Outcomes of Peptide Vaccine GV1001 Treatment in a Murine Model of Acute Noise-Induced Hearing Loss


 , , and
, , and 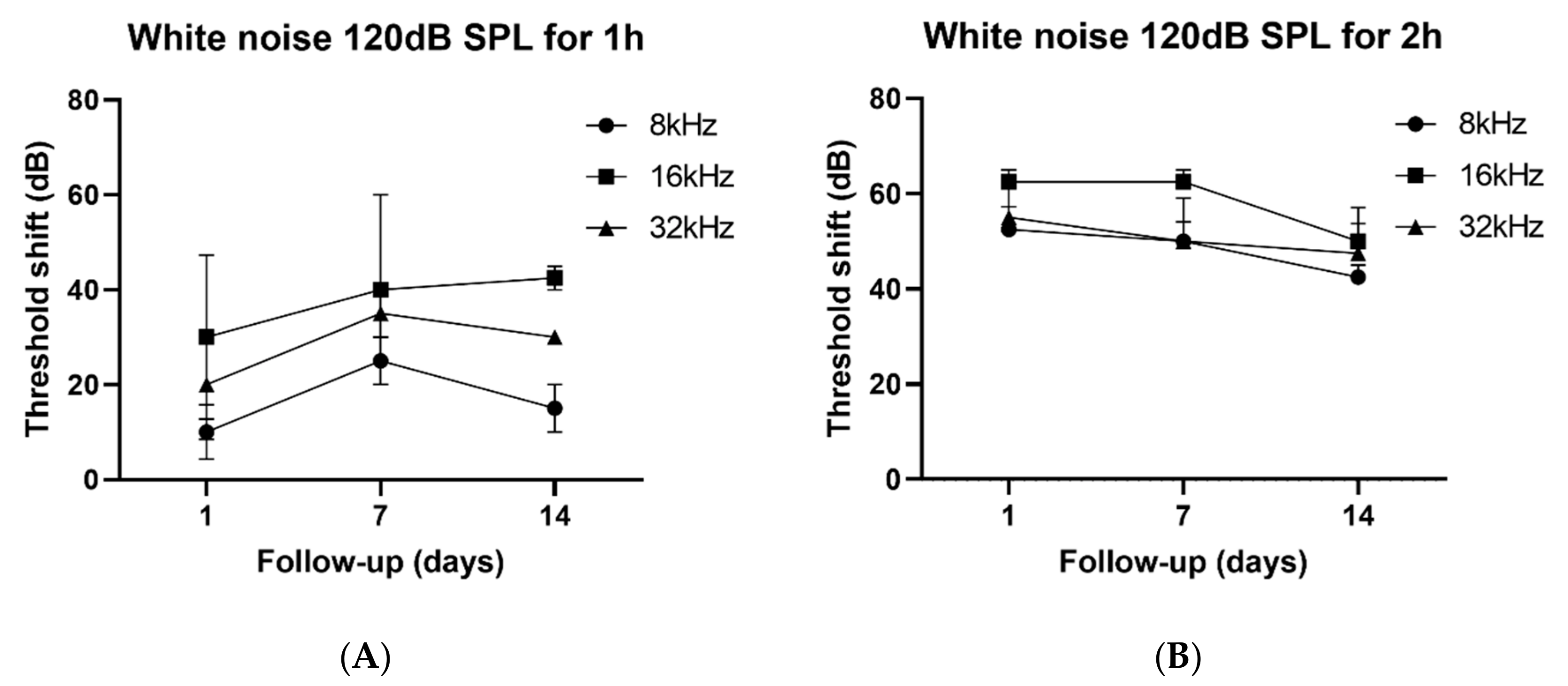{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. ABR Threshold Recording
2.3. Tissue Preparation
2.4. Statistical analysis
3. Results
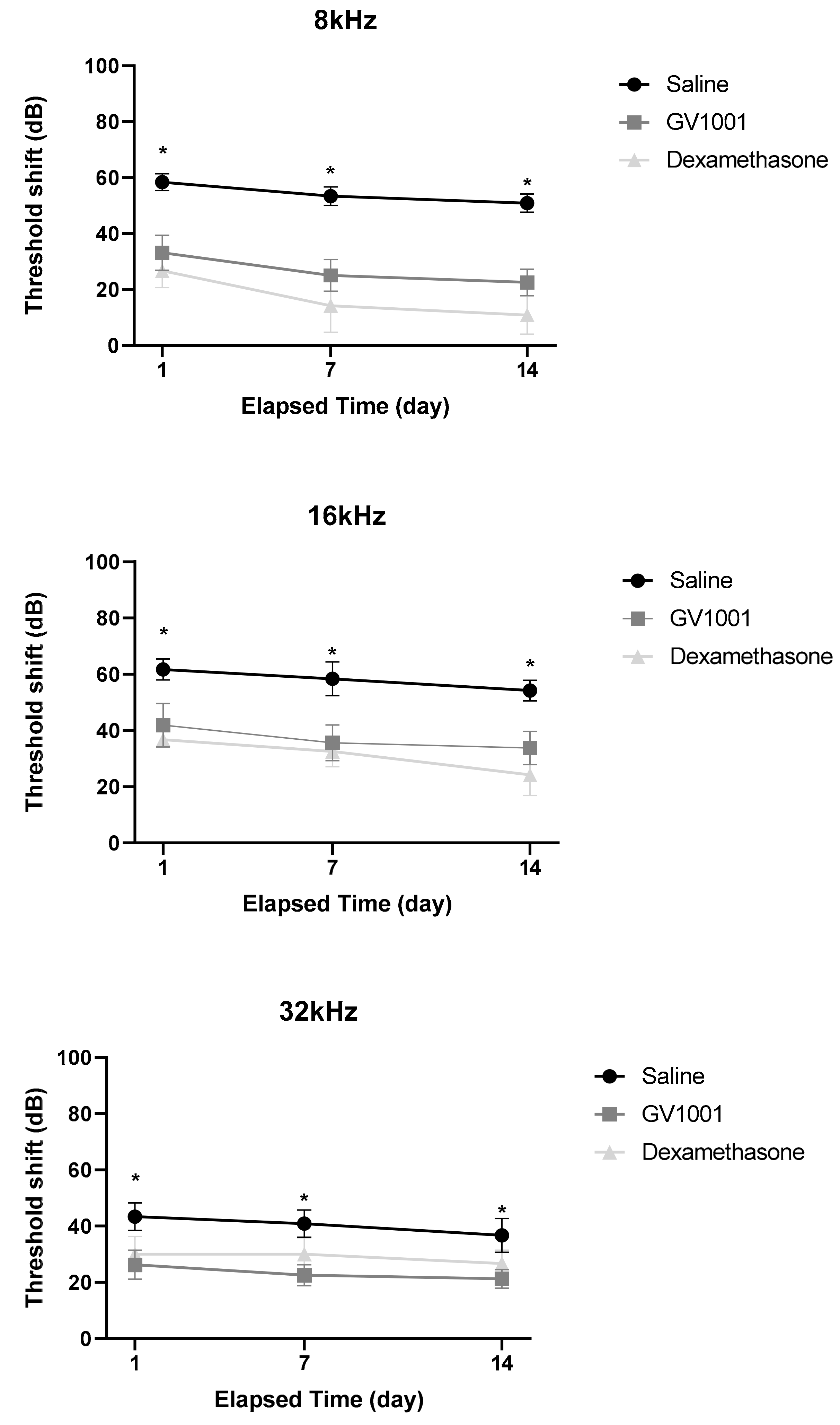
3.1. GV1001 Rescues Hearing Threshold Shifts After Severe PTS-Noise
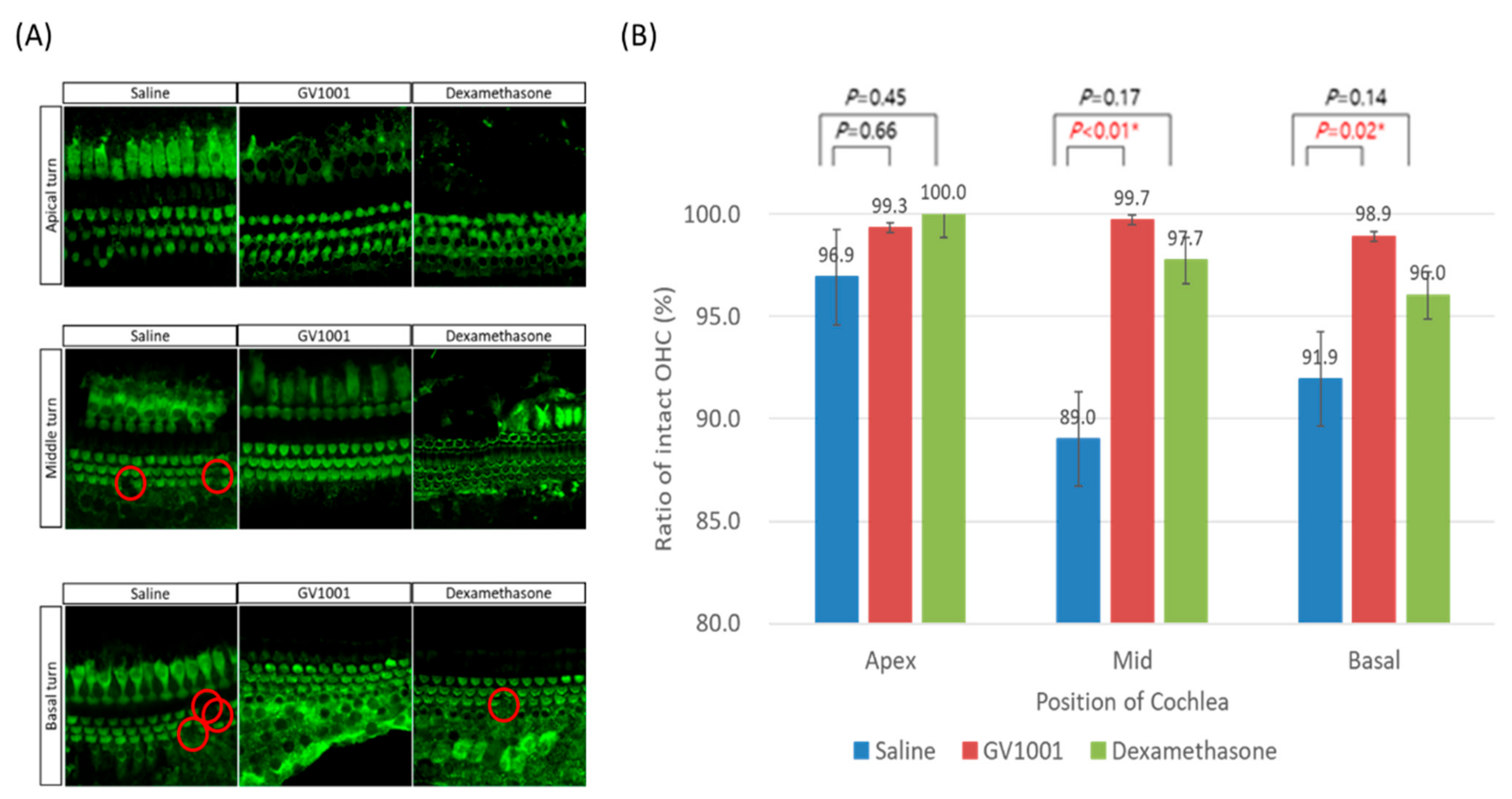
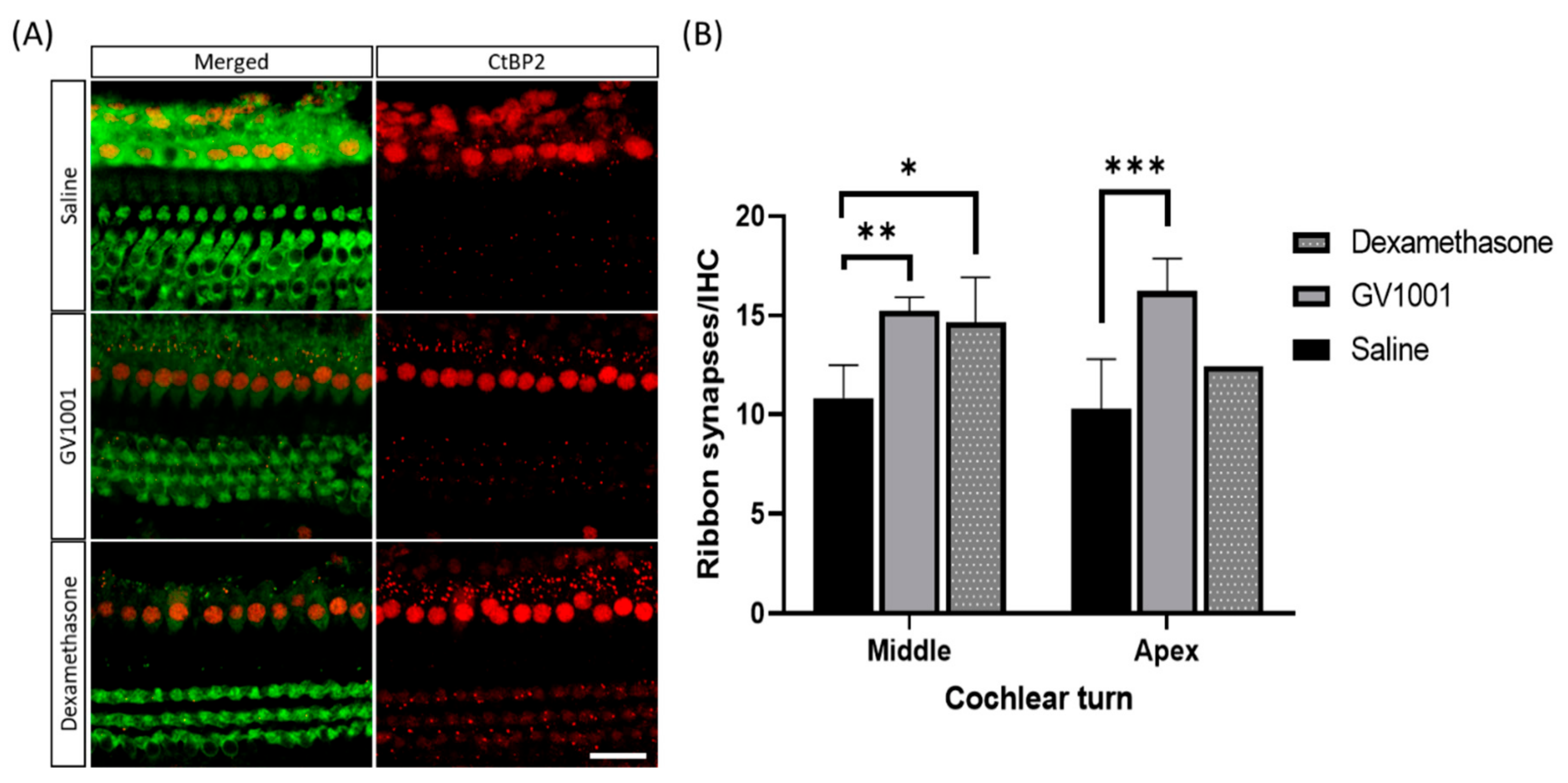
3.2. GV1001 Attenuates Hair Cell Damages Including Synaptic Ribbons
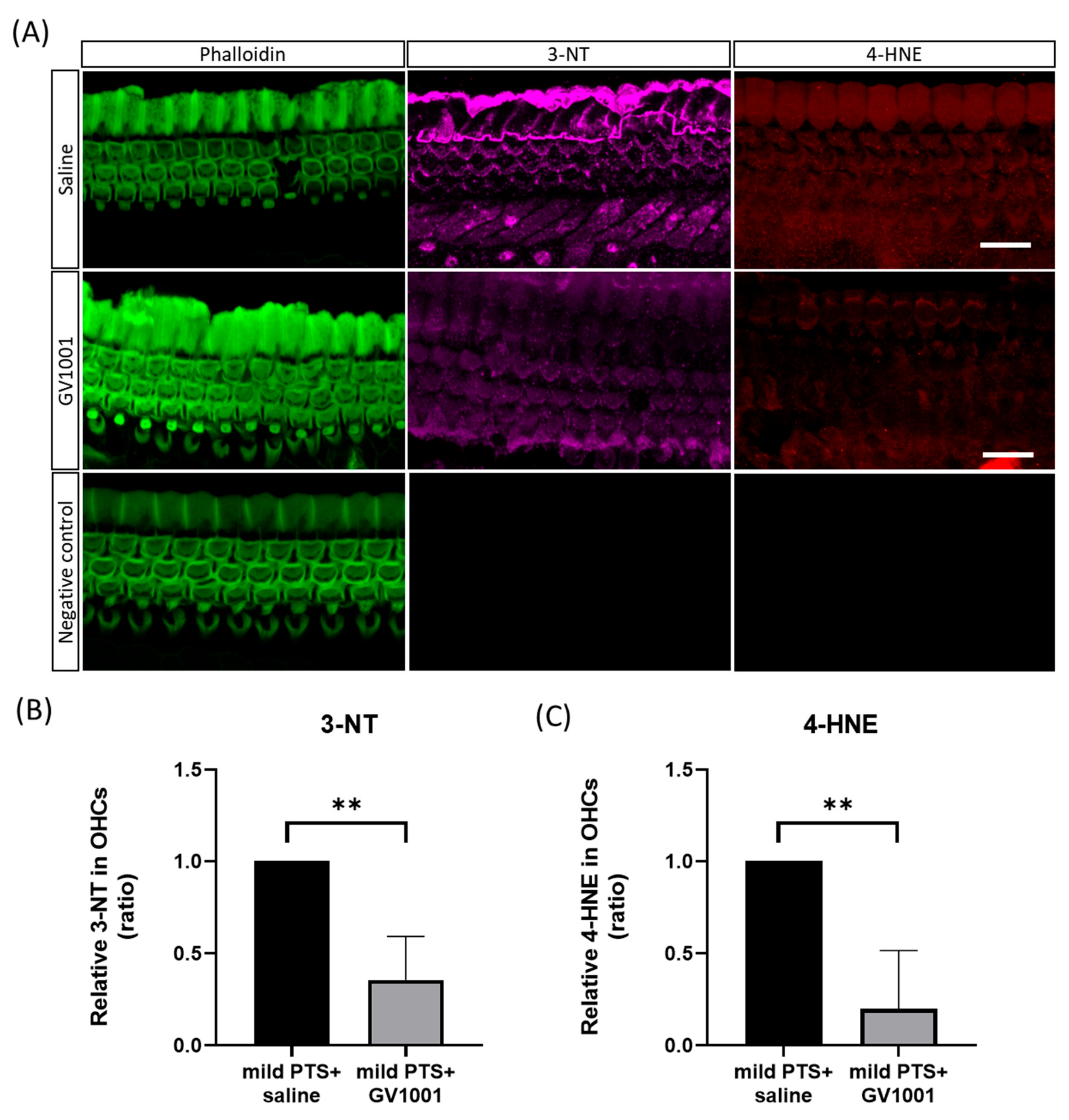
3.3. GV1001 Decreases the Expression of Oxidative Stress Markers
4. Discussion
4.1. Summary and Interpretation of the Results
4.2. Otoprotective Effect of GV1001 on NIHL and its Mechanism Associated with Oxidative Stress
4.3. PTS-Noise-Induced Cochlear Damage: Discrepancy between Morphology and Function
4.4. GV1001 Exhibits Comparable Protection with Dexamethasone against NIHL
4.5. Strengths and Limitations of the Current Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kujawa, S.G.; Liberman, M.C. Synaptopathy in the noise-exposed and aging cochlea: Primary neural degeneration in acquired sensorineural hearing loss. Hear. Res. 2015, 330, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, L.L.; Tucci, D.L. Restoring synaptic connections in the inner ear after noise damage. N. Engl. J. Med. 2015, 372, 181–182. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Wang, X.; Hill, K.; Chen, J.; Lemasters, J.; Yang, S.-M.; Sha, S.-H. Autophagy attenuates noise-induced hearing loss by reducing oxidative stress. Antioxid. Redox Signal. 2015, 22, 1308–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kujawa, S.G.; Liberman, M.C. Adding insult to injury: Cochlear nerve degeneration after “temporary” noise-induced hearing loss. J. Neurosci. 2009, 29, 14077–14085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Prell, C.G.; Yamashita, D.; Minami, S.B.; Yamasoba, T.; Miller, J.M. Mechanisms of noise-induced hearing loss indicate multiple methods of prevention. Hear. Res. 2007, 226, 22–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurabi, A.; Keithley, E.M.; Housley, G.D.; Ryan, A.F.; Wong, A.C. Cellular mechanisms of noise-induced hearing loss. Hear. Res. 2017, 349, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.E.; Kim, H.J.; Jheon, S.; Lim, C. Protective effects of GV1001 on myocardial ischemia-reperfusion injury. Mol. Med. Rep. 2017, 16, 7315–7320. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Kim, H.; Kim, Y.; Jang, M.; Jeon, J.; Hwang, Y.-i.; Shon, W.J.; Song, Y.W.; Kang, J.S.; Lee, W.J. The anti-inflammatory effect of GV1001 mediated by the downregulation of ENO1-induced pro-inflammatory cytokine production. Immune Netw. 2015, 15, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Park, H.-H.; Yu, H.-J.; Kim, S.; Kim, G.; Choi, N.