Multidirectional Pharma-Toxicological Study on Harpagophytum procumbens DC. ex Meisn.: An IBD-Focused Investigation



 ,
,  ,
, 


 , ,
, ,  , ,
, , 


Abstract
:1. Introduction
2. Materials and Methods
2.1. Pharmacognostic Studies
2.1.1. Plant Material and Extraction Procedure
2.1.2. Phytochemical Profile
2.2. Toxicological, Pharmacological, and Microbiological Studies
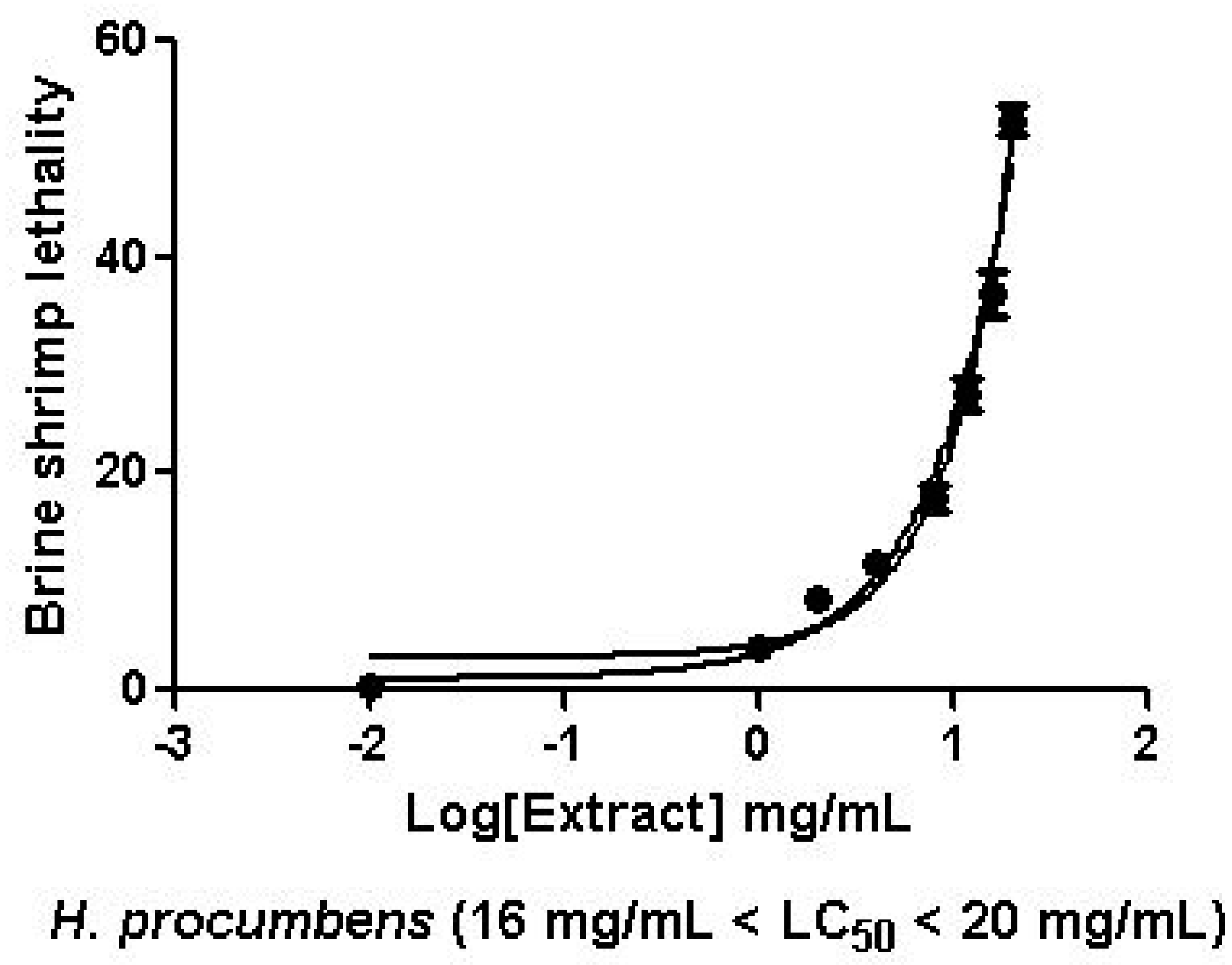
2.2.1. Artemia Salina Lethality Bioassay
