Z.P.; Feng, J.; Yang, Y. Structure elucidation of a bioactive polysaccharide from fruiting bodies of Hericium erinaceus in different maturation stages. Carbohydr. Polym. 2016, 144, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.H.; Ye, J.M.; Wang, G.F. Evaluation of in vivo antioxidant activity of Hericium erinaceus polysaccharides. Int. J. Biol. Macromol. 2013, 52, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, Y.; Xiao, X.; Xu, D.; Gao, Y.; Gao, Q. A polysaccharide isolated from mycelia of the lion’s mane medicinal mushroom Hericium erinaceus (Agaricomycetes) induced apoptosis in precancerous human gastric cells. Int. J. Med. Mushrooms 2017, 19, 1053–1060. [Google Scholar] [CrossRef] [PubMed]
- Zan, X.; Cui, F.; Li, Y.; Yang, Y.; Wu, D.; Sun, W.; Ping, L. Hericium erinaceus polysaccharide-protein HEG-5 inhibits SGC-7901 cell growth via cell cycle arrest and apoptosis. Int. J. Biol. Macromol. 2015, 76, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Yong, Y.; Gu, Y.; Wang, Z.; Zhang, S.; Lu, L. Comparison of antioxidant and antiproliferation activities of polysaccharides from eight species of medicinal mushrooms. Int. J. Med. Mushrooms 2015, 17, 287–295. [Google Scholar] [CrossRef]
- Cheng, J.H.; Tsai, C.L.; Lien, Y.Y.; Lee, M.S.; Sheu, S.C. High molecular weight of polysaccharides from Hericium erinaceus against amyloid beta-induced neurotoxicity. BMC Complement. Altern. Med. 2016, 16, 170. [Google Scholar] [CrossRef]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2014, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Health, M.O. New Cancer Registrations. Available online: https://www.health.govt.nz/publication/new-cancer-registrations-2016 (accessed on 7 May 2019).
- Verma, O.; Kumar, A.; Kumar, P. Anticancer Potential of Plants and Natural Products: A Review. Am. J. Pharmacol. Sci. 2013, 1, 104–115. [Google Scholar] [Green Version]
- Cunningham, D.; You, Z. In vitro and in vivo model systems used in prostate cancer research. J. Biol. Methods 2015, 2, e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirier, J.; Delisle, M.C.; Quirion, R.; Aubert, I.; Farlow, M.; Lahiri, D.; Hui, S.; Bertrand, P.; Nalbantoglu, J.; Gilfix, B.M. Apolipoprotein E4 allele as a predictor of cholinergic deficits and treatment outcome in Alzheimer disease. Proc. Natl. Acad. Sci. USA 1995, 92, 12260. [Google Scholar] [CrossRef] [PubMed]
- Enz, A.; Amstutz, R.; Boddeke, H.; Gmelin, G.; Malanowski, J. Chapter 53: Brain selective inhibition of acetylcholinesterase: A novel approach to therapy for Alzheimer’s disease. In Progress in Brain Research; Cuello, A.C., Ed.; Elsevier: Amsterdam, The Netherlands, 1993; Volume 98, pp. 431–438. [Google Scholar]
- Mukherjee, P.K.; Kumar, V.; Houghton, P.J. Screening of Indian medicinal plants for acetylcholinesterase inhibitory activity. Phytother. Res. 2007, 21, 1142–1145. [Google Scholar] [CrossRef] [PubMed]