-Y.; Lee, E.-H.; Lee, Y.J.; Yoon, M.-Y.; Lee, K.-Y.; Koh, S.-H. Neural stem cells injured by oxidative stress can be rejuvenated by GV1001, a novel peptide, through scavenging free radicals and enhancing survival signals. Neurotoxicology 2016, 55, 131–141. [Google Scholar] [CrossRef]
- Kyte, J.A. Cancer vaccination with telomerase peptide GV1001. Expert Opin. Investig. Drugs 2009, 18, 687–694. [Google Scholar] [CrossRef]
- Shaw, V.; Naisbitt, D.; Costello, E.; Greenhalf, W.; Park, B.; Neoptolemos, J.; Middleton, G. Current status of GV1001 and other telomerase vaccination strategies in the treatment of cancer. Expert Rev. Vaccines 2010, 9, 1007–1016. [Google Scholar] [CrossRef]
- Kim, S.H.; Jung, G.; Kim, S.; Koo, J.-W. Novel peptide vaccine GV1001 rescues hearing in kanamycin/furosemide-treated mice. Front. Cell. Neurosci. 2018, 12, 3. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Jung, G.; Shim, Y.J.; Koo, J.-W. The Novel Peptide Vaccine GV1001 Protects Hearing in a Kanamycin-induced Ototoxicity Mouse Model. Otol. Neurotol. 2018, 39, e731–e737. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Park, S.K.; Cho, Y.-S.; Lee, H.-S.; Kim, K.R.; Kim, M.G.; Chung, W.-H. Gentamicin induced nitric oxide-related oxidative damages on vestibular afferents in the guinea pig. Hear. Res. 2006, 211, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Umugire, A.; Lee, S.; Kim, D.; Choi, M.; Kim, H.-S.; Cho, H.