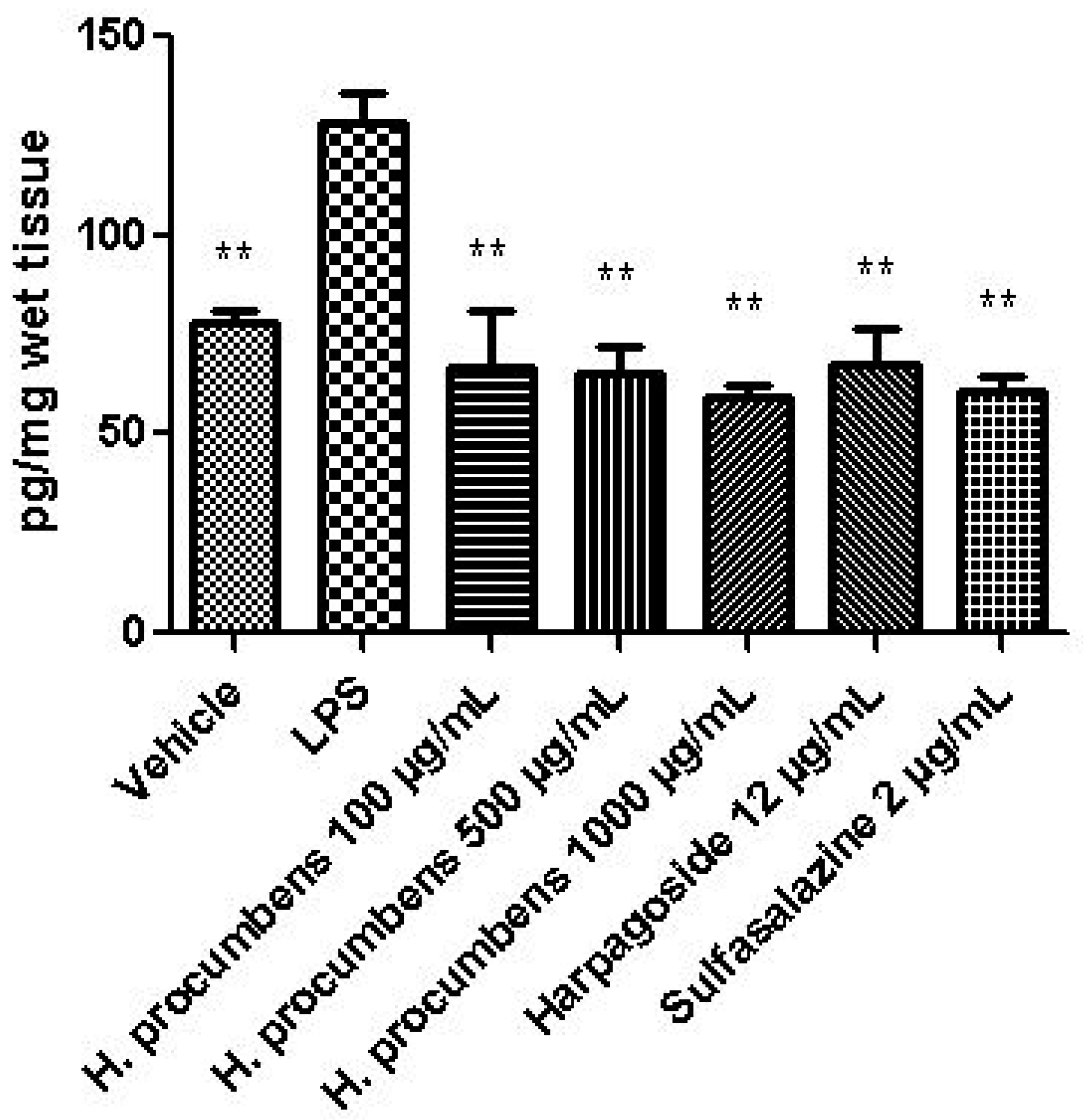
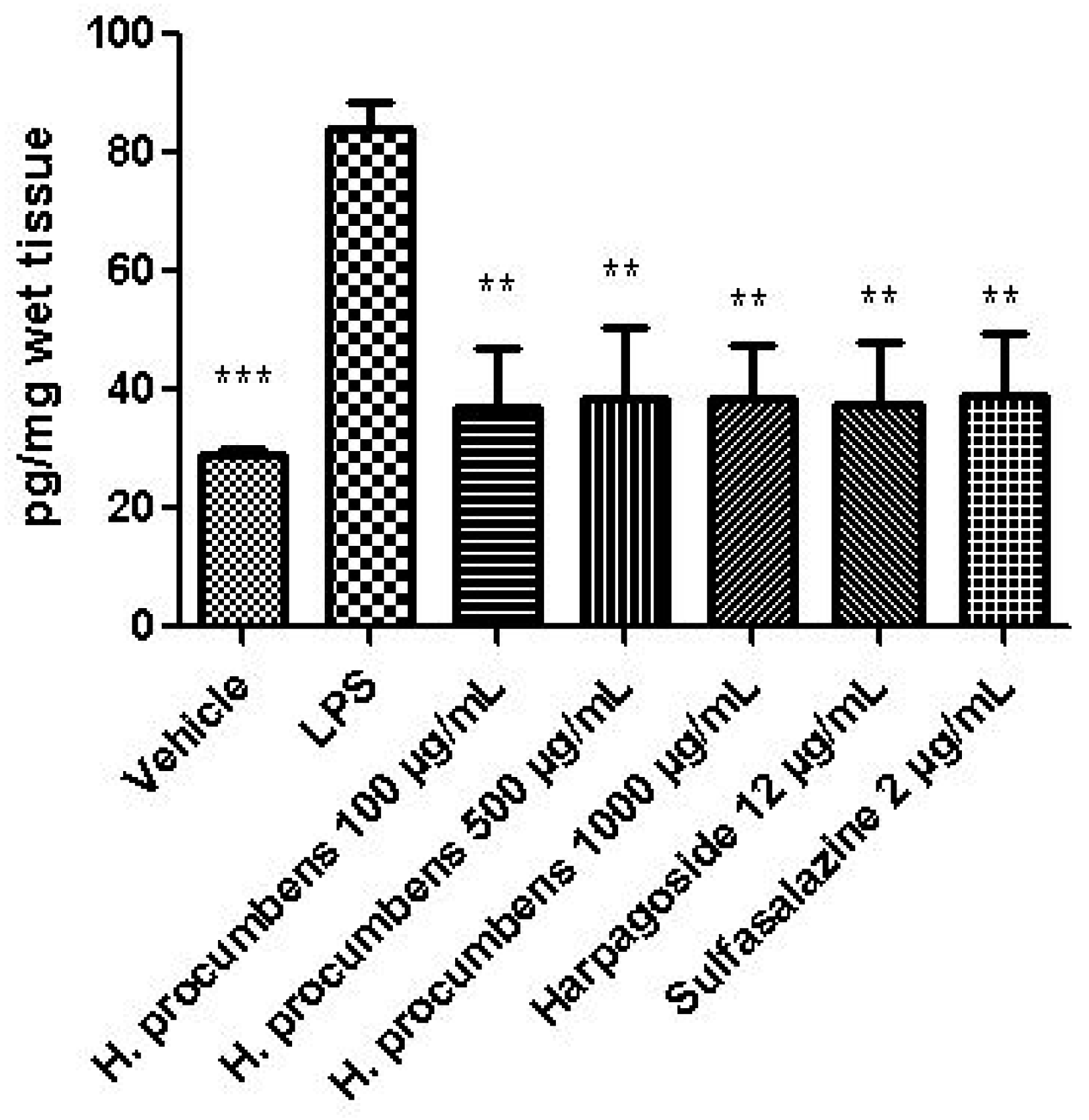
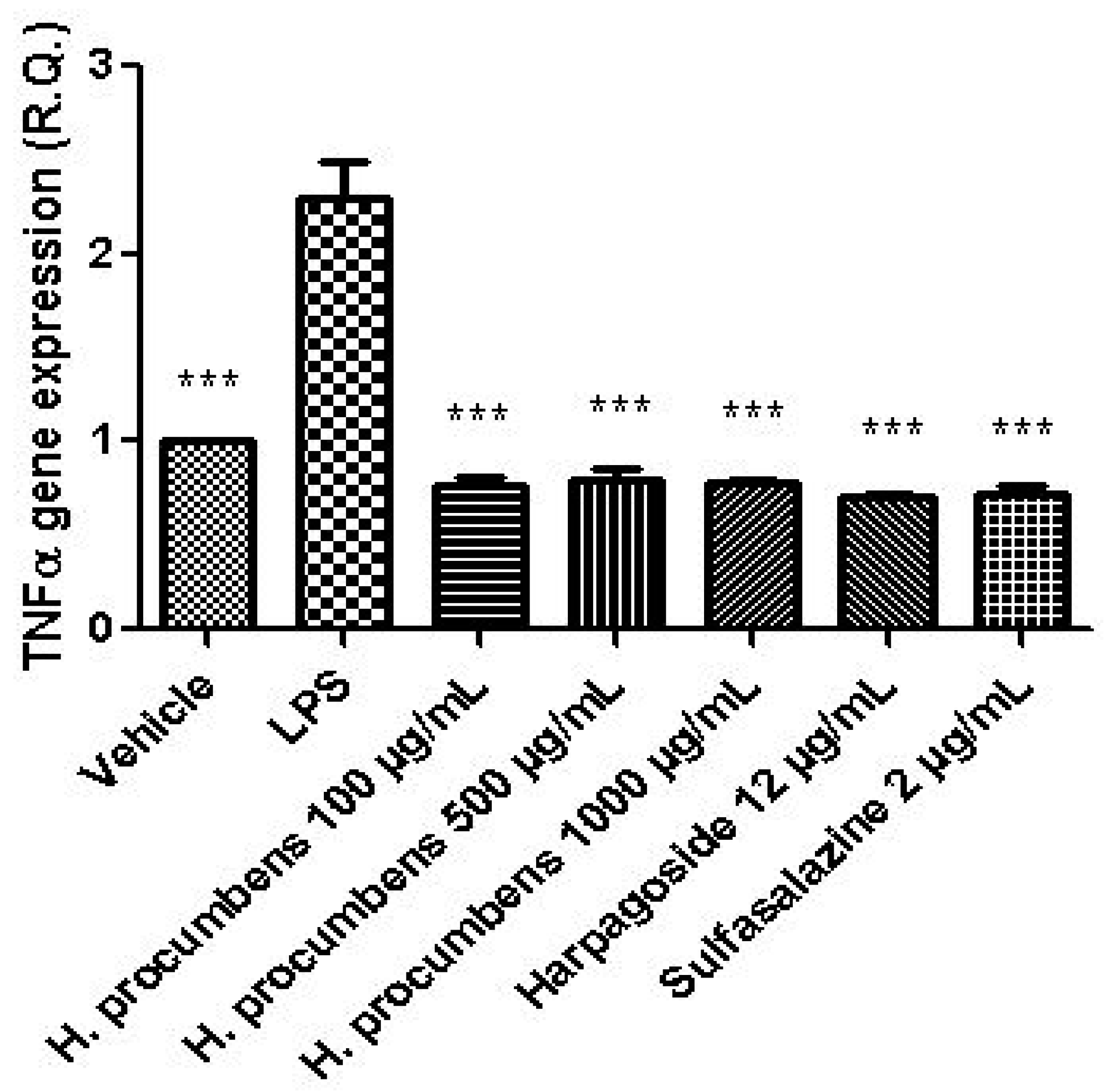
