Screening of In Vitro Health Benefits of Tangerine Tomatoes




Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Tomato Varieties and Extracts
2.3. Prostate Cancer Cell Lines and Culture Media
2.4. Cell Lines and Culture Media for the Anti-Inflammatory Assays
2.5. Antioxidant Assays
2.5.1. FRAP Assay
2.5.2. ABTS Assay
2.6. Prostate Cancer Proliferation Assay
2.7. Anti-Inflammatory Assays
2.8. Data Analyses
3. Results
3.1. Antioxidant Assays
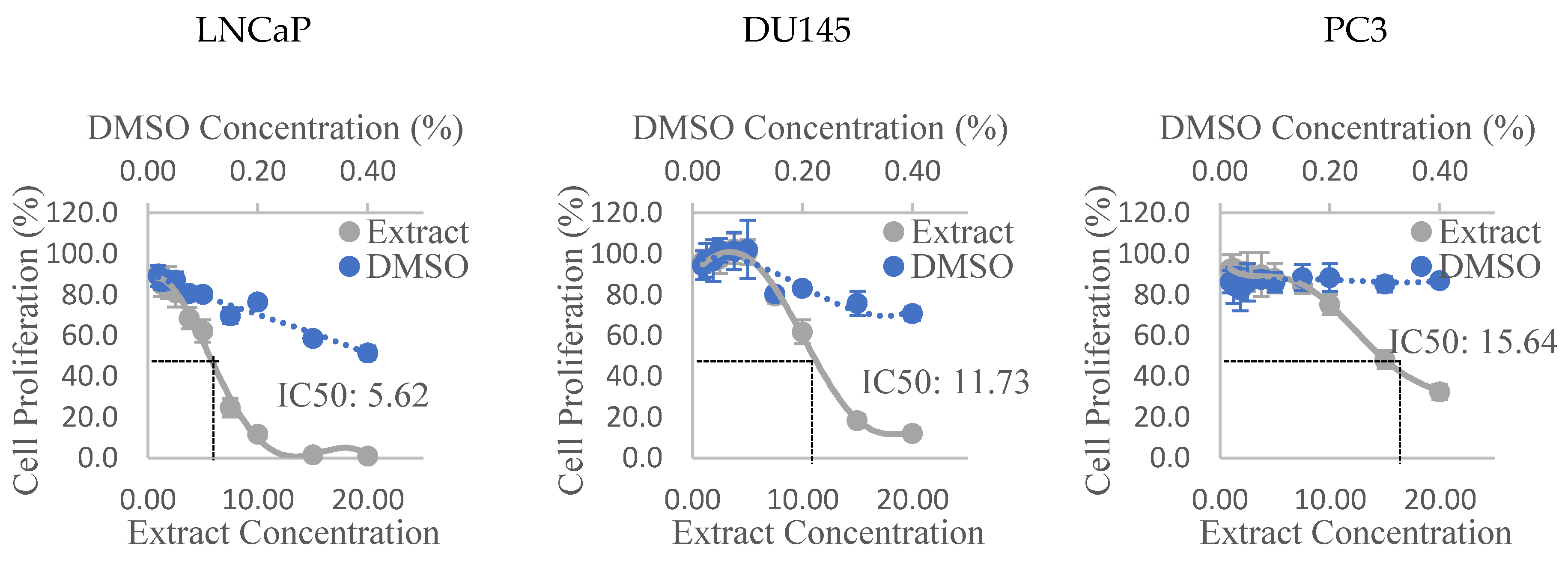
3.2. Anti-Proliferative Activity
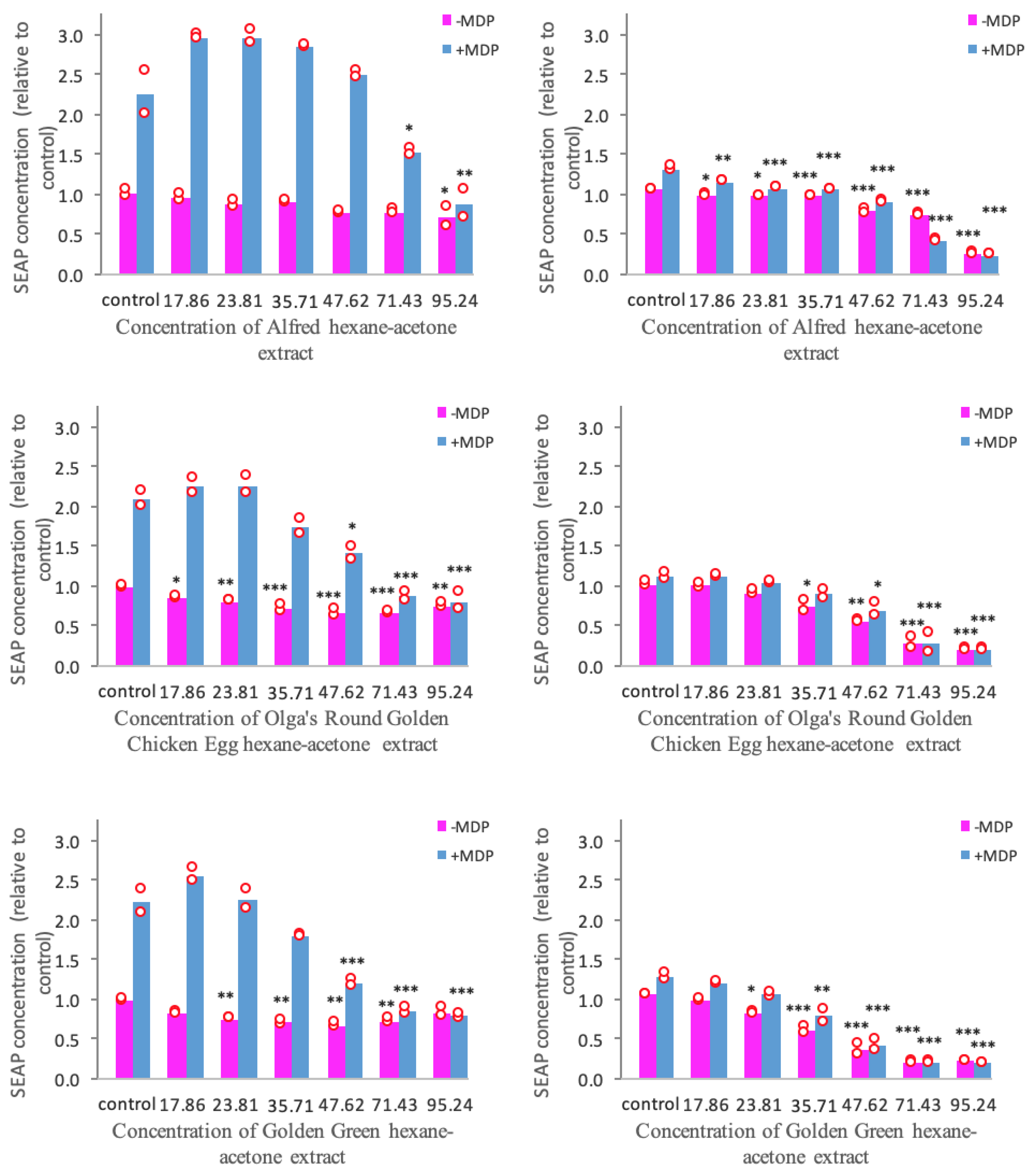
3.3. Anti-Inflammatory Activity Mediated through the TLR2 and TLR4 Pathways
3.4. NOD2-Mediated Anti-Inflammatory Pathway
4. Discussion
4.1. Antioxidant Activity
4.2. Anti-Proliferative Activity
4.3. Anti-Inflammatory Activity
4.3.1. TLR2 and TLR4 Pathways
4.3.2. NOD2 Pathway
4.4. Future Research
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cheng, H.M.; Koutsidis, G.; Lodge, J.K.; Ashor, A.; Siervo, M.; Lara, J. Tomato and lycopene supplementation and cardiovascular risk factors: A systematic review and meta-analysis. Atherosclerosis 2017, 257, 100–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowles, J.L.; Ranard, K.M.; Applegate, C.C.; Jeon, S.; An, R.; Erdman, J.W. Processed and raw tomato consumption and risk of prostate cancer: A systematic review and dose–response meta-analysis. Prostate Cancer Prostatic Dis. 2018, 21, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Yang, X.; Wang, X.; Wang, Y.; Song, Z. The role of tomato products and lycopene in the prevention of gastric cancer: A meta-analysis of epidemiologic studies. Med. Hypotheses 2013, 80, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, J. Meta-analysis of the association between dietary lycopene intake and ovarian cancer risk in postmenopausal women. Sci. Rep. 2014, 4, 4885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.M.; Koutsidis, G.; Lodge, J.K.; Ashor, A.W.; Siervo, M.; Lara, J. Lycopene and tomato and risk of cardiovascular diseases: A systematic review and meta-analysis of epidemiological evidence. Crit. Rev. Food Sci. Nutr. 2019, 59, 141–158. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Zhang, W.; Wang, X.; Zhao, K.; Negi, D.S.; Zhuo, L.; Qi, M.; Wang, X.; Zhang, X. Lycopene and risk of prostate cancer: A systematic review and meta-analysis. Medicine 2015, 94. [Google Scholar] [CrossRef] [PubMed]