-H. Avenanthramide-C prevents noise-and drug-induced hearing loss while protecting auditory hair cells from oxidative stress. Cell Death Discov. 2019, 5, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.A.; Back, S.A.; Kim, H.L.; Park, S.Y.; Yeo, S.W.; Park, S.N. Therapeutic effect of dexamethasone for noise-induced hearing loss: Systemic versus intratympanic injection in mice. Otol. Neurotol. 2015, 36, 755–762. [Google Scholar] [CrossRef]
- Waqas, M.; Gao, S.; Ali, M.K.; Ma, Y.; Li, W. Inner Ear Hair Cell Protection in Mammals against the Noise-Induced Cochlear Damage. Neural Plast. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Gumrukcu, S.S.; Topaloglu, İ.; Salturk, Z.; Tutar, B.; Atar, Y.; Berkiten, G.; Göker, A.E. Effects of intratympanic dexamethasone on noise-induced hearing loss: An experimental study. Am. J. Otolaryngol. 2018, 39, 71–73. [Google Scholar] [CrossRef]
- Yamane, H.; Nakai, Y.; Takayama, M.; Iguchi, H.; Nakagawa, T.; Kojima, A. Appearance of free radicals in the guinea pig inner ear after noise-induced acoustic trauma. Eur. Arch Otorhinolaryngol. 1995, 252, 504–508. [Google Scholar] [CrossRef]
- Choung, Y.; Taura, A.; Pak, K.; Choi, S.; Masuda, M.; Ryan, A. Generation of highly-reactive oxygen species is closely related to hair cell damage in rat organ of Corti treated with gentamicin. Neuroscience 2009, 161, 214–226. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.-Q.; Zheng, H.-W.; Hill, K.; Sha, S.