Impact of Enzymatic and Microbial Bioprocessing on Antioxidant Properties of Hemp (Cannabis sativa L.)

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Material, Enzymes and Microorganisms
2.2. Bioprocessing
2.3. Microbiological Analysis
2.4. Biochemical Characterization
2.4.1. pH and Organic Acids
2.4.2. Proteins and Protein Derivatives
2.4.3. Protein Digestibility
2.5. In Vitro Antioxidant Activity
2.6. Characterization of the Antioxidant Compounds
2.6.1. Purification of Antioxidant Peptides
2.6.2. Stability to Digestion and Thermal Treatments
2.6.3. Identification of Antioxidant Peptides
2.6.4. Extraction of Phenolic Compounds and Qualitative and Quantitative Analysis by UPLC-PDA-ESI-QTOF
2.7. Antioxidant Activity on Human Keratinocytes Cell Cultures
2.7.1. Cytotoxicity
2.7.2. Protective Effect on Oxidative-Induced Stress
2.8. Statistical Analysis
3. Results
3.1. Microbiological Analysis
3.2. Bioprocessing
3.2.1. Acidification
3.2.2. Proteins and Protein Derivatives
3.2.3. In Vitro Protein Digestibility
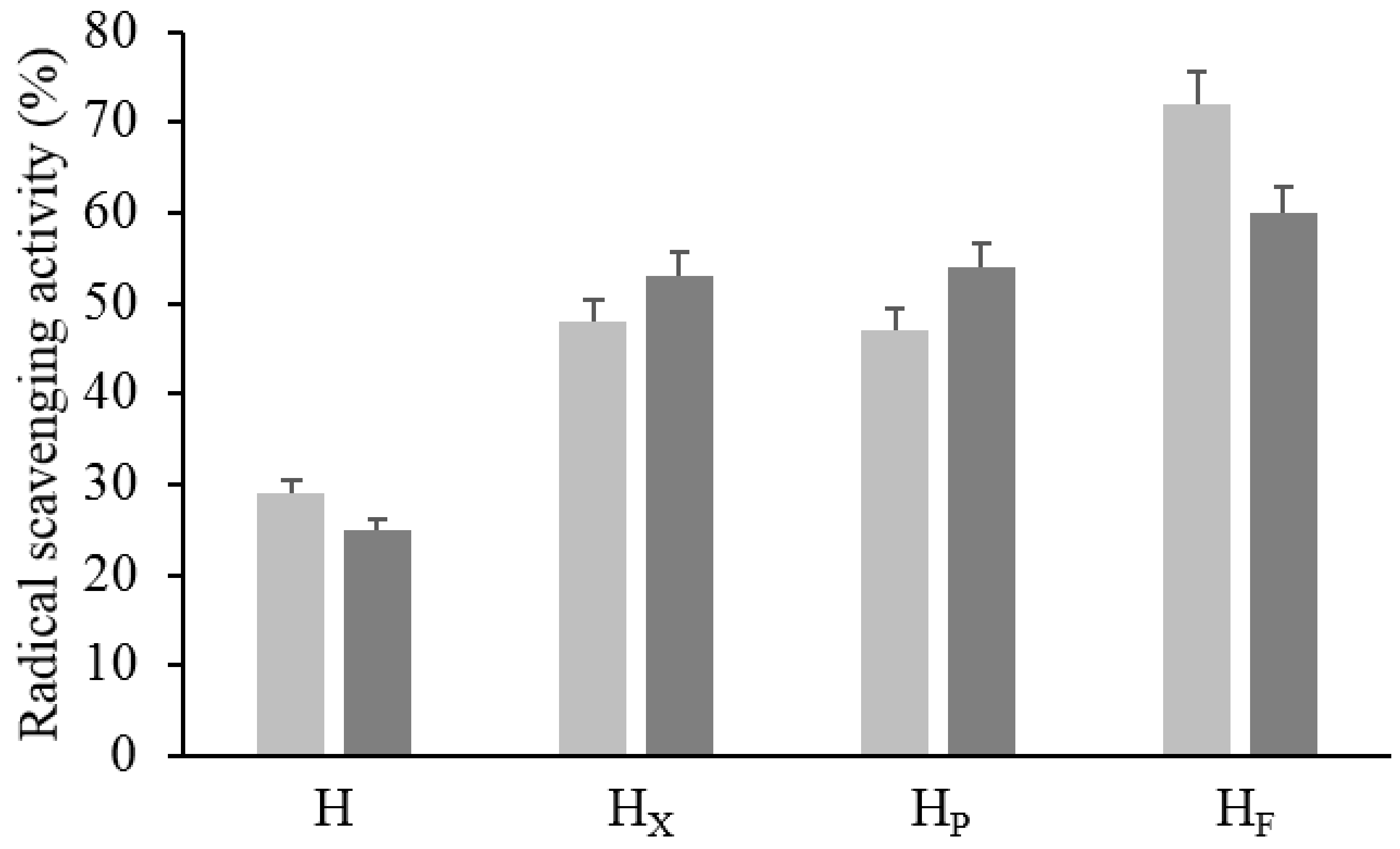
3.2.4. In Vitro Radical Scavenging Activity
3.3. Identification of the Antioxidant Compounds
3.3.1. Peptides Identification
3.3.2. Phenolic Profile
3.3.3. Quantification of Phenolic Compounds
3.4. Citotoxicity and Antioxidant Activity on Human Keratinocytes
3.4.1. Cytotoxicity
3.4.2. Protective Effect towards Oxidative-Induced Stress in Human Keratinocytes Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Farinon, B.; Molinari, R.; Costantini, L.; Merendino, N. The seed of industrial hemp (Cannabis sativa L.): Nutritional quality and potential functionality for human health and nutrition. Nutrients 2020, 12, 1935. [Google Scholar] [CrossRef] [PubMed]
- Leonard, W.; Zhang, P.; Ying, D.; Fang, Z. Hempseed in food industry: Nutritional value, health benefits, and industrial applications. Compr. Rev. Food Sci. Food Saf. 2019, 19, 282–308. [Google Scholar] [CrossRef] [Green Version]
- House, J.D.; Naufeld, J.; Leson, G. Evaluating the quality of protein from hemp seed (Cannabis sativa L.) products through the use of the protein digestibility-corrected amino acid score method. J. Agric. Food Chem. 2010, 58, 11801–11807. [Google Scholar] [CrossRef] [PubMed]
- Siano, F.; Moccia, S.; Picariello, G.; Russo, G.L.; Sorrentino, G.; Di Stasio, M.; La Cara, F.; Volpe, M.G. Comparative study of chemical, biochemical characteristic and atr-ftir analysis of seeds, oil and flour of the edible fedora cultivar hemp (Cannabis sativa L.). Molecules 2018, 24, 83. [Google Scholar] [CrossRef] [Green Version]
- Vonapartis, E.; Aubin, M.; Seguin, P.; Mustafa, A.F.; Charron, J. Seed composition of ten industrial hemp cultivars approved for production in Canada. J. Food Compos. Anal. 2015, 39, 8–12. [Google Scholar] [CrossRef]
- Irakli, M.; Tsaliki, E.; Kalivas, A.; Kleisiaris, F.; Sarrou, E.; Cook, C.M. Effect οf genotype and growing year on the nutritional, phytochemical, and antioxidant properties of industrial hemp (Cannabis sativa L.) Seeds. Antioxidants 2019, 8, 491. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Bazer, F.W.; Davis, T.A.; Kim, S.W.; Li, P.; Rhoads, J.M.; Satterfield, M.C.; Smith, S.B.; Yin, Y. Arginine metabolism and nutrition in growth, health and disease. Amino Acids 2009, 37, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Russo, R.; Reggiani, R. Variability of antinutritional compounds in hempseed meal of italian and french varieties. Plant 2013, 1, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.; Pingitore, E.V.; Mozzi, F.; Saavedra, L.; Villegas, M.J.; Hebert, M.E. Lactic acid bacteria as cell factories for the generation of bioactive peptides. Protein Pept. Lett. 2017, 24, 146–155. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Polo, A.; Rizzello, C.G. The sourdough fermentation is the powerful process to exploit the potential of legumes, pseudo-cereals and milling by-products in baking industry. Crit. Rev. Food Sci. 2019, 60, 2158–2173. [Google Scholar] [CrossRef]
- Gänzle, M.G. Food fermentations for improved digestibility of plant foods–an essential ex situ digestion step in agricultural societies? Curr. Opin. Food Sci. 2020, 32, 124–132. [Google Scholar] [CrossRef]
- Nionelli, L.; Montemurro, M.; Pontonio, E.; Verni, M.; Gobbetti, M.; Rizzello, C.G. Pro-technological and functional characterization of lactic acid bacteria to be used as starters for hemp (Cannabis sativa L.) sourdough fermentation and wheat bread fortification. Int. J. Food Microbiol. 2018, 279, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Shen, P.; Gao, Z.; Xu, M.; Ohm, J.B.; Rao, J.; Chen, B. The impact of hempseed dehulling on chemical composition, structure properties and aromatic profile of hemp protein isolate. Food Hydrocoll. 2020, 106, 105889. [Google Scholar] [CrossRef]
