Panax ginseng Fruit Has Anti-Inflammatory Effect and Induces Osteogenic Differentiation by Regulating Nrf2/HO-1 Signaling Pathway in In Vitro and In Vivo Models of Periodontitis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Materials
2.3. Cell Culture
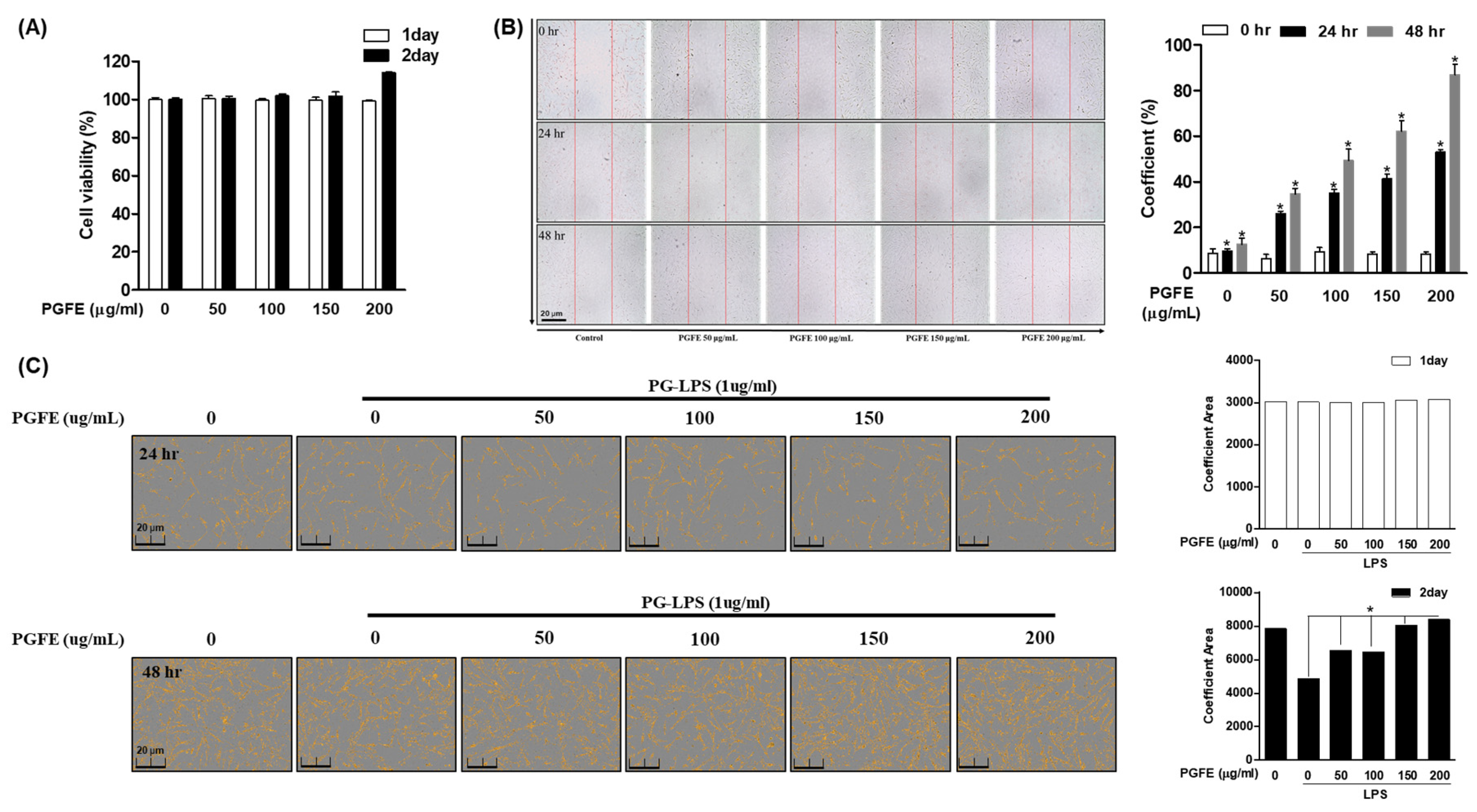
2.4. Cell Viability and Coefficient Assays
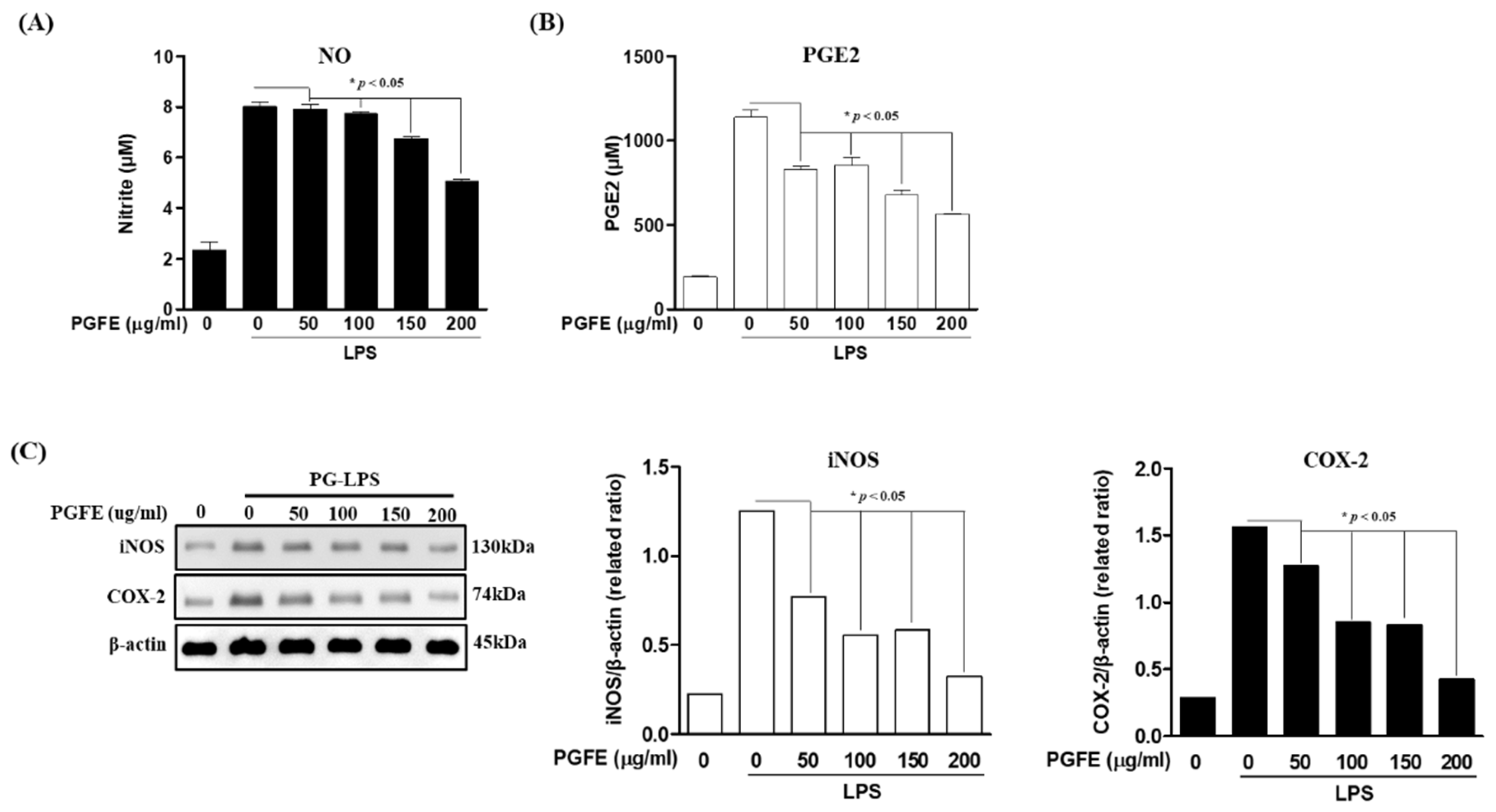
2.5. Nitrite Assay Assays
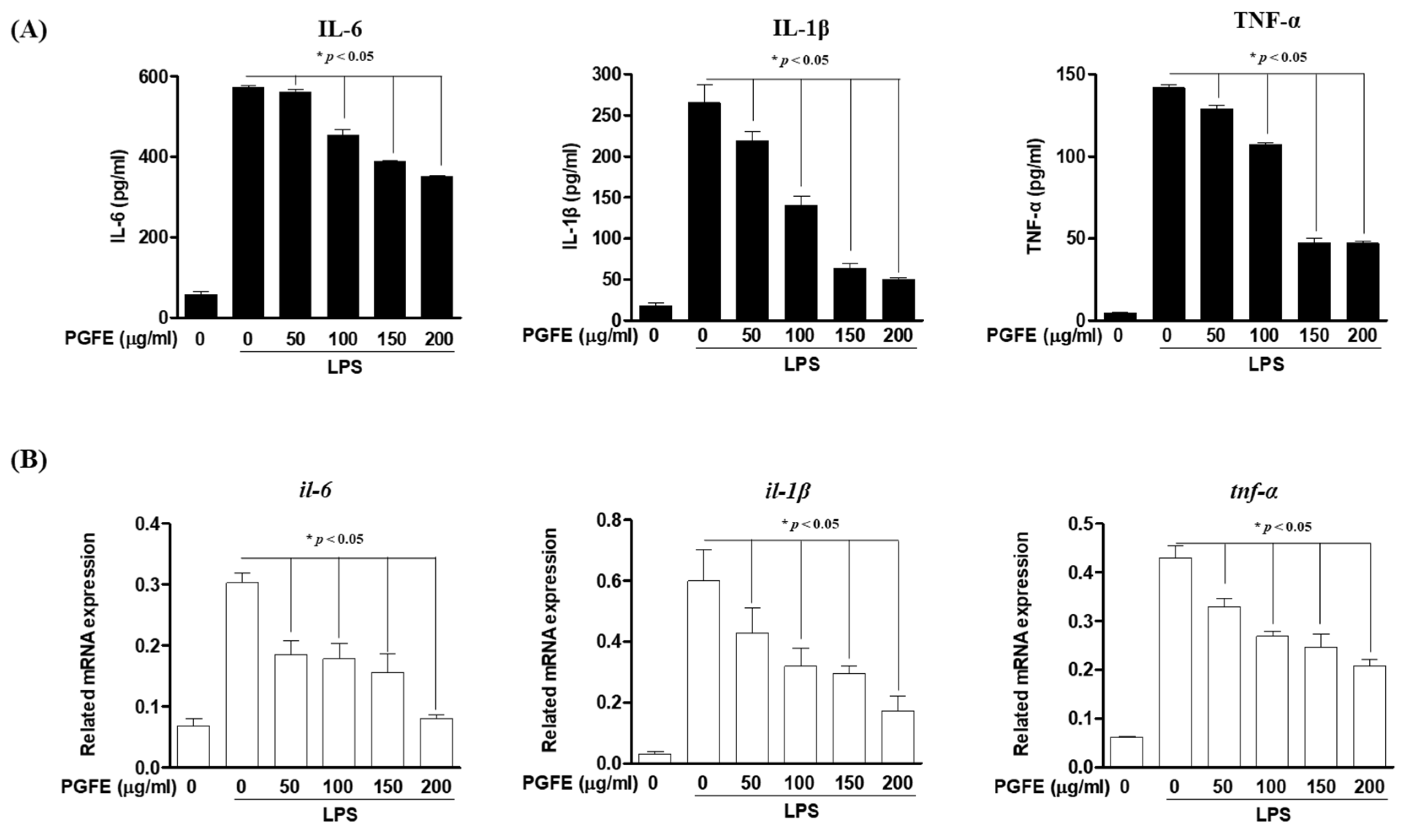
2.6. Cytokines Production Assay
2.7. Wound-Healing Assays
2.8. Mineralization Assay
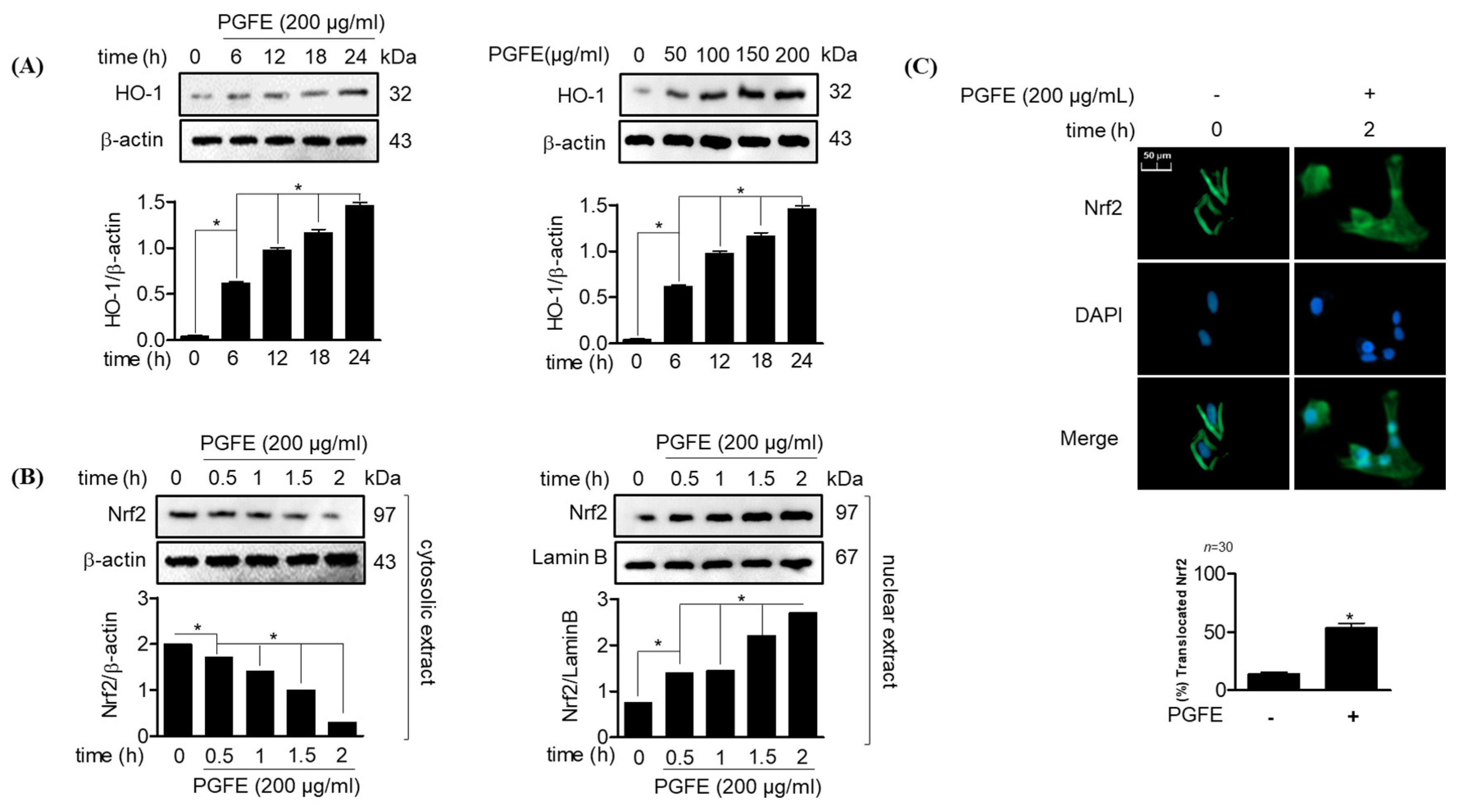
2.9. Immunofluorescence Analysis
2.10. Cytosolic and Nuclear Protein Extraction
2.11. Western Blot Analysis
2.12. RT-qPCR Analysis
2.13. Animal
2.14. Ligature-Induced Periodontitis Model
2.15. PG-LPS-Induced Periodontitis Model
2.16. Micro-CT Imaging and Analysis
2.17. Histological Staining
2.18. Statistical Analysis
3. Results
3.1. P. ginseng Fruit Extract (PGFE) Is Not Cytotoxic and Promotes HPDL Cell Proliferation
3.2. PGFE Promotes HO-1 Expression and Nrf2 Translocation in HPDL Cells
3.3. Inhibitory Effects of PGFE on PG-LPS-Induced Expression of Pro-Inflammatory Cytokines and Mediators in HPDL Cells
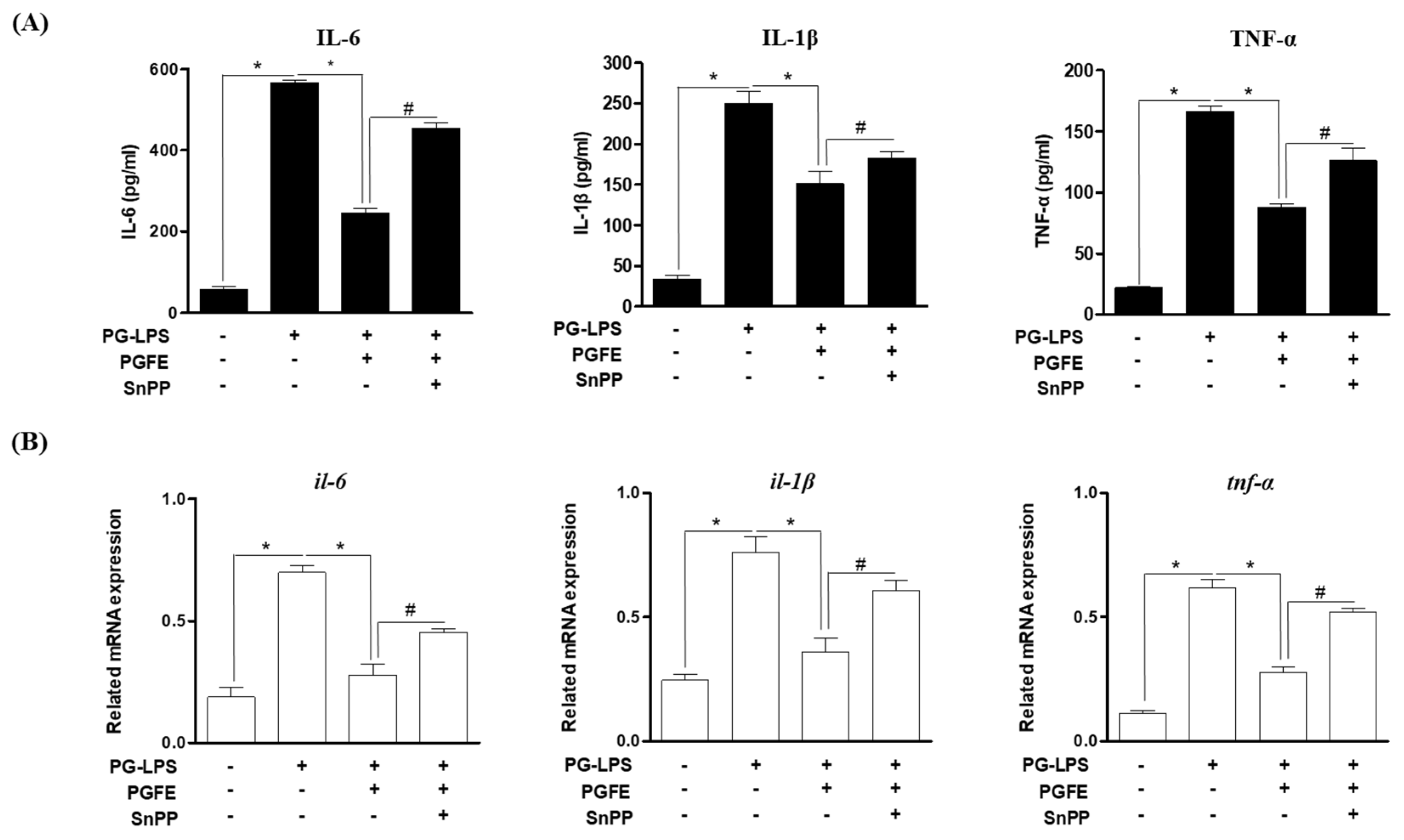
3.4. PGFE Inhibits PG-LPS-Induced Pro-Inflammatory Cytokines by Promoting HO-1 Expression
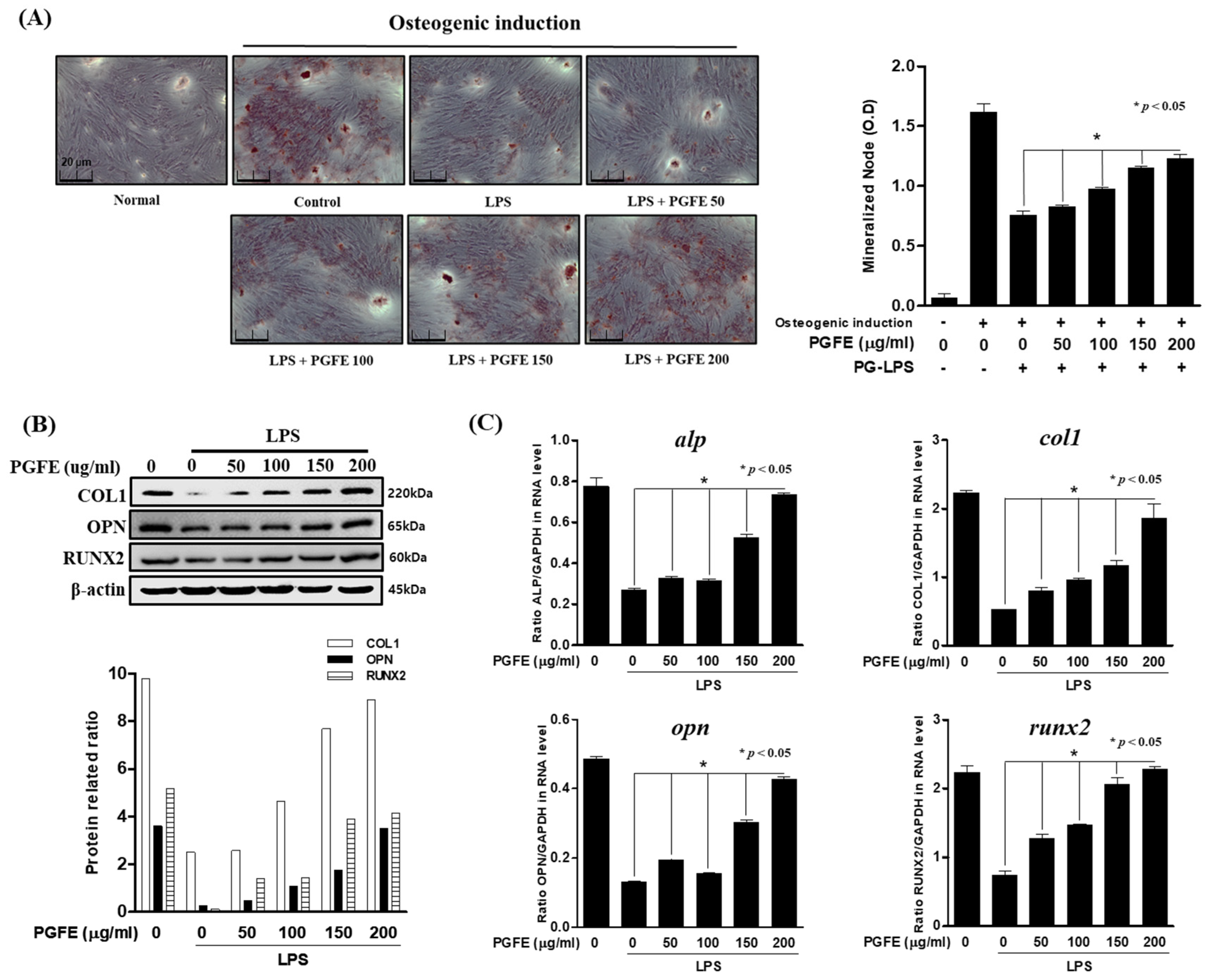
3.5. Osteogenic Induction of HPDL Cells by PGFE
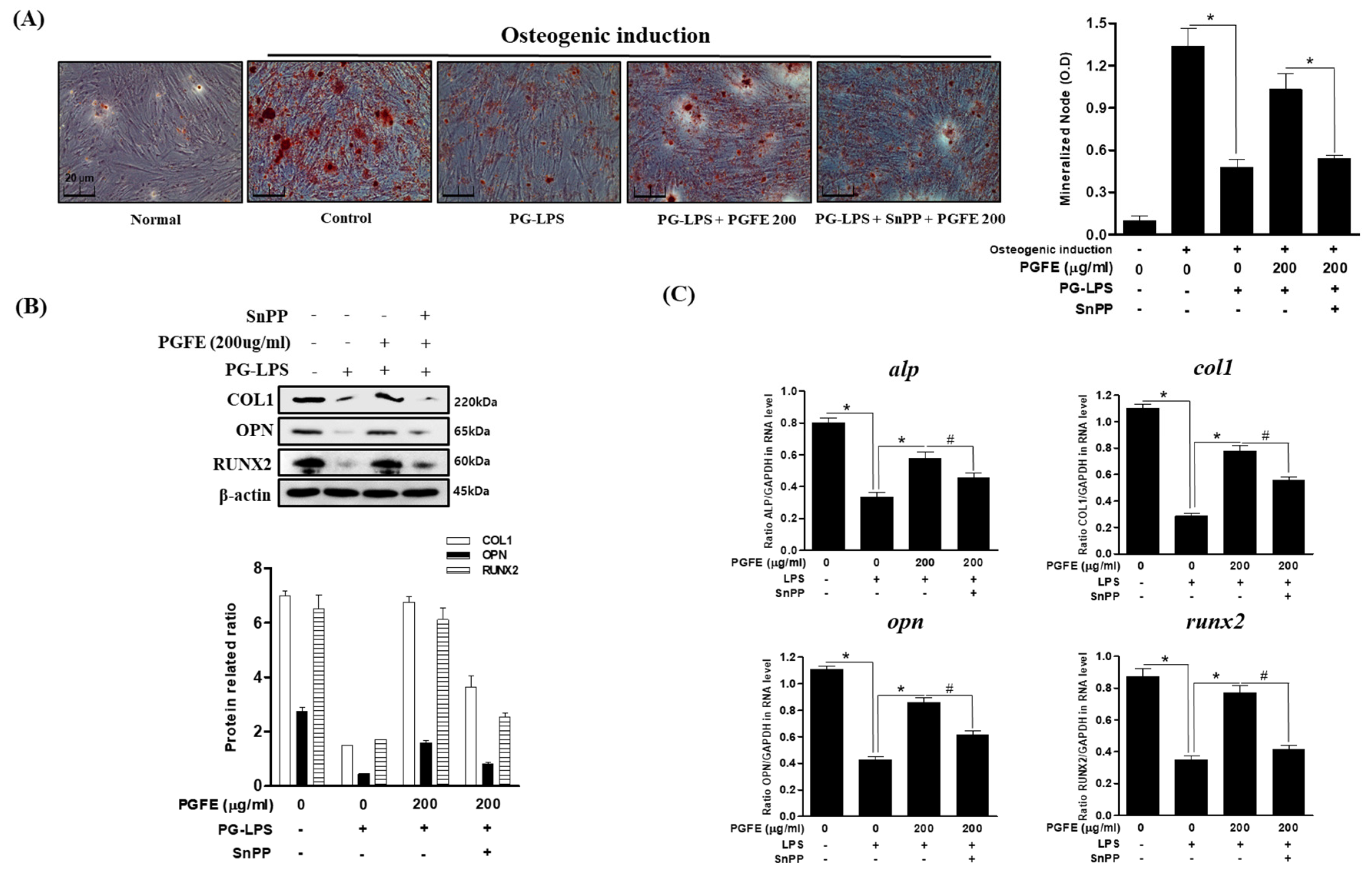