- Grassi, D.; Desideri, G.; Tiberti, S.; Ferri, C. Oxidative stress, endothelial dysfunction and prevention of cardiovascular diseases. Agro Food Ind. Hi Tech 2009, 20, 8–11. [Google Scholar]
- Anand, R.; Gill, K.D.; Mahdi, A.A. Therapeutics of Alzheimer’s disease: Past, present and future. Neuropharmacology 2014, 76, 27–50. [Google Scholar] [CrossRef]
- Jia, L.N.; Liu, L.; Dong, Q.; Fang, J.N. Main polysaccharide fractions isolated from fruiting bodies of Hericium erinaceus. Chin. Tradit. Herb. Drugs 2005, 36, 10–12. [Google Scholar]
- Sevag, M.G. Eine neue physikalische Enteiweißungsmethode zur Darstellung biologisch wirksamer Substanzen. Biochem. Z 1934, 419, 102. [Google Scholar]
- Sedmak, J.J.; Grossberg, S.E. A rapid, sensitive, and versatile assay for protein using Coomassie brilliant blue G250. Anal. Biochem. 1977, 79, 544–552. [Google Scholar] [CrossRef]
- Vistica, D.; McMahon, J.; Warren, J.T.; Skehan, P.; Storeng, R.; Kenney, S.; Monks, A.; Scudiero, D.; Bokesch, H.; Boyd, M.R. New Colorimetric Cytotoxicity Assay for Anticancer-Drug Screening. J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar]
- Chaib, H.; Cockrell, E.K.; Rubin, M.A.; Macoska, J.A. Profiling and Verification of Gene Expression Patterns in Normal and Malignant Human Prostate Tissues by cDNA Microarray Analysis. Neoplasia 2001, 3, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Benabent, M.; Vilanova, E.; Sogorb, M.Á.; Estévez, J. Cholinesterase assay by an efficient fixed time endpoint method. Methods X 2014, 1, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Li, X.; Zhang, B.; Chen, P.X.; Liu, R.; Tsao, R. Characterisation of phenolics, betanins and antioxidant activities in seeds of three Chenopodium quinoa Willd. genotypes. Food Chem. 2015, 166, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Du, G.; Li, M.; Ma, F.; Liang, D. Antioxidant capacity and the relationship with polyphenol and Vitamin C in Actinidia fruits. Food Chem. 2009, 113, 557–562. [Google Scholar] [CrossRef]
- Fatima, I.; Ahmad, I.; Anis, I.; Malik, A.; Afza, N.; Iqbal, L.; Latif, M. New butyrylcholinesterase inhibitory steroid and peroxy acid from Leucas urticifolia. Arch. Pharm. Res. 2008, 31, 999–1003. [Google Scholar] [CrossRef]
- Singh, A.V.; Xiao, D.; Lew, K.L.; Dhir, R.; Singh, S.V. Sulforaphane induces caspase-mediated apoptosis in cultured PC-3 human prostate cancer cells and retards growth of PC-3 xenografts in vivo. Carcinogenesis 2004, 25, 83–90. [Google Scholar] [CrossRef]
- Bowen, C.; Voeller, H.J.; Kikly, K.; Gelmann, E.P. Synthesis of procaspases-3 and -7 during apoptosis in prostate cancer cells. Cell Death Differ. 1999, 6, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turan-Zitouni, G.; Ozdemir, A.; Kaplancikli, Z.A.; Altintop, M.D.; Temel, H.E.; Çiftçi, G.A. Synthesis and biological evaluation of some thiazole derivatives as new cholinesterase inhibitors. J. Enzyme Inhib. Med. Chem. 2013, 28, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; An, S.; Hu, W.; Teng, M.; Wang, X.; Qu, Y.; Liu, Y.; Yuan, Y.; Wang, D. The Neuroprotective Properties of Hericium erinaceus in Glutamate-Damaged Differentiated PC12 Cells and an Alzheimer’s Disease Mouse Model. Int. J. Mol. Sci. 2016, 17, 1810. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Polysaccharides | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| WCP (mg/mL) | 1.333 | 1.000 | 0.667 | 0.500 | 0.333 | 0.250 | 0.167 | 0.125 | 0.083 | 0.062 |