- Arte, E.; Rizzello, G.C.; Verni, M.; Nordlund, E.; Katina, K.; Coda, R. Impact of enzymatic and microbial bioprocessing on protein modification and nutritional properties of wheat bran. J. Agric. Food Chem. 2015, 63, 8685–8693. [Google Scholar] [CrossRef] [PubMed]
- Gobbetti, M.; Gänzle, M. Physiology and biochemistry of lactic acid bacteria. In Handbook on Sourdough Biotechnology; Gobbetti, M., Gänzle, M., Eds.; Springer: New York, NY, USA, 2013; pp. 183–216. [Google Scholar]
- Gobbetti, M.; Rizzello, C.G.; Di Cagno, R.; De Angelis, M. How the sourdough may affect the functional features of leavened baked goods. Food Microbiol. 2014, 37, 30–40. [Google Scholar] [CrossRef]
- Katina, K.; Juvonen, R.; Laitilia, A.; Flander, L.; Nordlund, E.; Kariluoto, S.; Piironen, V.; Poutanen, K. Fermented wheat bran as a functional ingredient in baking. Cereal Chem. 2017, 89, 126–135. [Google Scholar] [CrossRef]
- Filannino, P.; Di Cagno, R.; Gobbetti, M. Metabolic and functional paths of lactic acid bacteria in plant foods: Get out of the labyrinth. Curr. Opin. Biotechnol. 2018, 49, 64–72. [Google Scholar] [CrossRef]
- Verni, M.; Pontonio, E.; Krona, A.; Jacob, S.; Pinto, D.; Rinaldi, F.; Verardo, V.; Diaz de Cerio, E.; Coda, R.; Rizzello, C.G. Bioprocessing of brewers’ spent grain enhances its antioxidant activity: Characterization of phenolic compounds and bioactive peptides. Front. Microbiol. 2020, 11, 1831. [Google Scholar] [CrossRef]
- Pontonio, E.; Di Cagno, R.; Tarraf, W.; Filannino, P.; De Mastro, G.; Gobbetti, M. Dynamic and assembly of epiphyte and endophyte lactic acid bacteria during the life cycle of Origanum vulgare L. Front. Microbiol. 2018, 9, 1372. [Google Scholar] [CrossRef]
- Weiss, W.; Vogelmeier, C.; Gorg, A. Electrophoretic characterization of wheat grain allergens from different cultivars involved in bakers’ asthma. Electrophoresis 1993, 14, 805–816. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Nionelli, L.; Coda, R.; De Angelis, M.; Gobbetti, M. Effect of sourdough fermentation on stabilisation, and chemical and nutritional characteristics of wheat germ. Food Chem. 2010, 119, 1079–1089. [Google Scholar] [CrossRef]
- Breadford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Di Cagno, R.; De Angelis, M.; Lavermicocca, P.; De Vincenzi, M.; Giovannini, C.; Gobbetti, M. Proteolysis by sourdough lactic acid bacteria: Effects on wheat flour protein fractions and gliadin peptides involved in human cereal intolerance. Appl. Environ. Microbiol. 2002, 68, 623–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bini, L.; Magi, B.; Marzocchi, B.; Arcuri, F.; Tripodi, S.; Cintorino, M.; Sanchez, J.C.; Frutiger, S.; Hughes, G.; Pallini, V.; et al. Protein expression profiles in human breast ductal carcinoma and histologically normal tissue. Electrophoresis 1997, 18, 2832–2841. [Google Scholar] [CrossRef] [PubMed]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignazni, G.L. Spectrophotometric assay using o-phthaldialdehyde for determination of proteolysis in milk and isolated milk proteins. J. Dairy Sci. 1983, 66, 1219–1227. [Google Scholar] [CrossRef]
- Akeson, W.R.; Stahmann, M.A. A pepsin pancreatin digest index of protein quality evaluation. J. Nutr. 1964, 83, 257–261. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Curiel, J.; Nionelli, L.; Vincentini, O.; Di Cagno, R.; Silano, M.; Gobbetti, M.; Coda, R. Use of fungal proteases and selected sourdough lactic acid bacteria for making wheat bread with an intermediate content of gluten. Food Microbiol. 2014, 37, 59–68. [Google Scholar] [CrossRef]
- Yu, L.; Perret, J.; Harris, M.; Wilson, J.; Haley, S. Antioxidant properties of bran extracts from “Akron” wheat grown at different locations. J. Agric. Food Chem. 2003, 51, 1566–1570. [Google Scholar] [CrossRef]
- Pasini, G.; Simonato, B.; Giannattasio, M.; Peruffo, A.D.; Curioni, A. Modifications of wheat flour proteins during in vitro digestion of bread dough, crumb, and crust: An electrophoretic and immunological study. J. Agric. Food Chem. 2001, 49, 2254–2261. [Google Scholar] [CrossRef]
- Chen, Y.; Kwon, S.W.; Kim, S.C.; Zhao, Y. Integrated approach for manual evaluation of peptides identified by searching protein sequence databases with tandem mass spectra. J. Proteome Res. 2005, 4, 998–1005. [Google Scholar] [CrossRef]