- Cooperstone, J.L.; Ralston, R.A.; Riedl, K.M.; Haufe, T.C.; Schweiggert, R.M.; King, S.A.; Timmers, C.D.; Francis, D.M.; Lesinski, G.B.; Clinton, S.K. Enhanced bioavailability of lycopene when consumed as cis-isomers from tangerine compared to red tomato juice, a randomized, cross-over clinical trial. Mol. Nutr. Food Res. 2015, 59, 658–669. [Google Scholar] [CrossRef]
- Unlu, N.Z.; Bohn, T.; Francis, D.; Clinton, S.K.; Schwartz, S.J. Carotenoid absorption in humans consuming tomato sauces obtained from tangerine or high-β-carotene varieties of tomatoes. J. Agric. Food Chem. 2007, 55, 1597–1603. [Google Scholar] [CrossRef]
- Gärtner, C.; Stahl, W.; Sies, H. Lycopene is more bioavailable from tomato paste than from fresh tomatoes. Am. J. Clin. Nutr. 1997, 66, 116–122. [Google Scholar] [CrossRef]
- Boileau, T.W.M.; Boileau, A.C.; Erdman, J.W., Jr. Bioavailability of all-trans and cis–Isomers of Lycopene. Exp. Biol. Med. 2002, 227, 914–919. [Google Scholar] [CrossRef]
- Wang, X.D. Lycopene metabolism and its biological significance. Am. J. Clin. Nutr. 2012, 96, 1214S–1222S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaacson, T.; Ronen, G.; Zamir, D.; Hirschberg, J. Cloning of tangerine from tomato reveals a carotenoid isomerase essential for the production of β-carotene and xanthophylls in plants. Plant Cell 2002, 14, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Wertz, K.; Siler, U.; Goralczyk, R. Lycopene: Modes of action to promote prostate health. Arch. Biochem. Biophys. 2004, 430, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Health. New Cancer Registrations 2016. Available online: https://www.health.govt.nz/publication/new-cancer-registrations-2016 (accessed on 27 May 2019).