3.6. Effect of PGFE-Induced HO-1 Expression on Osteogenic Differentiation
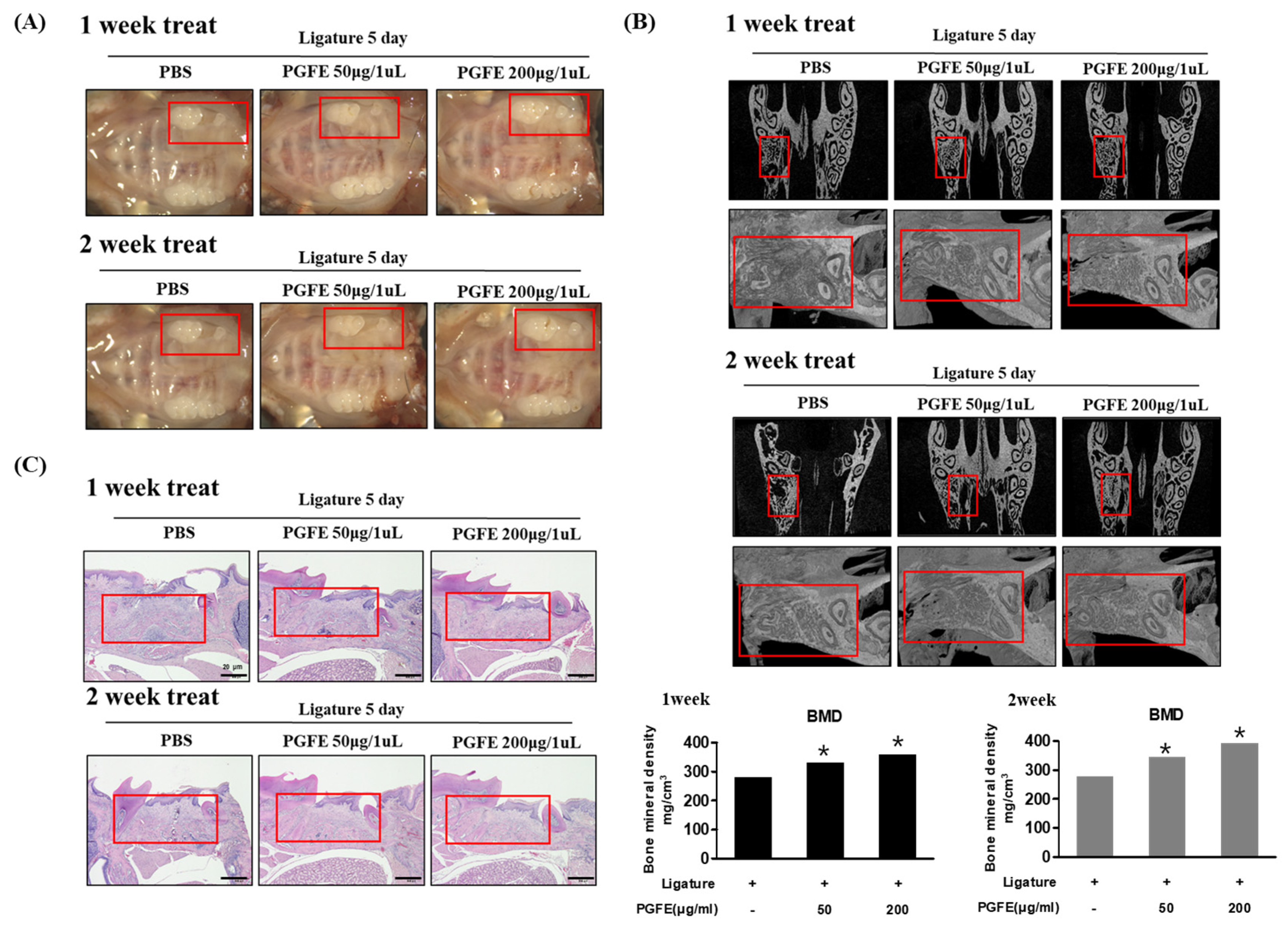
3.7. Inhibitory Effect of PGFE on Periodontitis in a Ligature-Induced In Vivo Model
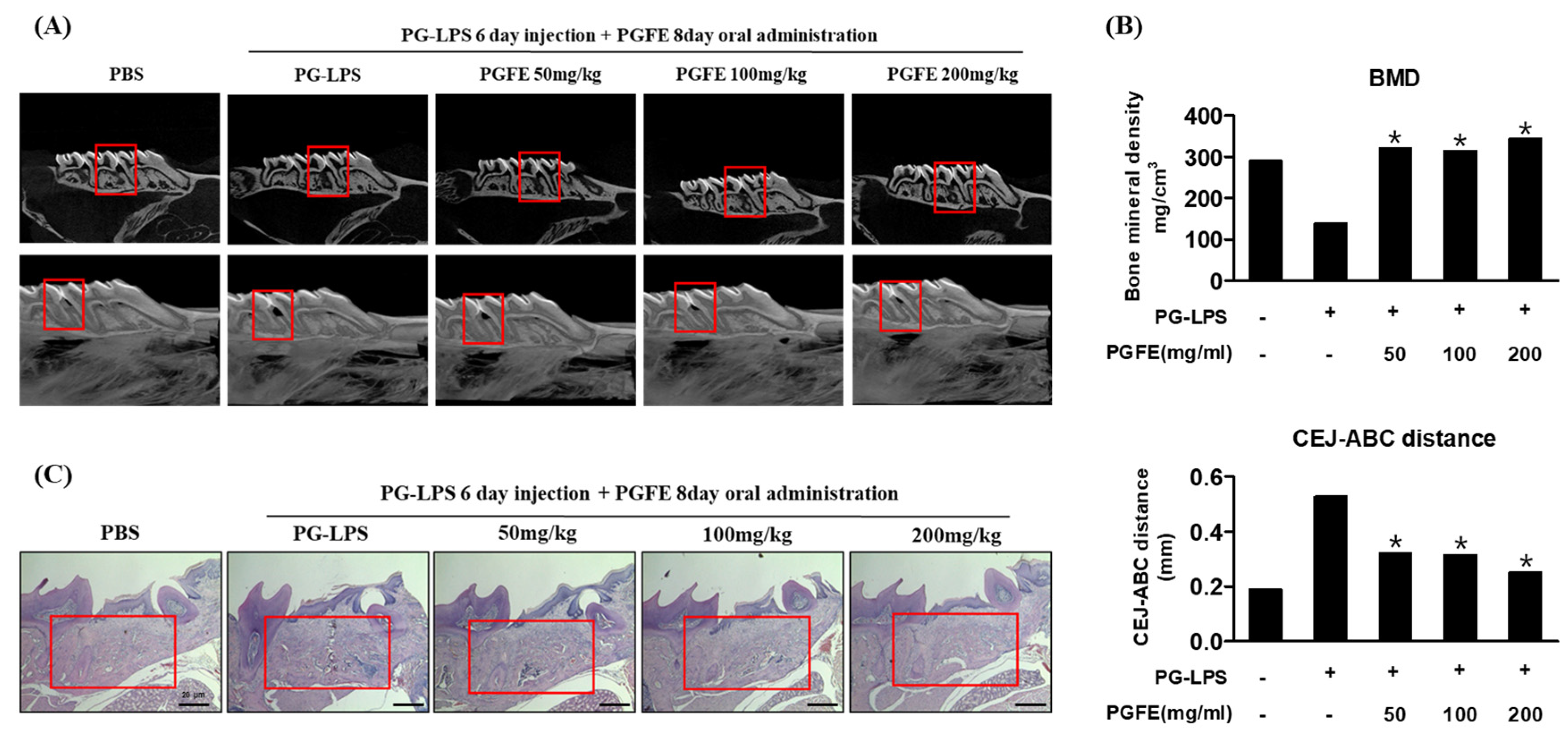
3.8. Inhibitory Effect of PGFE on Periodontitis in PG-LPS Induced Periodontitis In Vivo Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Profio, B.D.; Villar, C.C.; Saraiva, L.; Ortega, L.K.; Pannuti, C.M. Is periodontitis a risk factor for infections in cirrhotic patients. Med. Hypotheses 2017, 106, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Genco, R.J. Current view of risk factors for periodontal diseases. J. Periodontol. 1996, 67, 1041–1049. [Google Scholar] [PubMed]
- Pihlstrom, B.L.; Michalowicz, B.S.; Johnson, N.W. Periodontal diseases. Lancet 2005, 366, 1809–1820. [Google Scholar] [CrossRef] [Green Version]
- Guha, M.; Mackman, N. LPS induction of gene expression in human monocytes. Cell. Signal. 2001, 13, 85–94. [Google Scholar] [CrossRef]
- Kjeldsen, M.; Holmstrup, P.F.; Bendtzen, K. Marginal Periodontitis and Cytokines: A Review of the Literature. J. Periodontol. 1993, 64, 1013–1022. [Google Scholar] [CrossRef]
- Maruyama, T.; Tomofuji, T.; Endo, Y.; Irie, K. Supplementation of green tea catechins in dentifrices suppresses gingival oxidative stress and periodontal inflammation. Arch. Oral Biol. 2011, 56, 48–53. [Google Scholar] [CrossRef]