- Verardo, V.; Arraìez-Romaìn, D.; Segura-Carretero, A.; Marconi, E.; Fernaìndez-Gutieìrrez, A.; Caboni, M.F. Determination of free and bound phenolic compounds in buckwheat spaghetti by RP-HPLC-ESI-TOF-MS: Effect of thermal processing from farm to fork. J. Agric. Food Chem. 2011, 59, 7700–7707. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Caravaca, A.M.; Verardo, V.; Berardinelli, A.; Marconi, E.; Caboni, M.F. A chemometric approach to determine the phenolic compounds in different barley samples by two different stationary phases: A comparison between C18 and pentafluorophenyl core shell columns. J. Chromatogr. A 2014, 1355, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Coda, R.; Rizzello, C.G.; Pinto, D.; Gobbetti, M. Selected lactic acid bacteria synthesize antioxidant peptides during sourdough fermentation of cereal flours. Appl. Environ. Microbiol. 2012, 78, 1087–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aluko, R.E. Hemp Seed (Cannabis sativa L.) Proteins: Composition, structure, enzymatic modification, and functional or bioactive properties. In Sustainable Protein Sources; Nadathur, S.R., Wanasundara, J.P.D., Scanlin, L., Eds.; Elsevier: Amsterdam, The Netherlands; Academic Press Inc.: San Diego, CA, USA, 2017; pp. 121–132. [Google Scholar]
- López-Cobo, A.; Gómez-Caravaca, A.M.; Pasini, F.; Caboni, M.F.; Segura-Carretero, A.; Fernández-Gutiérrez, A. HPLC-DAD-ESI-QTOF-MS and HPLC-FLD-MS as valuable tools for the determination of phenolic and other polar compounds in the edible part and by-products of avocado. LWT 2016, 73, 505–513. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, C.; Gómez-Caravaca, A.M.; Guerra-Hernández, E.; Cerretani, L.; García-Villanova, B.; Verardo, V. Comprehensive metabolite profiling of Solanum tuberosum L.(potato) leaves by HPLC-ESI-QTOF-MS. Food Res. Int. 2018, 112, 390–399. [Google Scholar] [CrossRef]
- Rodríguez, H.; Landete, J.M.; De Las Rivas, B.; Muñoz, R. Metabolism of food phenolic acids by Lactobacillus plantarum CECT 748T. Food Chem. 2008, 107, 1393–1398. [Google Scholar] [CrossRef] [Green Version]
- Frassinetti, S.; Moccia, E.; Caltavuturo, L.; Gabriele, M.; Longo, V.; Bellani, L.; Giorgi, G.; Giorgetti, L. Nutraceutical potential of hemp (Cannabis sativa L.) seeds and sprouts. Food Chem. 2018, 262, 56–66. [Google Scholar] [CrossRef]
- Fabre, N.; Rustan, I.; De Hoffmann, E.; Quetin-Leclercq, J. Determination of flavone, flavonol, and flavanone aglycones by negative ion liquid chromatography electrospray ion trap mass spectrometry. J. Am. Soc. Mass Spectrom. 2001, 12, 707–715. [Google Scholar] [CrossRef] [Green Version]
- Davoli, P.; Weber, R.W. Simple method for reversed-phase high-performance liquid chromatographic analysis of fungal pigments in fruit-bodies of Boletales (Fungi). J. Chromatogr. A 2002, 964, 129–135. [Google Scholar] [CrossRef]
- Keskin, Ş.; Şirin, Y.; Çakir, H.E.; Keskin, M. An investigation of Humulus lupulus L.: Phenolic composition, antioxidant capacity and inhibition properties of clinically important enzymes. S. Afr. J. 2019, 120, 170–174. [Google Scholar] [CrossRef]
- Yan, X.; Tang, J.; Dos Santos Passos, C.; Nurisso, A.; Simoes-Pires, C.A.; Ji, M.; Lou, H.; Fan, P. Characterization of lignanamides from hemp (Cannabis sativa L.) seed and their antioxidant and acetylcholinesterase inhibitory activities. J. Agric. Food Chem. 2015, 63, 10611–10619. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.R.; Qian, P.; Sun, Z.; Zhou, X.H.; Chen, T.P.; He, J.F.; Zhang, H.; Wu, J. Hemp seed protein derived antioxidative peptides: Purification, identification and protection from hydrogen peroxide-induced apoptosis in PC12 cells. Food Chem. 2010, 123, 1210–1218. [Google Scholar] [CrossRef]
- Girgih, A.T.; He, R.; Malomo, S.; Offengenden, M.; Wu, J.; Aluko, R.E. Structural and functional characterization of hemp seed (Cannabis sativa L.) protein-derived antioxidant and antihypertensive peptides. J. Funct. Foods. 2014, 6, 384–394. [Google Scholar] [CrossRef]
- Orio, L.P.; Boschin, G.; Recca, T.; Morelli, C.F.; Ragona, L.; Francescato, P.; Arnoldi, A.; Speranza, G. New ACE-inhibitory peptides from hemp seed (Cannabis sativa L.) proteins. J. Agric. Food Chem. 2017, 65, 10482–10488. [Google Scholar] [CrossRef]
- Wang, Q.; Xiong, Y.L. Processing, nutrition, and functionality of hempseed protein: A review. Compr. Rev. Food. Sci. 2019, 18, 936–952. [Google Scholar] [CrossRef] [Green Version]
- Mattila, P.; Mäkinen, S.; Eurola, M.; Jalava, T.; Pihlava, J.-M.; Hellström, J.; Pihlanto, A. Nutritional value of commercial protein-rich plant products. Plant Foods Hum. Nutr. 2018, 73, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Pal, R.S.; Bhartiya, A.; Arunkumar, R.; Kant, L.; Aditya, J.P.; Bisht, J.K. Impact of dehulling and germination on nutrients, antinutrients, and antioxidant properties in horsegram. J. Food Sci. Technol. 2016, 53, 337–347. [Google Scholar] [CrossRef] [Green Version]