-H. Traumatic noise activates Rho-family GTPases through transient cellular energy depletion. J. Neurosci. 2012, 32, 12421–12430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.K.; Nata’atmaja, B.S.; Kim, B.H.; Pak, C.S.; Heo, C.Y. Protective effect of telomerase-based 16-mer peptide vaccine (GV1001) on inferior epigastric island skin flap survivability in ischaemia-reperfusion injury rat model. J. Plast. Surg. Hand Surg. 2017, 51, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-H.; Lee, K.-Y.; Kim, S.; Lee, J.W.; Choi, N.-Y.; Lee, E.-H.; Lee, Y.J.; Lee, S.-H.; Koh, S.-H. The novel vaccine peptide GV1001 effectively blocks β-amyloid toxicity by mimicking the extra-telomeric functions of human telomerase reverse transcriptase. Neurobiol. Aging 2014, 35, 1255–1274. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-X.; Zhu, G.-X.; Liu, X.-Q.; Sun, F.; Zhou, K.; Wang, S.; Wang, C.-M.; Jia, J.-W.; Song, J.-T.; Lu, L.-J. Noise alters guinea pig’s blood-labyrinth barrier ultrastructure and permeability along with a decrease of cochlear Claudin-5 and Occludin. BMC Neuroscience 2014, 15, 136. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Choi, C.-H.; Chen, K.; Cheng, W.; Floyd, R.A.; Kopke, R.D. Reduced formation of oxidative stress biomarkers and migration of mononuclear phagocytes in the cochleae of chinchilla after antioxidant treatment in acute acoustic trauma. Int. J. Otolaryngol. 2011, 2011. [Google Scholar] [CrossRef]