- Shao, M.Y.; Huang, P.; Cheng, R.; Hu, T. Interleukin-6 polymorphisms modify the risk of periodontitis: A systematic review and meta-analysis. J. Zhejiang Univ. Sci. B 2009, 10, 920–927. [Google Scholar] [CrossRef] [Green Version]
- Hayami, T.; Zhang, Q.; Kapila, Y.; Kapila, S. Dexamethasone’s enhancement of osteoblastic markers in human periodontal ligament cells is associated with inhibition of collagenase expression. Bone 2007, 40, 93–104. [Google Scholar] [CrossRef]
- Bao, T.H.L.; Doan, V.N.; Le, H.T.N.; Ngo, L.T.Q. Various methods for isolation of multipotent human periodontal ligament cells for regenerative medicine. In Vitro Cell. Dev. Biol. Anim. 2014, 50, 597–602. [Google Scholar]
- Ari, G.; Cherukuri, S.; Namasivayam, A. Epigenetics and Periodontitis: A Contemporary Review. J. Clin. Diagn. Res. 2016, 10, ZE07–ZE09. [Google Scholar] [CrossRef]
- Bezerra, M.M.; Lima, V.; Alencar, V.B.; Vieira, I.B.; Brito, G.A.; Ribeiro, R.A.; Rocha, F.A. Selective cyclooxygenase-2 inhibition prevents alveolar bone loss in experimental periodontitis in rats. J. Periodontol. 2000, 71, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Otterbein, L.E.; Choi, A.M. Heme oxygenase: Colors of defense against cellular stress. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, 1029–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Davies, K.J.A.; Forman, H.J. Oxidative stress response and Nrf2 signaling in aging. Free Radic. Biol. Med. 2015, 88, 314–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.; Kook, S.H.; Ji, H.; Lee, S.A.; Choi, K.C.; Lee, K.Y.; Lee, J.C. N-acetyl cysteine inhibits H2O2-mediated reduction in the mineralization of MC3T3-E1 cells by down-regulating Nrf2/HO-1 pathway. BMB Rep. 2015, 48, 636–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.M.; Suh, K.S.; Kim, Y.J.; Hong, S.M.; Park, S.Y.; Chon, S. Glabridin alleviates the toxic effects of Methylglyoxal on osteoblastic MC3T3-E1 cells by increasing expression of the glyoxalase system and Nrf2/HO-1 signaling and protecting mitochondrial function. J. Agric. Food Chem. 2016, 64, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, D.H.; Park, S.J.; Kim, J.M.; Ryu, J.H. Ginseng in traditional herbal prescriptions. J. Ginseng Res. 2012, 36, 225–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.R.; Xiang, Z.J.; Ye, T.X.; Yuan, Y.J.; Guo, Z.X. Antioxidant activities of Salvia miltiorrhiza and Panax notoginseng. Food Chem. 2006, 99, 767–774. [Google Scholar] [CrossRef]
- Lee, I.A.; Hyam, S.R.; Jang, S.E.; Han, M.J.; Kim, D.H. Ginsenoside Re ameliorates inflammation by inhibiting the binding of lipopolysaccharide to TLR4 on macrophages. J. Agric. Food Chem. 2012, 60, 9595–9602. [Google Scholar] [CrossRef]
- Seo, B.M.; Miura, M.; Gronthos, S.; Bartold, P.M.; Batouli, S.; Brahim, J.; Young, M.; Robey, P.G.; Wang, C.Y.; Shi, S. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef]