- Pallavi, J.; Varma, K. Effect of germination and dehulling on the nutritive value of soybean. Nutr. Food Sci. 2016, 46, 595–603. [Google Scholar]
- Rommi, K.; Hakala, T.K.; Holopainen, U.; Nordlund, E.; Poutanen, K.; Lantto, R. Effect of enzyme-aided cell wall disintegration on protein extractability from intact and dehulled rapeseed (Brassica rapa L. and Brassica napus L.) press cakes. J. Agric. Food Chem. 2014, 62, 7989–7997. [Google Scholar] [CrossRef]
- Kabera, J.N.; Semana, E.; Mussa, A.R.; He, X. Plant Secondary Metabolites: Biosynthesis, Classification, Function and Pharmacological Properties. J. Pharm. Pharmacol. 2014, 2, 377–392. [Google Scholar]
- Tang, C.; Wang, X.; Yang, X. Enzymatic hydrolysis of hemp (Cannabis sativa L.) protein isolate by various proteases and antioxidant properties of the resulting hydrolysates. Food Chem. 2009, 114, 1484–1490. [Google Scholar] [CrossRef]
- The, S.S.; Bekhit, A.E.D.A.; Carne, A.; Birch, J. Antioxidant and ACE-inhibitory activities of hemp (Cannabis sativa L.) protein hydrolysates produced by the proteases AFP, HT, Pro-G, actinidin and zingibain. Food Chem. 2016, 203, 199–206. [Google Scholar]
- Girgih, A.T.; Alashi, A.M.; He, R.; Malomo, S.A.; Raj, P.; Netticadan, T.; Aluko, R.E. A novel hemp seed meal protein hydrolysate reduces oxidative stress factors in spontaneously hypertensive rats. Nutrients 2014, 6, 5652–5666. [Google Scholar] [CrossRef] [Green Version]
- Schettino, R.; Pontonio, E.; Rizzello, C.G. Use of fermented hemp, chickpea and milling by-products to improve the nutritional value of semolina pasta. Foods 2019, 8, 604. [Google Scholar] [CrossRef] [Green Version]
- Hess, J.M. Role of lactic acid bacteria in impacting nutrient bioavailability. In Lactic Acid Bacteria: A Functional Approach; De Albuquerque, M.A.C., De LeBlanc, A.M., LeBlanc, J.G., Bedani, R., Eds.; CRC Press: Boca Raton, FL, USA, 2020; Volume 3, pp. 35–57. [Google Scholar]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Calasso, M.; Archetti, G.; Rizzello, C.G. Novel insights on the functional/nutritional features of the sourdough fermentation. Int. J. Food Microbiol. 2019, 302, 103–113. [Google Scholar] [CrossRef]
- Bartkiene, E.; Schleining, G.; Krungleviciute, V.; Zadeike, D.; Zavistanaviciute, P.; Dimaite, I.; Kuzmaite, I.; Riskeviciene, V.; Juodeikiene, G. Development and quality evaluation of lacto-fermented product based on hulled and not hulled hempseed (Cannabis sativa L.). LWT-Food Sci. Technol. 2016, 72, 544–551. [Google Scholar] [CrossRef]
- Logarušić, M.; Slivac, I.; Radošević, K.; Bagović, M.; Redovniković, I.R.; Srček, V.G. Hempseed protein hydrolysates’ effects on the proliferation and induced oxidative stress in normal and cancer cell lines. Mol. Biol. Rep. 2019, 46, 6079–6085. [Google Scholar] [CrossRef]
- Tang, C.; Wang, X.; Yang, X. Physicochemical and functional properties of hemp (Cannabis sativa L.) protein. J. Agric. Food Chem. 2006, 54, 8945–8950. [Google Scholar] [CrossRef]
- Patel, S.; Cudney, R.; McPherson, A. Crystallographic characterization and molecular symmetry of edestin, a legumin from hemp. J. Mol. Biol. 1994, 235, 361–363. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.S.; Tang, C.H.; Yang, X.Q.; Guo, W.R. Characterization, amino acid composition and in vitro digestibility of hemp (Cannabis sativa L.) proteins. Food Chem. 2008, 107, 11–18. [Google Scholar] [CrossRef]
- Docimo, T.; Caruso, I.; Ponzoni, E.; Mattana, M.; Galasso, I. Molecular characterization of edestin gene family in Cannabis sativa L. Plant Physiol. Biochem. 2014, 84, 142–148. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Verni, M.; Bordignon, S.; Gramaglia, V.; Gobbetti, M. Hydrolysate from a mixture of legume flours with antifungal activity as an ingredient for prolonging the shelf-life of wheat bread. Food Microbiol. 2017, 64, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Montemurro, M.; Pontonio, E.; Gobbetti, M.; Rizzello, G.C. Investigation of the nutritional, functional and technological effects of the sourdough fermentation of sprouted flours. Int. J. Food Microbiol. 2019, 302, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Gänzle, M.; Loponen, J.; Gobbetti, M. Proteolysis in sourdough fermentations: Mechanisms and potential for improved bread quality. Trends Food Sci. Technol. 2008, 19, 513–521. [Google Scholar] [CrossRef]
- Görgüç, A.; Gençdağ, E.; Yılmaz, F.M. Bioactive peptides derived from plant origin by-products: Biological activities and techno-functional utilizations in food developments—A review. Food Res. Int. 2020, 136, 109504. [Google Scholar] [CrossRef]
- Chen, H.M.; Muramoto, K.; Yamauchi, F.; Fujimoto, K.; Nokihara, K. Antioxidative properties of histidine-containing peptides designed from peptide fragments found in the digests of a soybean protein. J. Agric. Food Chem. 1998, 46, 49–53. [Google Scholar] [CrossRef]