- Henderson, D.; Hu, B.; Bielefeld, E. Patterns and mechanisms of noise-induced cochlear pathology. In Auditory Trauma, Protection, and Repair; Springer: Boston, MA, USA, 2008; pp. 195–217. [Google Scholar]
- Sha, S.-H.; Taylor, R.; Forge, A.; Schacht, J. Differential vulnerability of basal and apical hair cells is based on intrinsic susceptibility to free radicals. Hear. Res. 2001, 155, 1–8. [Google Scholar] [CrossRef]
- Paciello, F.; Fetoni, A.; Rolesi, R.; Wright, M.B.; Grassi, C.; Troiani, D.; Paludetti, G. Pioglitazone represents an effective therapeutic target in preventing oxidative/inflammatory cochlear damage induced by noise exposure. Front. Pharmacol. 2018, 9, 1103. [Google Scholar] [CrossRef] [Green Version]
- Muller, M.; von Hunerbein, K.; Hoidis, S.; Smolders, J.W. A physiological place-frequency map of the cochlea in the CBA/J mouse. Hear. Res. 2005, 202, 63–73. [Google Scholar] [CrossRef]
- Hu, B.H.; Zheng, X.Y.; McFadden, S.L.; Kopke, R.D.; Henderson, D. R-phenylisopropyladenosine attenuates noise-induced hearing loss in the chinchilla. Hear. Res. 1997, 113, 198–206. [Google Scholar] [CrossRef]
- Engström, B.; Flock, Å.; Borg, E. Ultrastructural studies of stereocilia in noise-exposed rabbits. Hear. Res. 1983, 12, 251–264. [Google Scholar] [CrossRef]