- Assuma, R.; Oates, T.; Cochran, D.; Amar, S.; Graves, D.T. IL-1 and TNF antagonists inhibit the inflammatory response and bone loss in experimental periodontitis. J. Immunol. 1998, 160, 403–409. [Google Scholar]
- Zhu, W.; Liang, M. Periodontal ligament stem cells: Current status, concerns, and future prospects. Stem Cells Int. 2015, 972313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassir, S.H.; Wisitrasameewong, W.; Raanan, J.; Ghaffarigarakani, S.; Chung, J.; Freire, M.; Andrada, L.C.; Intini, G. Potential for stem cell-based periodontal therapy. J. Cell. Physiol. 2016, 231, 50–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, Y.; Yashiro, Y.; Sanggarnjanavanich, S.; Yamaguchi, T.; Arai, C.; Noda, K.; Takano, Y.; Nakamura, Y.; Hanada, N. Human periodontal ligament fibroblasts are the optimal cell source for induced pluripotent stem cells. Histochem. Cell Biol. 2012, 137, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Otterbein, L.E.; Soares, M.P.; Yamashita, K.; Bach, F.H. Heme oxygenase-1: Unleashing the protective properties of heme. Trends Immunol. 2003, 24, 449–455. [Google Scholar] [CrossRef]
- Page, R.C. The role of inflammatory mediators in the pathogenesis of periodontal disease. J. Periodontal. Res. 1991, 26, 230–342. [Google Scholar] [CrossRef]