- Chen, H.M.; Muramoto, K.; Yamauchi, F.; Nokihara, K. Antioxidant activity of design peptides based on the antioxidative peptide isolated from digests of a soybean protein. J. Agric. Food Chem. 1996, 44, 2619–2623. [Google Scholar] [CrossRef]
- Li, X.X.; Han, L.J.; Chen, L.J. In vitro antioxidant activity of protein hydrolysates prepared from corn gluten meal. J. Sci. Food Agric. 2008, 88, 1660–1666. [Google Scholar] [CrossRef]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef]
- Chen, T.; He, J.; Zhang, J.; Li, X.; Zhang, H.; Li, L. The isolation and identification of two compounds with predominant radical scavenging activity in hempseed (seed of Cannabis sativa L.). Food Chem. 2012, 134, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- Filannino, P.; Gobbetti, M.; De Angelis, M.; Di Cagno, R. Hydroxycinnamic acids used as external acceptors of electrons: An energetic advantage for strictly heterofermentative lactic acid bacteria. Appl. Environ. Microbiol. 2014, 80, 7574–7582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najmanová, I.; Pourová, J.; Vopršalová, M.; Pilařová, V.; Semecký, V.; Nováková, L.; Mladěnka, P. Flavonoid metabolite 3-(3-hydroxyphenyl) propionic acid formed by human microflora decreases arterial blood pressure in rats. Mol. Nutr. Food Res. 2016, 60, 981–991. [Google Scholar] [CrossRef] [PubMed]
- Hadnađev, M.; Dizdar, M.; Hadnađev-Dapčević, T.; Jovanov, P.; Mišan, A.; Sakač, M. Hydrolyzed hemp seed proteins as bioactive peptides. J. Process. Energy Agric. 2018, 22, 90–94. [Google Scholar] [CrossRef] [Green Version]
- Verni, M.; Verardo, V.; Rizzello, C.G. How fermentation affects the antioxidant properties of cereals and legumes. Foods 2019, 8, 362. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H | HX | HP | HF | |
|---|---|---|---|---|
| Microbiological characteristics | ||||
| TMB (log10 cfu/g) | 3.5 ± 0.2 c | 5.8 ± 0.1 b | 5.8 ± 0.1 b | 9.0 ± 0.2 a |
| LAB (log10 cfu/g) | 3.5 ± 0.3 c | 4.7 ± 0.2 b | 4.7 ± 0.1 b | 9.0 ± 0.2 a |
| Yeasts (log10 cfu/g) | 3.1 ± 0.2 b | 4.1 ± 0.3 a | 4.0 ± 0.2 a | 3.9 ±0.1 a |
| Molds (log10 cfu/g) | 2.5 ± 0.2 b | 3.4 ± 0.3 a | 3.2 ± 0.2 a | 3.1 ±0.1 a |
| Enterobacteriacae (log10 cfu/g) | 4.7 ± 0.1 b | 5.9 ± 0.2 a | 5.7 ± 0.2 a | 4.6 ± 0.1 b |
| Biochemical properties | ||||
| pH | 6.60 ± 0.03 a | 5.61 ± 0.02 c | 5.77 ± 0.03 b | 5.21 ± 0.04 d |
| Lactic acid (mmol/kg) | 1.79 ± 0.24 d | 50.18 ± 1.2 b | 40.93 ± 1.1 c | 88.25 ± 2.2 a |
| Acetic acid (mmol/kg) | 3.16 ± 0.31 d | 18.46 ± 0.94 b | 13.40 ± 1.11 c | 37.67 ± 2.4 a |
| TFAA (mg/kg) | 1656 ± 23 d | 4843 ± 27 c | 5115 ± 31 b | 5198 ± 26 a |
| Peptides (mg/kg) | 259.84 ± 10 d | 338.13 ± 11 c | 371.31 ± 23 a | 350.92 ± 15 b |
| Fraction | Sequence | Weight (Da) | Length (aa) | Net Charge | Hydrophobic-AA (%) | BCAA * (%) | NCBI Accession n° (Protein) | Fragment |
|---|---|---|---|---|---|---|---|---|
| 1 | ALASIGKATR | 986.6 | 10 | 2 | 60 | 20 | XP_030502118.1 (nephrocystin-3) | 233–242 |
| 2 | IGQSHPQALMYPLLVACKSISNLR | 2638.4 | 24 | 3 | 52 | 28 | XP_030508478.1 (Serine/threonine-protein kinase TOR) | 1863–1886 |
| 5 | AQVSVGGGR | 829.4 | 9 | 2 | 66 | 22 | XP_030499964.1 (edestin 3) | 119–127 |
| 7 | AIENGAVSVSEPEEK | 1557.7 | 15 | 2 | 44 | 19 | XP_030503904.1 (uncharacterized protein) | 99–113 |
| DLQIIAPSR | 1011.5 | 9 | 2 | 55 | 33 | SNQ45155.1 (edestin 3) | 286–194 |
| N° | Polar Compound | RT (min) | Molecular Formula | m/z Expected | m/z Calculated | Error (ppm) | Score (%) | Fragments (MS2) |
|---|---|---|---|---|---|---|---|---|
| Free compounds | ||||||||
| 1 | Protocatechuic acid-glucoside isomer 1 | 2.946 | C13H16O9 | 315.0703 | 315.0716 | −4.1 | 99.78 | |
| 2 | Protocatechuic acid-glucoside isomer 2 | 3.281 | C13H16O9 | 315.0713 | 315.0716 | −1 | 98.96 | |
| 3 | Catechol | 4.560 | C6H6O2 | 109.0288 | 109.0290 | −1.8 | 92.24 | |
| 4 | 4-vinyl catechol | 4.730 | C8H8O2 | 135.0435 | 135.0446 | −8.1 | 87.98 | |
| 5 | Tuberonic acid glucoside isomer 1 | 7.939 | C18H28O9 | 387.1649 | 387.1655 | −1.5 | 94.94 | |
| 6 | 4-vinyl guaiacol | 8.178 | C9H10O2 | 149.0603 | 149.1770 | −14.1 | 79.88 | |
| 7 | Tuberonic acid glucoside isomer 2 | 8.295 | C18H28O9 | 387.1642 | 387.1655 | −3.4 | 94.94 | |
| 8 | 3-(3-Hydroxyphenyl)propionic acid | 8.530 | C9H10O3 | 165.0543 | 165.0552 | −5.5 | 92.86 | |
| 9 | 4-vinyl phenol | 8.543 | C8H8O0 | 119.0489 | 119.0497 | −6.7 | 81.46 | |
| 10 | Tuberonic acid glucoside isomer 3 | 8.783 | C18H28O9 | 387.1646 | 387.1655 | −2.3 | 94.94 | |
| 11 | Tuberonic acid glucoside isomer 4 | 8.882 | C18H28O9 | 387.1656 | 387.1655 | 0.3 | 94.94 | |