- Hwang, E.S.; Bowen, P.E. Effects of tomato paste extracts on cell proliferation, cell-cycle arrest and apoptosis in LNCaP human prostate cancer cells. Biofactors 2005, 23, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Waseem, Z.; Agarwal, S. Lycopene content of tomatoes and tomato products and their contribution to dietary lycopene. Food Res. Int. 1998, 31, 737–741. [Google Scholar] [CrossRef]
- Clough, J.M.; Pattenden, G. Stereochemical assignment of prolycopene and other poly-Z-isomeric carotenoids in fruits of the tangerine tomato Lycopersicon esculentum var.‘Tangella’. J. Chem. Soc. Perkin Trans. 1 1983, 3011–3018. [Google Scholar] [CrossRef]
- Taungbodhitham, A.K.; Jones, G.P.; Wahlqvist, M.L.; Briggs, D.R. Evaluation of extraction method for the analysis of carotenoids in fruits and vegetables. Food Chem. 1998, 63, 577–584. [Google Scholar] [CrossRef]
- Masters, J.R.; Thomson, J.A.; Daly-Burns, B.; Reid, Y.A.; Dirks, W.G.; Packer, P.; Toji, L.H.; Ohno, T.; Tanabe, H.; Arlett, C.F. Short tandem repeat profiling provides an international reference standard for human cell lines. Proc. Natl. Acad. Sci. USA 2001, 98, 8012–8017. [Google Scholar] [CrossRef] [Green Version]
- Philpott, M.; Mackay, L.; Ferguson, L.R.; Forbes, D.; Skinner, M. Cell culture models in developing nutrigenomics foods for inflammatory bowel disease. Mutat. Res./Fundam. Mol. Mech. Mutagenes. 2007, 622, 94–102. [Google Scholar] [CrossRef]
- Folkard, D.L.; Marlow, G.; Mithen, R.F.; Ferguson, L.R. Effect of Sulforaphane on NOD2 via NF-κB: Implications for Crohn’s disease. J. Inflamm. 2015, 12, 6. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Miller, N.J.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef]
- Ozgen, M.; Reese, R.N.; Tulio, A.Z.; Scheerens, J.C.; Miller, A.R. Modified 2, 2-azino-bis-3-ethylbenzothiazoline-6-sulfonic acid (ABTS) method to measure antioxidant capacity of selected small fruits and comparison to ferric reducing antioxidant power (FRAP) and 2, 2 ‘-diphenyl-1-picrylhydrazyl (DPPH) methods. J. Agric. Food Chem. 2006, 54, 1151–1157. [Google Scholar] [CrossRef]