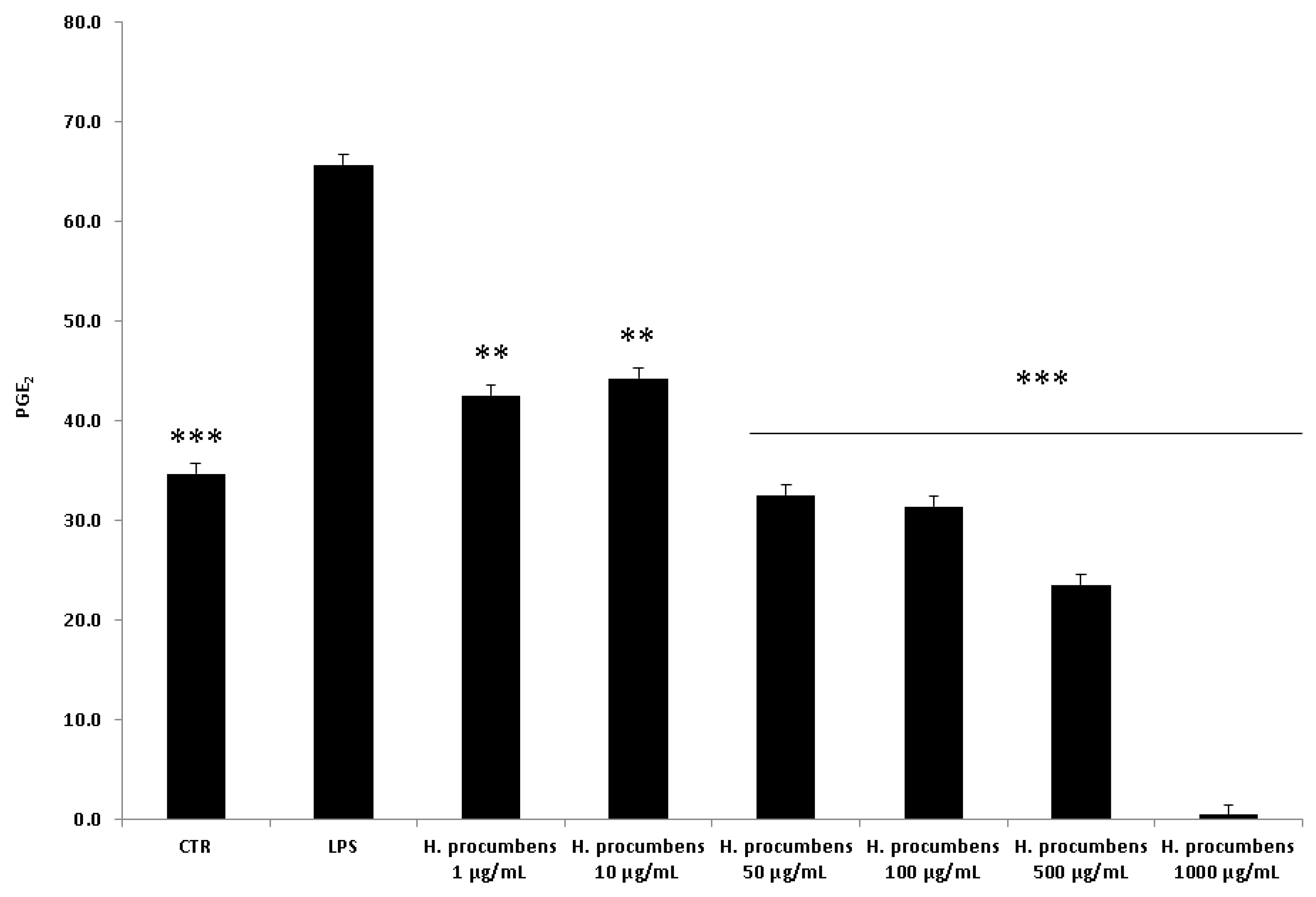
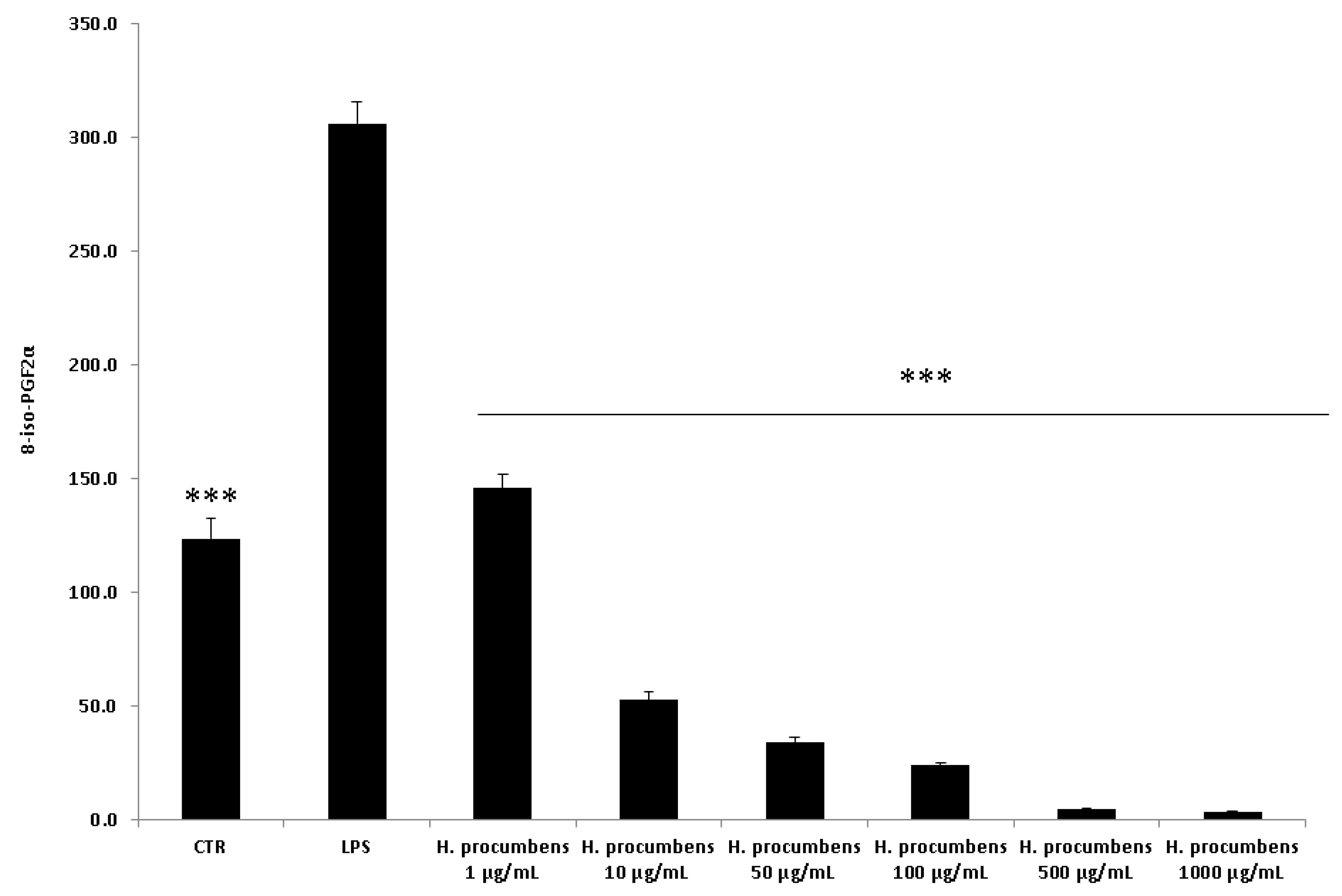
2.2.2. Ex Vivo Studies