| 12 | Tuberonic acid glucoside isomer 5 | 9.797 | C18H28O9 | 387.1652 | 387.1655 | −0.8 | 94.94 | |
| 13 | N-caffeoyltyramine | 12.3 | C17H17NO4 | 298.1074 | 298.1079 | −1.7 | 99.54 | |
| 14 | Luteolin | 12.63 | C15H9O6 | 285.0396 | 285.0396 | −1.1 | 98.68 | |
| 15 | N-caffeoyltyramine | 13.81 | C17H17NO4 | 298.1074 | 298.1079 | −1.7 | 99.54 | |
| 16 | Cannabisin B isomer 1 | 15.98 | C34H32N2O8 | 595.2066 | 595.2080 | −2.4 | 78.54 | 432.1450, 322.1068, 269.0814 |
| 17 | Cannabisin B isomer 2 | 16.18 | C34H32N2O8 | 595.2076 | 595.2080 | −0.7 | 78.54 | 432.1450, 322.1068, 269.0814 |
| 18 | Cannabisin B isomer 3 | 16.81 | C34H32N2O8 | 595.2079 | 595.2080 | −0.1 | 78.54 | |
| 19 | Cannabisin B isomer 4 | 17.3 | C34H32N2O8 | 595.209 | 595.2080 | 1.7 | 78.54 | 432.1450, 322.1068, 269.0814 |
| 20 | Cannabisin C isomer 1 | 17.61 | C35H34N2O8 | 609.2242 | 609.2237 | 0.8 | 76.51 | |
| 21 | Cannabisin C isomer 2 | 17.87 | C35H34N2O8 | 609.2249 | 609.2237 | 2 | 76.51 | 446.1611, 283.0970, 268.0736 |
| 22 | Cannabisin C isomer 3 | 18.38 | C35H34N2O8 | 609.2243 | 609.2237 | 1 | 76.51 | |
| 23 | Cannabisin D isomer 1 | 19.04 | C36H36N2O8 | 623.2393 | 623.2393 | 0 | 70.77 | 460.1760, 445.1523 |
| 24 | Cannabisin D isomer 2 | 19.35 | C36H36N2O8 | 623.2386 | 623.2393 | −1.1 | 70.77 | 460.1760, 445.1523 |
| 25 | Cannabisin D isomer 3 | 20.73 | C36H36N2O8 | 623.2386 | 623.2393 | −1.1 | 70.77 | |
| 26 | Trihydroxyoctadeceno ic acid | 21.11 | C18H34O5 | 329.2320 | 329.2328 | −2.4 | 98.65 | |
| 27 | Trihydroxyoctadeceno ic acid | 21.21 | C18H34O5 | 329.2319 | 329.2328 | −0.9 | 98.65 | |
| 28 | Trihydroxyoctadeceno ic acid | 21.32 | C18H34O5 | 329.2326 | 329.2328 | −0.2 | 98.65 | |
| 29 | Cannabisin D isomer 4 | 21.57 | C36H36N2O8 | 623.2393 | 623.2393 | 0 | 70.77 | 460.1760, 445.1523 |
| 30 | Cannabisin D isomer 5 | 21.74 | C36H36N2O8 | 623.2409 | 623.2393 | 2.6 | 70.77 | |
| 31 | Cannabidiolic acid glucoside | 23.35 | C28H40O9 | 519.2593 | 519.2594 | −0.2 | 56.29 | 373.1255 |
| 32 | Cannabielsoic acid | 23.61 | C22H30O5 | 373.2012 | 373.2015 | −0.8 | 97.08 | |
| 33 | Cannabielsoic acid | 23.65 | C22H30O5 | 373.2014 | 373.2015 | −0.3 | 97.08 | |
| 34 | Cannabielsoic acid | 23.86 | C22H30O5 | 373.2008 | 373.2015 | −0.7 | 97.08 | |
| 35 | Cannabielsoic acid | 23.97 | C22H30O5 | 373.2015 | 373.2015 | 0 | 97.08 | |
| 36 | Cannabielsoic acid | 24.12 | C22H30O5 | 373.2008 | 373.2015 | −0.7 | 97.08 | |
| 37 | Cannabielsoic acid | 24.41 | C22H30O5 | 373.2013 | 373.2015 | −0.5 | 97.08 | |
| 38 | Cannabielsoic acid | 24.92 | C22H30O5 | 373.2025 | 373.2015 | 2.7 | 97.08 | |
| 39 | THCA derivative | 25.18 | C46H58O8 | 737.4056 | 737.4053 | 0.4 | 96.53 | |
| 40 | Fatty acid C18:3 | 25.77 | C18H30O2 | 277.2158 | 277.2168 | −3.6 | 99.75 | |
| 41 | Fatty acid C18:3 | 25.806 | C18H30O2 | 277.2164 | 277.2168 | −1.4 | 97.64 | |
| 42 | Fatty acid C18:2 | 26.117 | C18H32O2 | 279.232 | 279.2324 | −1.14 | 91.75 | |
| Bound compounds | ||||||||
| 1 | Citric acid | 0659 | C6H8O7 | 191.0182 | 191.0192 | −5.2 | 79.55 | 111.0079 |
| 2 | Citric acid | 0.882 | C6H8O8 | 191.0185 | 191.0192 | −3.7 | 79.55 | 111.0079 |
| 3 | Protocatechuic acid derivative | 3.231 | C15H8O9 | 331.0024 | 331.256 | 1.2 | 98.65 | 153.0186, 109.0287 |
| 4 | Protocatechuic acid glucoside | 4.357 | C13H16O9 | 315.0654 | 315.0716 | −19.7 | 70.77 | |
| 5 | Salicylic acid derivative | 4.849 | C15H8O7 | 299.0131 | 299.0192 | −17.7 | 98.65 | 137.0233 |
| 6 | Benzoic acid aldehyde | 6.566 | C7H6O2 | 121.0283 | 121.029 | −5.8 | 79.27 | 92.0246 |
| 7 | Catechin | 6.619 | C15H14O6 | 289.0702 | 289.0712 | −3.5 | 75.65 | |
| 8 | Xerocomic acid | 8.659 | C18H12O8 | 355.0442 | 355.0454 | −3.1 | 86.29 | 337.0336, 311.0544, 267.0651 |
| 9 | trans p-Coumaric acid | 9.24 | C9H8O3 | 163.0388 | 163.0395 | −4.3 | 98.97 | 119.0485 |
| 10 | cis p-Coumaric acid | 9.47 | C9H8O3 | 163.0386 | 163.0395 | −5.5 | 96.28 | 119.0485 |
| 11 | Coumaric acid derivative | 9.84 | C18H16O6 | 327.0858 | 327.0869 | −4.3 | 95.71 | 163.039, 119.0481 |
| 12 | Coumaric acid derivative | 10.36 | C18H16O6 | 327.0858 | 327.0869 | −4.3 | 95.71 | 163.039, 119.0481 |
| 13 | Flavodic acid | 10.87 | C19H14O8 | 369.0589 | 369.061 | −5.7 | 96.08 | 337.0344, 265.0555 |
| 14 | Flavodic acid | 12.151 | C19H14O8 | 369.0589 | 369.061 | −2.4 | 95.98 | 337.0344. 265.0493 |
| 15 | Flavodic acid | 12.8 | C19H14O8 | 369.0589 | 369.061 | −2.4 | 95.98 | |
| 16 | N-caffeoyltyramine | 13.87 | C17H17NO4 | 298.1096 | 298.1079 | −5 | 98.29 | 178.0506, 135.0434 |
| 17 | Flavodic acid derivative | 14.11 | C20H16O8 | 383.0779 | 383.0767 | 3.1 | 95.56 | 337.0344, 351.0500, 263.0555 |
| 18 | Cannabisin B isomer 1 | 16.06 | C34H32N2O8 | 595.2089 | 595.208 | −1.3 | 97.08 | |