- Müller, L.; Goupy, P.; Fröhlich, K.; Dangles, O.; Caris-Veyrat, C.; Böhm, V. Comparative study on antioxidant activity of lycopene (Z)-isomers in different assays. J. Agric. Food Chem. 2011, 59, 4504–4511. [Google Scholar] [CrossRef]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxicity assay for anticancer-drug screening. JNCI J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef]
- Tomblin, V.; Ferguson, L.R.; Han, D.Y.; Murray, P.; Schlothauer, R. Potential pathway of anti-inflammatory effect by New Zealand honeys. Int. J. Gen. Med. 2014, 7, 149. [Google Scholar] [CrossRef]
- Peng, Y.; Bishop, K.; Ferguson, L.; Quek, S. Screening of Cytotoxicity and Anti-Inflammatory Properties of Feijoa Extracts Using Genetically Modified Cell Models Targeting TLR2, TLR4 and NOD2 Pathways, and the Implication for Inflammatory Bowel Disease. Nutrients 2018, 10, 1188. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Routledge: New York, NY, USA, 1988; pp. 66–67. [Google Scholar]
- Apak, R.; Gorinstein, S.; Böhm, V.; Schaich, K.M.; Özyürek, M.; Güçlü, K. Methods of measurement and evaluation of natural antioxidant capacity/activity (IUPAC Technical Report). Pure Appl. Chem. 2013, 85, 957–998. [Google Scholar] [CrossRef] [Green Version]
- AOAC. AOAC SMPR 2011.011 Standard Method Performance Requirements for In Vitro Determination of Total Antioxidant Activity. Available online: http://www.aoac.org/aoac_prod_imis/AOAC_Docs/OMA/OMAChangesinMethods_050913.pdf (accessed on 27 May 2019).
- Schaich, K.M.; Tian, X.; Xie, J. Reprint of “Hurdles and pitfalls in measuring antioxidant efficacy: A critical evaluation of ABTS, DPPH, and ORAC assays”. J. Funct. Foods 2015, 18, 782–796. [Google Scholar] [CrossRef]
- Müller, L.; Fröhlich, K.; Böhm, V. Comparative antioxidant activities of carotenoids measured by ferric reducing antioxidant power (FRAP), ABTS bleaching assay (αTEAC), DPPH assay and peroxyl radical scavenging assay. Food Chem. 2011, 129, 139–148. [Google Scholar] [CrossRef]
- McGhie, T.K.; Cordiner, S. Carotenoid Composition of Tomatoes. Available online: https://www.heritagefoodcrops.org.nz/static/media/2015-carotenoid-composition-of-tomatoes.9d149f68.pdf (accessed on 27 May 2019).