| BP1 (mg/mL) | 1.000 | 0.750 | 0.500 | 0.375 | 0.250 | 0.188 | 0.125 | 0.094 | 0.063 | 0.047 |
| Water control (%) | 3.333 | 2.500 | 1.666 | 1.250 | 0.833 | 0.625 | 0.417 | 0.312 | 0.228 | 0.156 |
| Gene | Assay ID | Human Target Chromosome Location | Amplicon Length (bp) |
|---|---|---|---|
| CASP3 | Hs00991555_m1 | Chr.4: 184627696 - 184649475 | 118 |
| CASP8 | Hs01018160_m1 | Chr.2: 201233443 - 201287711 | 124 |
| CASP9 | Hs00962278_m1 | Chr.1: 15491401 - 15524912 | 84 |
| GAPDH | Hs99999905_m1 | Chr.12: 6534405 - 6538375 | 122 |
| HPRT1 | Hs99999909_m1 | Chr.X: 134460145 - 134500668 | 100 |
| IL24 | Hs01114274_m1 | Chr.1: 206897404 - 206904139 | 67 |
| IL6 | Hs00174131_m1 | Chr.7: 22725889 - 22732002 | 95 |
| Polysaccharides | DU145 | PC3 | LNCaP | HEK293 |
|---|---|---|---|---|
| WCP (mg/mL) | >> 1.33 | 0.96 ± 0.10 | 0.61 ± 0.05 * | 1.50 ± 0.12 |
| BP1 (mg/mL) | >> 1.00 | >> 1.00 | 1.02 ± 0.12 * | 1.12 ± 0.11 |
| BP2 (mg/mL) | x | x | x | x |
| - | LNCaP | PC3 | ||
|---|---|---|---|---|
| GAPDH | HPRT1 | GAPDH | HPRT1 | |
| CASP3 | 1.18 ± 0.14 | 2.10±0.15 * | 1.44 ± 0.11 | 1.10 ± 0.15 |
| CASP8 | 0.93 ± 0.79 | 3.89±0.41 * | 1.54 ± 0.11 | 1.49 ± 0.12 |
| CASP9 | 1.45 ± 0.16 | 3.14±0.26 * | 0.79 ± 0.08 | 0.82 ± 0.09 |
| IL6 | X | X | 1.02 ± 0.11 | 0.52±0.05 * |
| IL24 | X | X | 5.67±0.48 * | 3.61±0.29 * |
| Activity | WCP | BP1 | BP2 |
|---|---|---|---|
| AChE inhibition (mg/mL IC50) | 8.26 ± 0.52 | 8.64 ± 0.45 | x |
| DPPH (µmol TE/g) | 59.74 ± 1.52 | x | x |
| FRAP (µmol TE/g) | 48.24±1.27 * | 41.49 ± 1.24 | 32.18 ± 0.97 |
| ABTS (µmol TE/g) | 65.58 ± 1.87 | 83.95±2.59 * | 58.35 ± 1.57 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Bishop, K.S.; Tanambell, H.; Buchanan, P.; Quek, S.Y. Assessment of In Vitro Bioactivities of Polysaccharides Isolated from Hericium Novae-Zealandiae. Antioxidants 2019, 8, 211. https://doi.org/10.3390/antiox8070211
Chen Z, Bishop KS, Tanambell H, Buchanan P, Quek SY. Assessment of In Vitro Bioactivities of Polysaccharides Isolated from Hericium Novae-Zealandiae. Antioxidants. 2019; 8(7):211. https://doi.org/10.3390/antiox8070211
Chicago/Turabian StyleChen, Zhixia (Grace), Karen Suzanne Bishop, Hartono Tanambell, Peter Buchanan, and Siew Young Quek. 2019. "Assessment of In Vitro Bioactivities of Polysaccharides Isolated from Hericium Novae-Zealandiae" Antioxidants 8, no. 7: 211. https://doi.org/10.3390/antiox8070211
APA StyleChen, Z., Bishop, K. S., Tanambell, H., Buchanan, P., & Quek, S. Y. (2019). Assessment of In Vitro Bioactivities of Polysaccharides Isolated from Hericium Novae-Zealandiae. Antioxidants, 8(7), 211. https://doi.org/10.3390/antiox8070211