- Jeong, G.S.; Lee, D.S.; Li, B.; Kim, J.J.; Kim, E.C.; Kim, Y.C. Anti-inflammatory effects of lindenenyl acetate via heme oxygenase-1 and AMPK in human periodontal ligament cells. Eur. J. Pharmacol. 2011, 670, 295–303. [Google Scholar] [CrossRef]
- Takanche, J.S.; Kim, J.E.; Han, S.H.; Yi, H.K. Effect of gomisin A on osteoblast differentiation in high glucose-mediated oxidative stress. Phytomedicine 2020, 66, 153107. [Google Scholar] [CrossRef]
- Ram, V.S.; Sudhakar, U.P. Bone biomarkers in periodontal disease: A review article. J. Clin. Diagn. Res. 2015, 9, 7–10. [Google Scholar]
- Enomoto, H.; Furuichi, T.; Zanma, A. Runx2 deficiency in chondrocytes causes adipogenic changes in vitro. J. Cell Sci. 2004, 117, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Im, G.Y.; Ma, J.Y.; Kim, K.W.; Choi, J.K.; Kang, D.K.; Kwon, T.R.; Jang, S.Y.; Jeong, Y.J. Quality characteristics of 4 year-old ginseng by enzymatic hydrolysis conditions. J. Korean Soc. Food Sci. Nutr. 2011, 40, 229–234. [Google Scholar] [CrossRef]
- Kim, N.M.; Lee, J.S.; Lee, B.H. Enzymatic hydrolysis of korean ginseng starch and characteristics of produced maltooligosaccharides. J. Ginseng Res. 2000, 24, 41–45. [Google Scholar]
- Kook, K.E.; Kim, C.; Kang, W.; Hwang, J.K. Inhibitory Effect of Standardized Curcuma xanthorrhiza Supercritical Extract on LPS-Induced Periodontitis in Rats. J. Microbiol. Biotechnol. 