2.2.3. Antimicrobial Susceptibility Testing
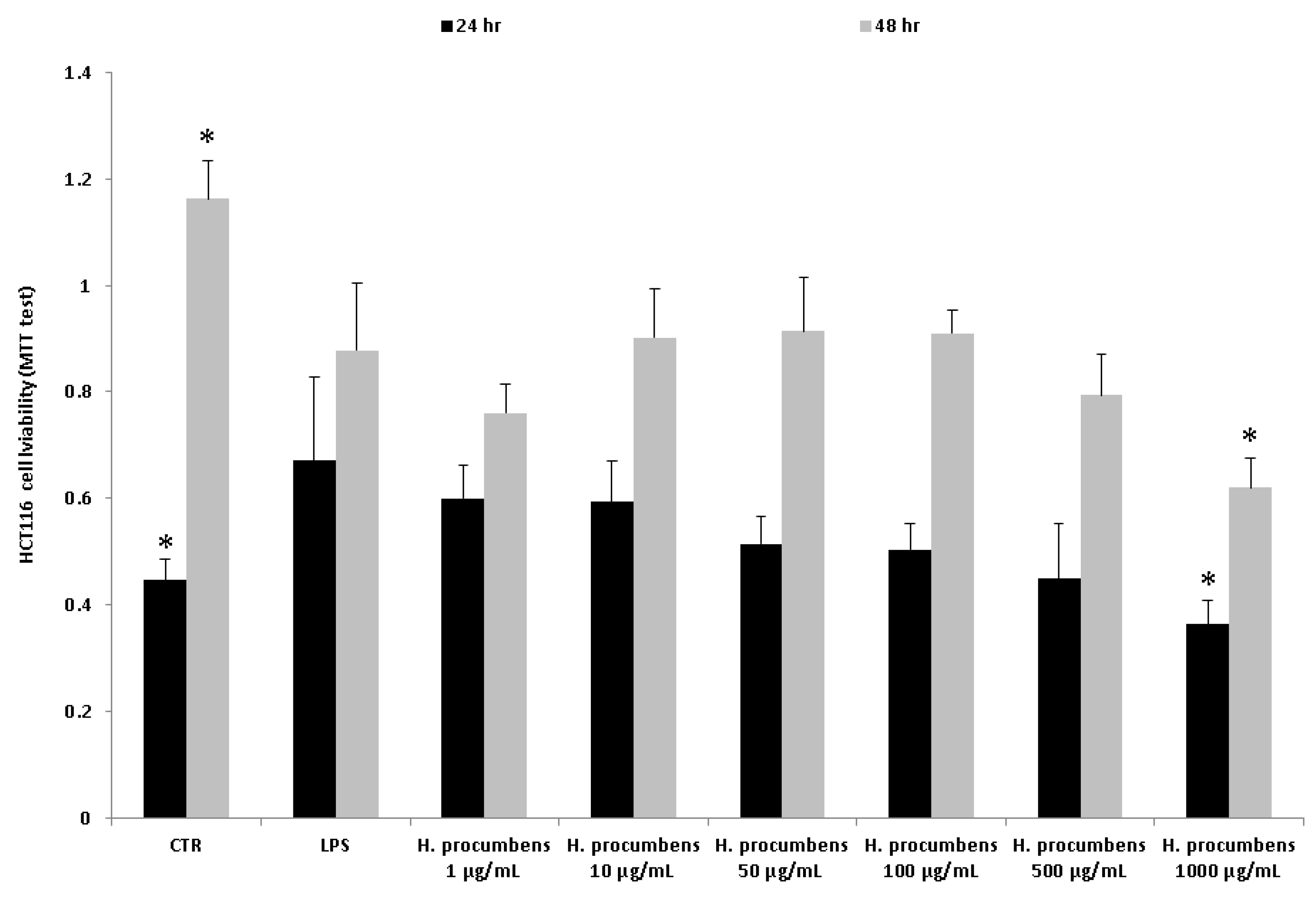
2.2.4. Human Colon Cancer HCT116 Cell Culture