- Khimich, D.; Nouvian, R.; Pujol, R.; Tom Dieck, S.; Egner, A.; Gundelfinger, E.D.; Moser, T. Hair cell synaptic ribbons are essential for synchronous auditory signalling. Nature 2005, 434, 889. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Omelchenko, I.; Shi, X.; Nuttall, A.L. The influence of NF-κB signal-transduction pathways on the murine inner ear by acoustic overstimulation. J. Neurosci. Res. 2009, 87, 1832–1840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milon, B.; Mitra, S.; Song, Y.; Margulies, Z.; Casserly, R.; Drake, V.; Mong, J.A.; Depireux, D.A.; Hertzano, R. The impact of biological sex on the response to noise and otoprotective therapies against acoustic injury in mice. Biol. Sex Differ. 2018, 9, 12. [Google Scholar] [CrossRef]
- Park, S.-N.; Back, S.-A.; Park, K.-H.; Seo, J.-H.; Noh, H.-I.; Akil, O.; Lustig, L.R.; Yeo, S.W. Comparison of functional and morphologic characteristics of mice models of noise-induced hearing loss. Auris Nasus Larynx 2013, 40, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Fridberger, A.; Flock, Å.; Ulfendahl, M.; Flock, B. Acoustic overstimulation increases outer hair cell Ca2+ concentrations and causes dynamic contractions of the hearing organ. Proc. Natl. Acad. Sci. USA 1998, 95, 7127–7132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhu, Y.; Long, H.; Pan, S.; Xiong, H.; Fang, Q.; Hill, K.; Lai, R.; Yuan, H.; Sha, S.-H. Mitochondrial calcium transporters mediate sensitivity to noise-induced hearing loss and cochlear synapses. Front. Mol. Neurosci. 2018, 11, 469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo, C.; Donoso, P. Crosstalk between calcium and redox signaling: From molecular mechanisms to health implications. Antioxid. Redox Signal. 2008, 10, 1275–1312. [Google Scholar] [CrossRef]