2018, 28, 1614–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.G.; Kim, M.O.; Kim, S.H.; Kim, H.J.; Pokhrel, N.K.; Lee, J.H.; Lee, H.J.; Kim, J.Y.; Lee, Y. 6-Shogaol, an active ingredient of ginger, inhibits osteoclastogenesis and alveolar bone resorption in ligature-induced periodontitis in mice. J. Periodontol. 2020, 91, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.E.; Hyun, K.Y. Inhibitory effect of Acer tegmentosum maxim extracts on P. gingivalis LPS induced Periodontitis. Arch. Oral Biol. 2020, 109, 104529. [Google Scholar] [CrossRef]
- Lee, B.A.; Lee, H.S.; Jung, Y.S.; Kim, S.W.; Lee, Y.W.; Chang, S.H.; Chung, H.J.; Kim, O.S.; Kim, Y.J. The Effects of a Novel Botanical Agent on Lipopolysaccharide-Induced Alveolar Bone Loss in Rats. J. Periodontol. 2013, 84, 1221–1229. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Sequence (5′→3′) | Accession Number | |
|---|---|---|---|
| il-6 | Forward | AGTGAGGAACAAGCCAGAGC | NM_000600.4 |
| Reverse | GTCAGGGGTGGTTATTGCAT | ||
| il-1β | Forward | AACCTCTTCGAGGCACAAGG | NM_000576.2 |
| Reverse | GTCCTGGAAGGAGCACTTCAT | ||
| tnf-α | Forward | GCCTCTTCTCCTTCCTGATCGT | NM_000594.2 |
| Reverse | TGAGGGTTTGCTACAACATGGG | ||
| alp | Forward | TGCAGTACGAGCTGAACAGG | NM_000478 |
| Reverse | GTCAATTCTGCCTCCTTCCA | ||
| col1 | Forward | CCAGAAGAACTGGTACATCAGCAA | NM_000088 |
| Reverse | CGCCATACTCGAACTGGAATC | ||
| opn | Forward | TCAGCTGGATGACCAGAGTG | NM_001040060 |
| Reverse | TTGGGGTCTACAACCAGCAT | ||
| runx2 | Forward | TCTTAGAACAAATTCTGCCCTTT | NM_001024630.3 |
| Reverse | TGCTTTGGTCTTGAAATCACA | ||