| 19 | Cannabisin B isomer 2 | 16.23 | C34H32N2O8 | 595.2089 | 595.208 | 1.5 | 97.08 | 432.1450, 322.1068, 269.0814 |
| 20 | N-trans-feryroyltyramine | 16.544 | C18H19NO4 | 312.1225 | 312.1236 | −3.5 | 92.64 | 190.0481, 135.0426, 178.0484 |
| 21 | Cannabisin B isomer 3 | 16.853 | C34H32N2O8 | 595.2092 | 595.208 | 2 | 97.08 | 432.1450, 322.1068, 269.0814 |
| 22 | Cannabisin B isomer 4 | 17.24 | C34H32N2O8 | 595.2092 | 595.208 | 1 | 97.08 | |
| 23 | Cannabisin B isomer 5 | 17.366 | C34H32N2O8 | 595.2072 | 595.208 | −1.3 | 97.08 | 485.1701, 432.1450, 322.1068, 269.0814 |
| 24 | Cannabisin C isomer 1 | 17.65 | C35H34N2O8 | 609.2229 | 609.2237 | −1.3 | 96.22 | |
| 25 | Cannabisin B isomer 6 | 17.773 | C34H32N2O8 | 595.2065 | 595.208 | −2.5 | 97.08 | 322.1068, 269.0814 |
| 26 | Cannabisin C isomer 2 | 17.93 | C35H34N2O8 | 609.2229 | 609.2237 | −1.1 | 96.22 | |
| 27 | Cannabisin C isomer 3 | 18.12 | C35H34N2O8 | 609.2229 | 609.2237 | −1.2 | 96.22 | |
| 28 | Cannabisin C isomer 4 | 18.42 | C35H34N2O8 | 609.2231 | 609.2237 | −1 | 96.22 | 446.1611, 283.0970, 268.0736 |
| 29 | Cannabisin D isomer 1 | 19.06 | C36H36N2O8 | 623.2392 | 623.2393 | −0.2 | 98.39 | 460.1760, 445.1523 |
| 30 | Cannabisin B isomer 7 | 19.19 | C34H32N2O8 | 595.2065 | 595.208 | −2.5 | 97.08 | 432.1450, 322.1068, 269.0814 |
| 31 | Cannabisin B isomer 8 | 19.55 | C34H32N2O8 | 595.2065 | 595.208 | −2.5 | 97.08 | 432.1450, 322.1068, 269.0814 |
| 32 | Cannabisin D isomer 2 | 19.378 | C36H36N2O8 | 623.2402 | 623.2393 | 1.4 | 98.39 | |
| 33 | Cannabisin E isomer 1 | 19.633 | C36H38N2O9 | 641.2493 | 641.2499 | 1.6 | 97.81 | 489.2026, 328.1187, 151.0380 |
| 34 | Cannabisin E isomer 2 | 19.92 | C36H38N2O9 | 641.2509 | 641.2499 | 1.6 | 97.81 | 489.2026, 328.1187, 151.0380 |
| 35 | Cannabisin C isomer 5 | 20.156 | C35H34N2O8 | 609.2242 | 609.2237 | 0.8 | 96.22 | |
| 36 | Cannabisin D isomer 3 | 20.21 | C36H36N2O8 | 623.2398 | 623.2393 | 0.8 | 98.39 | |
| 37 | Cannabisin C isomer 6 | 20.52 | C35H34N2O8 | 609.223 | 609.2237 | −1.1 | 96.22 | |
| 38 | Cannabisin D isomer 4 | 21.579 | C36H36N2O8 | 623.2405 | 623.2393 | 1.9 | 98.39 | |
| 39 | THCA derivative | 25.18 | C46H58O8 | 737.4056 | 737.4053 | 0.4 | 96.53 | |
| H | HX | HP | HF | |
|---|---|---|---|---|
| Free phenolics | ||||
| Protocatechuic acid-glucoside | 27.0 ± 0.1 a | 3.7 ± 0.2 c | 6.1 ± 0.1 b | n.d |
| Protocatechuic acid-glucoside | 33.5 ± 0.8 a | 4.99 ± 0.06 c | 8.66 ± 0.07 b | 5.9 ± 0.6 c |
| Catechol | n.d. | 2.94 ± 0.08 a | 2.95 ± 0.01 a | 2.60 ± 0.09 b |
| 4-vinyl catechol | n.d. | n.d. | n.d. | 4.01 ± 0.01 |
| Tuberonic acid glucoside | 3.43 ± 0.05 a | 2.26 ± 0.03 b | 2.30 ± 0.11 b | 1.92 ± 0.01 c |
| 4-vinyl guaiacol | 2.14 ± 0.06 a | n.d. | 1.98 ± 0.01 b | 1.88 ± 0.01 b |
| Tuberonic acid glucoside | 3.09 ± 0.06 a | 1.99 ± 0.02 b | 2.08 ± 0.06 b | 1.89 ± 0.01 b |
| 3-(3-Hydroxyphenyl) propionic acid | 1.96 ± 0.02 c | 3.95 ± 0.07 b | 3.18 ± 0.08 b | 10.48 ± 0.04 a |
| 4-vinyl phenol | 1.87 ± 0.02 c | 2.53 ± 0.00 b | 2.28 ± 0.06 b | 4.70 ± 0.06 a |
| Tuberonic acid glucoside | 3.37 ± 0.01 a | 1.94 ± 0.01 b | 2.13 ± 0.06 b | 1.88 ± 0.01 c |
| Tuberonic acid glucoside | 19.4 ± 0.2 a | 3.32 ± 0.01 c | 5.6 ± 0.3 b | 2.67 ± 0.02 d |
| Tuberonic acid glucoside | 2.23 ± 0.03 a | 2.04 ± 0.04 b | 2.08 ± 0.06 b | 1.81 ± 0.01 c |
| N-caffeoyltyramine | 32.2 ± 0.5 a | 24 ± 2 b | 33 ± 2 a | 31.2 ± 0.6 a |
| Luteolin | <LOQ | <LOQ | <LOQ | <LOQ |
| N-caffeoyltyramine | 29 ± 2 b | 36 ± 1 a | 34 ± 4 a | 26.2 ± 0.9 b |
| Cannabisin B | 0.81 ± 0.05 b | 1.02 ± 0.06 a | 1.0 ± 0.1 a | 0.86 ± 0.05 b |
| Cannabisin B | 20.4 ± 0.4 a | 20 ± 2 a | 21.0 ± 0.1 a | 19.6 ± 0.5 a |
| Cannabisin B | 1.16 ± 0.02 b | 1.51 ± 0.09 a | 1.4 ± 0.2 a | 0.98 ± 0.03 c |
| Cannabisin B | 0.92 ± 0.09 a | 1.11 ± 0.02 a | 1.1 ± 0.2 a | 0.61 ± 0.08 b |
| Cannabisin C | 6.3 ± 0.1 ab | 6.72 ± 0.06 a | 6.5 ± 0.2 a | 5.7 ± 0.2 b |
| Cannabisin C | 5.60 ± 0.02 a | 5.7 ± 0.4 a | 5.9 ± 0.3 a | 5.0 ± 0.4 a |
| Cannabisin C | 3.0 ± 0.2 ab | 3.3 ± 0.3 a | 3.7 ± 0.3 a | 2.61 ± 0.03 b |
| Cannabisin D | 4.4 ± 0.1 a | 4.3 ± 0.5 a | 4.32 ± 0.05 a | 4.0 ± 0.3 a |
| Cannabisin D | 0.93 ± 0.07 a | 1.3 ± 0.3 a | 1.4 ± 0.6 a | 0.38 ± 0.06 b |
| Cannabisin D | 4.80 ± 0.09 b | 4.2 ± 0.4 b | 6.2 ± 0.3 a | 5.9 ± 0.2 a |
| Cannabisin D | 3.1 ± 0.1 b | 5.1 ± 0.2 a | 4.5 ± 0.5 a | 2.1 ± 0.2 c |
| Cannabisin D | 3.3 ± 0.2 a | 4.1 ± 0.6 a | 3.7 ± 0.5 a | 2.37 ± 0.04 b |
| Cannabidiolic acid glucoside | 7.62 ± 0.01 a | 3.3 ± 0.2 b | 4.0 ± 0.2 b | 3.77 ± 0.05 b |
| Cannabielsoic acid | 13.3 ± 0.2 a | 2.9 ± 0.2 d | 4.35 ± 0.01 b | 3.90 ± 0.09 c |
| Cannabielsoic acid | 9.86 ± 0.01 a | 2.7 ± 0.1 d | 3.5 ± 0.1 b | 3.24 ± 0.01 c |
| Cannabielsoic acid | 4.63 ± 0.04 a | 1.90 ± 0.01 c | 2.06 ± 0.01 b | 2.05 ± 0.04 b |