- Kotake-Nara, E.; Kushiro, M.; Zhang, H.; Sugawara, T.; Miyashita, K.; Nagao, A. Carotenoids affect proliferation of human prostate cancer cells. J. Nutr. 2001, 131, 3303–3306. [Google Scholar] [CrossRef]
- Williams, A.W.; Boileau, T.W.-M.; Zhou, J.R.; Clinton, S.K.; Erdman, J.W., Jr. β-Carotene modulates human prostate cancer cell growth and may undergo intracellular metabolism to retinol. J. Nutr. 2000, 130, 728–732. [Google Scholar] [CrossRef]
- Dulińska, J.; Gil, D.; Zagajewski, J.; Hartwich, J.; Bodzioch, M.; Dembińska-Kieć, A.; Langmann, T.; Schmitz, G.; Laidler, P. Different effect of beta-carotene on proliferation of prostate cancer cells. Biochimica Biophysica Acta (BBA)-Mol. Basis Dis. 2005, 1740, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Bank, S.; Andersen, P.S.; Burisch, J.; Pedersen, N.; Roug, S.; Galsgaard, J.; Turino, S.Y.; Brodersen, J.B.; Rashid, S.; Rasmussen, B.K. Polymorphisms in the inflammatory pathway genes TLR2, TLR4, TLR9, LY96, NFKBIA, NFKB1, TNFA, TNFRSF1A, IL6R, IL10, IL23R, PTPN22, and PPARG are associated with susceptibility of inflammatory bowel disease in a Danish cohort. PLoS ONE 2014, 9, e98815. [Google Scholar] [CrossRef]
- Jostins, L.; Ripke, S.; Weersma, R.K.; Duerr, R.H.; McGovern, D.P.; Hui, K.Y.; Lee, J.C.; Schumm, L.P.; Sharma, Y.; Anderson, C.A. Host–microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature 2012, 491, 119. [Google Scholar] [CrossRef]
- Curkovic, I.; Egbring, M.; Kullak-Ublick, G.A. Risks of inflammatory bowel disease treatment with glucocorticosteroids and aminosalicylates. Dig. Dis. 2013, 31, 368–373. [Google Scholar] [CrossRef]
- Hou, J.K.; Lee, D.; Lewis, J. Diet and inflammatory bowel disease: Review of patient-targeted recommendations. Clin. Gastroenterol. Hepatol. 2014, 12, 1592–1600. [Google Scholar] [CrossRef]
- Hartman, C.; Eliakim, R.; Shamir, R. Nutritional status and nutritional therapy in inflammatory bowel diseases. World J. Gastroenterol. WJG 2009, 15, 2570. [Google Scholar] [CrossRef]
- Marcotorchino, J.; Romier, B.; Gouranton, E.; Riollet, C.; Gleize, B.; Malezet-Desmoulins, C.; Landrier, J.F. Lycopene attenuates LPS-induced TNF-α secretion in macrophages and inflammatory markers in adipocytes exposed to macrophage-conditioned media. Mol. Nutr. Food Res. 2012, 56, 725–732. [Google Scholar] [CrossRef]
- Riso, P.; Visioli, F.; Grande, S.; Guarnieri, S.; Gardana, C.; Simonetti, P.; Porrini, M. Effect of a tomato-based drink on markers of inflammation, immunomodulation, and oxidative stress. J. Agric. Food Chem. 2006, 54, 2563–2566. [Google Scholar] [CrossRef]
- Maekawa, S.; Ohto, U.; Shibata, T.; Miyake, K.; Shimizu, T. Crystal structure of NOD2 and its implications in human disease. Nat. Commun. 2016, 7, 11813. [Google Scholar] [CrossRef] [Green Version]
- Cooperstone, J.L.; Francis, D.M.; Schwartz, S.J. Thermal processing differentially affects lycopene and other carotenoids in cis-lycopene containing, tangerine tomatoes. Food Chem. 2016, 210, 466–472. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Solvent | Ratio | Source |