| gapdh | Forward | TGTTCGTCATGGGTGTGAAC | NM_002046 |
| Reverse | GTCTTCTGGGTGGCAGTGAT | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.-N.; Kim, T.-Y.; Park, E.K.; Kim, J.-Y.; Jeong, G.-S. Panax ginseng Fruit Has Anti-Inflammatory Effect and Induces Osteogenic Differentiation by Regulating Nrf2/HO-1 Signaling Pathway in In Vitro and In Vivo Models of Periodontitis. Antioxidants 2020, 9, 1221. https://doi.org/10.3390/antiox9121221
Kim E-N, Kim T-Y, Park EK, Kim J-Y, Jeong G-S. Panax ginseng Fruit Has Anti-Inflammatory Effect and Induces Osteogenic Differentiation by Regulating Nrf2/HO-1 Signaling Pathway in In Vitro and In Vivo Models of Periodontitis. Antioxidants. 2020; 9(12):1221. https://doi.org/10.3390/antiox9121221
Chicago/Turabian StyleKim, Eun-Nam, Tae-Young Kim, Eui Kyun Park, Jae-Young Kim, and Gil-Saeng Jeong. 2020. "Panax ginseng Fruit Has Anti-Inflammatory Effect and Induces Osteogenic Differentiation by Regulating Nrf2/HO-1 Signaling Pathway in In Vitro and In Vivo Models of Periodontitis" Antioxidants 9, no. 12: 1221. https://doi.org/10.3390/antiox9121221
APA StyleKim, E. -N., Kim, T. -Y., Park, E. K., Kim, J. -Y., & Jeong, G. -S. (2020). Panax ginseng Fruit Has Anti-Inflammatory Effect and Induces Osteogenic Differentiation by Regulating Nrf2/HO-1 Signaling Pathway in In Vitro and In Vivo Models of Periodontitis. Antioxidants, 9(12), 1221. https://doi.org/10.3390/antiox9121221