| Cannabielsoic acid | 6.81 ± 0.02 a | 1.96 ± 0.03 d | 2.40 ± 0.01 b | 2.27 ± 0.01 c |
| Cannabielsoic acid | 4.57 ± 0.02 a | 1.96 ± 0.03 d | 2.64 ± 0.09 b | 2.22 ± 0.01 c |
| Cannabielsoic acid | 16.2 ± 0.1 a | 3.0 ± 0.1 b | 3.8 ± 0.1 b | 3.42 ± 0.07 b |
| Cannabielsoic acid | 30 ± 2 a | 3.5 ± 0.4 c | 4.8 ± 0.4 b | 4.6 ± 0.2 b |
| Total | 307 ± 6 a | 170 ± 6 c | 194 ± 4 b | 173 ± 2 c |
| Bound phenolics | ||||
| Salicylic acid derivative | 14.4 ± 0.5 c | 59 ± 2 a | 32 ± 2 b | 7.2 ± 0.5 d |
| Benzoic acid aldehyde | 26 ± 3 a | 7.20 ± 0.20 c | 6.60 ± 0.2 c | 13 ± 1 b |
| Xerocomic acid | 1.37 ± 0.01 c | 3.60 ± 0.20 b | 3.60 ± 0.1 b | 4.10 ± 0.11 a |
| trans p-Coumaric acid | 0.98 ± 0.03 a | 0.91 ± 0.02 a | 0.98 ± 0.02 a | 1.02 ± 0.06 a |
| cis p-Coumaric acid | 0.56 ± 0.02 a | 0.49 ± 0.01 b | 0.54 ± 0.01 a | 0.56 ± 0.01 a |
| Coumaric acid derivative | 0.84 ± 0.05 a | 0.50 ± 0.01 c | 0.68 ± 0.01 b | 0.90 ± 0.01 a |
| Coumaric acid derivative | 1.89 ± 0.07 a | 0.95 ± 0.07 c | 0.85 ± 0.02 c | 1.47 ± 0.02 b |
| Flavodic acid | 0.37 ± 0.02 a | 0.30 ± 0.02 b | 0.32 ± 0.01 b | 0.43 ± 0.01 a |
| Flavodic acid | 0.80 ± 0.10 c | 2.45 ± 0.06 a | 2.03 ± 0.08 b | 2.1 ± 0.10 b |
| Flavodic acid | 0.48 ± 0.01 c | 1.52 ± 0.10 a | 1.14 ± 0.04 b | 1.21 ± 0.04 b |
| N-caffeoyltyramine | 11.04 ± 0.02 c | 24 ± 2 a | 21.0 ± 0.6 b | 23.3 ± 0.9 a |
| Flavodic acid derivative | 0.70 ± 0.03 ab | 0.81 ± 0.07 a | 0.61 ± 0.04 b | 0.75 ± 0.04 a |
| Flavodic acid derivative | 0.23 ± 0.02 a | 0.26 ± 0.03 a | 0.16 ± 0.01 b | 0.24 ± 0.01 a |
| Cannabisin B isomer 1 | 0.17 ± 0.01 c | 0.58 ± 0.02 a | 0.53 ± 0.01 a | 0.26 ± 0.02 b |
| Cannabisin B isomer 2 | 2.01 ± 0.21 b | 4.4 ± 0.50 a | 3.6 ± 0.3 a | 2.4 ± 0.2 b |
| N-trans-feryroyltyramine | 3.51 ± 0.40 a | 3.21 ± 0.00 a | 3.11 ± 0.04 a | 3.5 ± 0.1 a |
| Cannabisin B isomer 3 | 1.12 ± 0.10 c | 1.56 ± 0.01 a | 1.37 ± 0.07 b | 1.55 ± 0.05 a |
| Cannabisin B isomer 4 | 0.14 ± 0.01 d | 0.74 ± 0.09 a | 0.49 ± 0.03 b | 0.32 ± 0.02 c |
| Cannabisin B isomer 5 | 0.61 ± 0.01 c | 1.33 ± 0.03 a | 1.12 ± 0.08 b | 0.9 ± 0.1 b |
| Cannabisin C isomer 1 | 1.17 ± 0.07 b | 2.41 ± 0.10 a | 2.22 ± 0.20 a | 2.30 ± 0.05 a |
| Cannabisin B isomer 6 | 0.16 ± 0.00 c | 0.33 ± 0.02 a | 0.28 ± 0.02 a | 0.23 ± 0.02 b |
| Cannabisin C isomer 2 | 0.37 ± 0.03 a | 1.70 ± 0.10 a | 1.60 ± 0.05 a | 1.26 ± 0.03 b |
| Cannabisin C isomer 3 | 0.53 ± 0.04 a | 0.59 ± 0.04 a | 0.53 ± 0.03 a | 0.54 ± 0.00 a |
| Cannabisin C isomer 4 | 2.21 ± 0.10 b | 2.89 ± 0.11 a | 2.63 ± 0.07 a | 2.72 ± 0.10 a |
| Cannabisin D isomer 1 | 1.92 ± 0.02 a | 1.71 ± 0.05 b | 1.64 ± 0.10 b | 1.68 ± 0.04 b |
| Cannabisin B isomer 7 | 0.36 ± 0.02 c | 1.14 ± 0.04 a | 1.00 ± 0.04 ab | 0.85 ± 0.02 b |
| Cannabisin B isomer 8 | 0.15 ± 0.01 c | 0.47 ± 0.02 a | 0.45 ± 0.02 a | 0.26 ± 0.01 b |
| Cannabisin D isomer 2 | 1.03 ± 0.05 a | 1.01 ± 0.01 a | 0.99 ± 0.03 a | 0.98 ± 0.02 a |
| Cannabisin E isomer 1 | 0.82 ± 0.01 a | 0.61 ± 0.05 b | 0.68 ± 0.01 b | 0.56 ± 0.01 c |
| Cannabisin E isomer 2 | 0.41 ± 0.02 a | 0.39 ± 0.03 a | 0.38 ± 0.03 a | 0.32 ± 0.03 a |
| Cannabisin C isomer 5 | 0.31 ± 0.01 a | 0.32 ± 0.02 a | 0.28 ± 0.01 a | 0.25 ± 0.02 b |
| Cannabisin D isomer 3 | 1.62 ± 0.10 a | 1.29 ± 0.05 b | 1.29 ± 0.04 b | 1.3 ± 0.10 b |
| Cannabisin C isomer 6 | 0.39 ± 0.01 b | 0.44 ± 0.04 a | 0.36 ± 0.01 b | 0.30 ± 0.03 c |
| Cannabisin D isomer 4 | 0.34 ± 0.04 b | 0.45 ± 0.00 a | 0.40 ± 0.04 a | 0.15 ± 0.02 c |
| Cannabisin D isomer 5 | 1.12 ± 0.09 b | 1.28 ± 0.09 a | 1.37 ± 0.07 a | 0.79 ± 0.05 c |
| Total | 81 ± 5 c | 130 ± 6 a | 97 ± 5 b | 80 ± 4 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pontonio, E.; Verni, M.; Dingeo, C.; Diaz-de-Cerio, E.; Pinto, D.; Rizzello, C.G. Impact of Enzymatic and Microbial Bioprocessing on Antioxidant Properties of Hemp (Cannabis sativa L.). Antioxidants 2020, 9, 1258. https://doi.org/10.3390/antiox9121258
Pontonio E, Verni M, Dingeo C, Diaz-de-Cerio E, Pinto D, Rizzello CG. Impact of Enzymatic and Microbial Bioprocessing on Antioxidant Properties of Hemp (Cannabis sativa L.). Antioxidants. 2020; 9(12):1258. https://doi.org/10.3390/antiox9121258
Chicago/Turabian StylePontonio, Erica, Michela Verni, Cinzia Dingeo, Elixabet Diaz-de-Cerio, Daniela Pinto, and Carlo Giuseppe Rizzello. 2020. "Impact of Enzymatic and Microbial Bioprocessing on Antioxidant Properties of Hemp (Cannabis sativa L.)" Antioxidants 9, no. 12: 1258. https://doi.org/10.3390/antiox9121258
APA StylePontonio, E., Verni, M., Dingeo, C., Diaz-de-Cerio, E., Pinto, D., & Rizzello, C. G. (2020). Impact of Enzymatic and Microbial Bioprocessing on Antioxidant Properties of Hemp (Cannabis sativa L.). Antioxidants, 9(12), 1258. https://doi.org/10.3390/antiox9121258