|---|---|---|
| Hexane | - | [17] |
| Hexane:Acetone:Ethanol | 2:1:1 | [18] |
| Hexane:Acetone | 1:1 | [19] |
| Hexane:Ethanol | 3:4 | [20] |
| Extraction Solvent | Tomato Variety | |||
|---|---|---|---|---|
| Alfred | Olga’s Round Golden Chicken Egg | Golden Green | Golden Eye | |
| FRAP Assay | ||||
| Hexane | 11.71 ± 0.62 d/B | 6.24 ± 0.30 c/A | 12.52 ± 0.54 c/B | 12.50 ± 0.35 d/B |
| HAE | 3.66 ± 0.12 b | 2.23 ± 0.21 a | 5.99 ± 0.31 a | 6.18 ± 0.34 b |
| HA | 7.10 ± 0.45 c | 4.72 ± 0.36 b | 10.54 ± 0.17 b | 8.60 ± 0.54 c |
| HE | 2.11 ± 0.22 a | 1.79 ± 0.22 a | 5.58 ± 0.25 a | 3.23 ± 0.23 a |
| ABTS Assay | ||||
| Hexane | 20.04 ± 1.40 d/B | 14.27 ± 1.03 d/A | 26.22 ± 0.32 c/C | 18.32 ± 1.16 d/B |
| HAE | 6.23 ± 0.50 b | 6.82 ± 0.64 b | 14.42 ± 1.06 a | 8.24 ± 0.70 b |
| HA | 13.84 ± 0.33 c | 11.46 ± 1.16 c | 19.76 ± 1.60 b | 11.87 ± 0.69 c |
| HE | 1.99 ± 0.20 a | 6.11 ± 0.44 a | 13.42 ± 1.04 a | 3.77 ± 0.26 a |
| Tomato Variety | Extraction Solvent | |||
|---|---|---|---|---|
| Hexane | HAE | HA | HE | |
| LNCaP | ||||
| Alfred | 14.46 | X | 11.76† | X |
| Olga’s | 5.62 † | 16.93 | 10.1 | X |
| Golden Green | 8.08 † | 15.45 | 10.19 | X |
| Golden Eye | + | + | + | + |
| DU145 | ||||
| Alfred | X | X | -† | X |
| Olga’s | 11.73 † | X | - | X |
| Golden Green | 17.84 † | X | 18.01 | X |
| Golden Eye | + | + | + | + |
| PC3 | ||||
| Alfred | X | X | -† | X |
| Olga’s | 15.64 † | X | - | X |
| Golden Green | -† | X | X | X |
| Golden Eye | + | + | + | + |
| - | Concentration (mg Dry Tomato Equivalent/mL) | Ibuprofen (mg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Alfred Hexane | Alfred HA | Olga’s Hexane | Olga’s HA | GG Hexane | GG HA | GE Hexane | GE HA | ||
| HEK-Blue™ hTLR2 | |||||||||
| IC30 | 60.24 | 59.35 | 53.69 * | 39.96 | 50.01 * | 32.83 | N/A | N/A | 0.27 |
| IC50 | 88.46 | 74.73 | 66.22 * | 55.62 | 61.16 * | 50.41 | N/A | N/A | 0.4 |
| IC70 | N/A | 90.1 | 78.75 * | 71.28 | 72.3 * | 67.99 | N/A | N/A | N/A |
| HEK-Blue™ hTLR4 | |||||||||
| IC30 | 84.56 * | 64.47 | 52.2 * | 51.51 | 56.69 * | 42.76 | N/A | N/A | 0.15 |
| IC50 | N/A | 76.01 | 61.44 * | 62.53 | 62.89 * | 53.08 | N/A | N/A | 0.27 |
| IC70 | N/A | 87.55 | 70.68 * | 73.55 | 69.09 * | 63.4 | N/A | N/A | 0.38 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanambell, H.; Quek, S.Y.; Bishop, K.S. Screening of In Vitro Health Benefits of Tangerine Tomatoes. Antioxidants 2019, 8, 230. https://doi.org/10.3390/antiox8070230
Tanambell H, Quek SY, Bishop KS. Screening of In Vitro Health Benefits of Tangerine Tomatoes. Antioxidants. 2019; 8(7):230. https://doi.org/10.3390/antiox8070230
Chicago/Turabian StyleTanambell, Hartono, Siew Young Quek, and Karen Suzanne Bishop. 2019. "Screening of In Vitro Health Benefits of Tangerine Tomatoes" Antioxidants 8, no. 7: 230. https://doi.org/10.3390/antiox8070230
APA StyleTanambell, H., Quek, S. Y., & Bishop, K. S. (2019). Screening of In Vitro Health Benefits of Tangerine Tomatoes. Antioxidants, 8(7), 230. https://doi.org/10.3390/antiox8070230