- Vlajkovic, S.M.; Lin, S.C.-Y.; Wong, A.C.Y.; Wackrow, B.; Thorne, P.R. Noise-induced changes in expression levels of NADPH oxidases in the cochlea. Hear. Res. 2013, 304, 145–152. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-Y.; Han, J.J.; Lee, S.-Y.; Jung, G.; Min, H.J.; Song, J.-J.; Koo, J.-W. Outcomes of Peptide Vaccine GV1001 Treatment in a Murine Model of Acute Noise-Induced Hearing Loss. Antioxidants 2020, 9, 112. https://doi.org/10.3390/antiox9020112
Lee S-Y, Han JJ, Lee S-Y, Jung G, Min HJ, Song J-J, Koo J-W. Outcomes of Peptide Vaccine GV1001 Treatment in a Murine Model of Acute Noise-Induced Hearing Loss. Antioxidants. 2020; 9(2):112. https://doi.org/10.3390/antiox9020112
Chicago/Turabian StyleLee, Sang-Yeon, Jae Joon Han, Sang-Youp Lee, Gaon Jung, Hyun Jin Min, Jae-Jin Song, and Ja-Won Koo. 2020. "Outcomes of Peptide Vaccine GV1001 Treatment in a Murine Model of Acute Noise-Induced Hearing Loss" Antioxidants 9, no. 2: 112. https://doi.org/10.3390/antiox9020112
APA StyleLee, S. -Y., Han, J. J., Lee, S. -Y., Jung, G., Min, H. J., Song, J. -J., & Koo, J. -W. (2020). Outcomes of Peptide Vaccine GV1001 Treatment in a Murine Model of Acute Noise-Induced Hearing Loss. Antioxidants, 9(2), 112. https://doi.org/10.3390/antiox9020112