2.3. Statistical Analysis
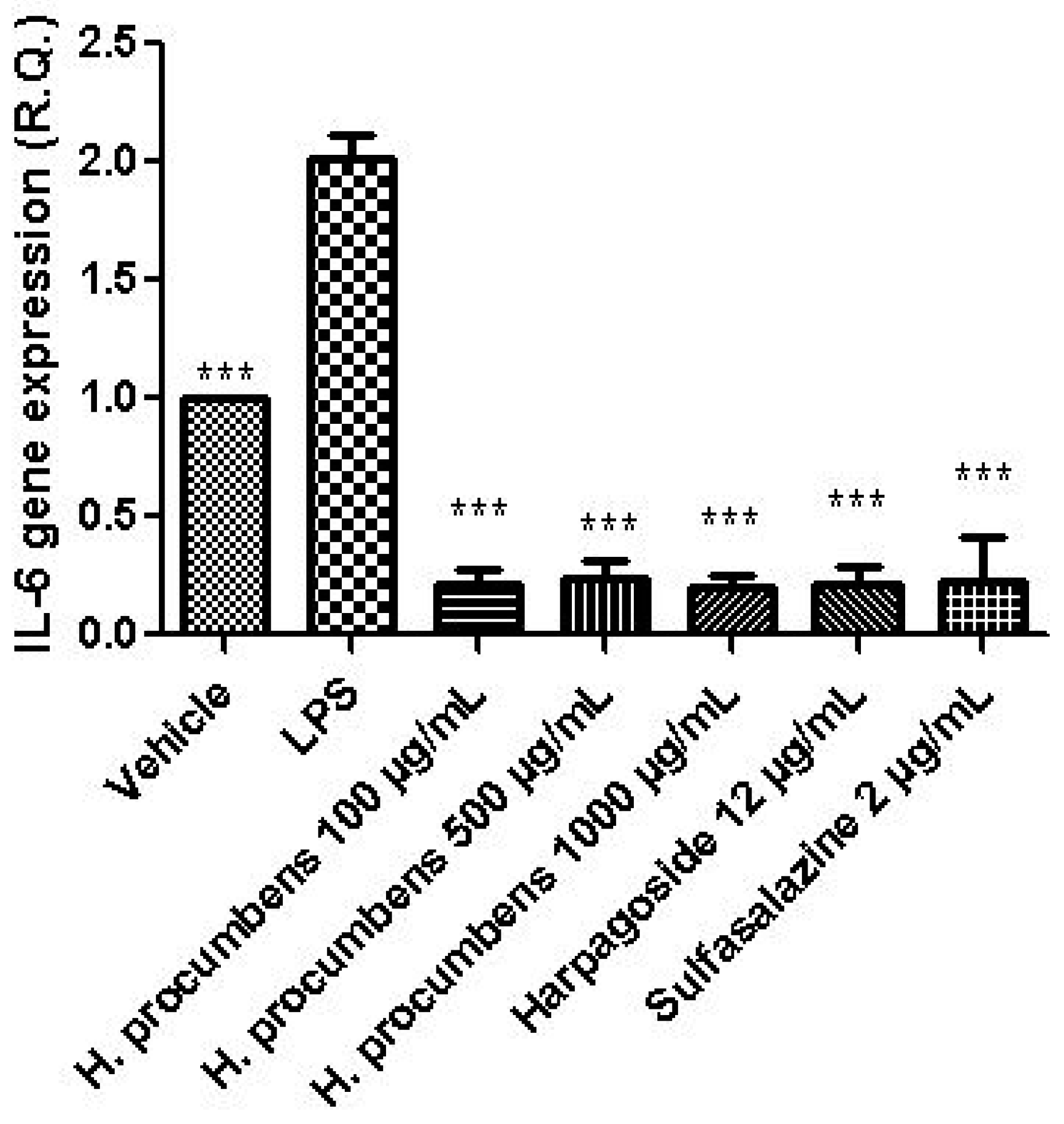
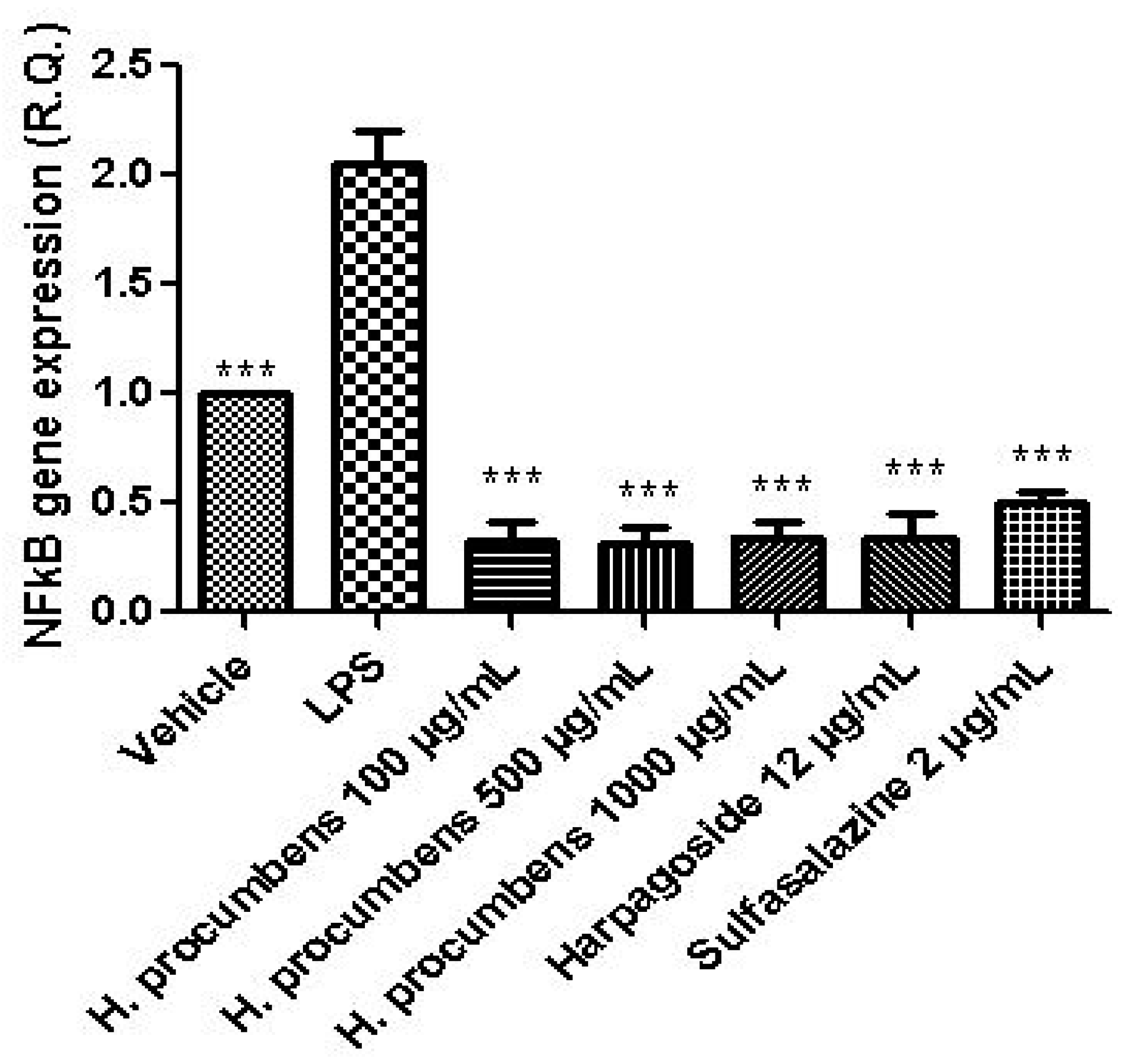
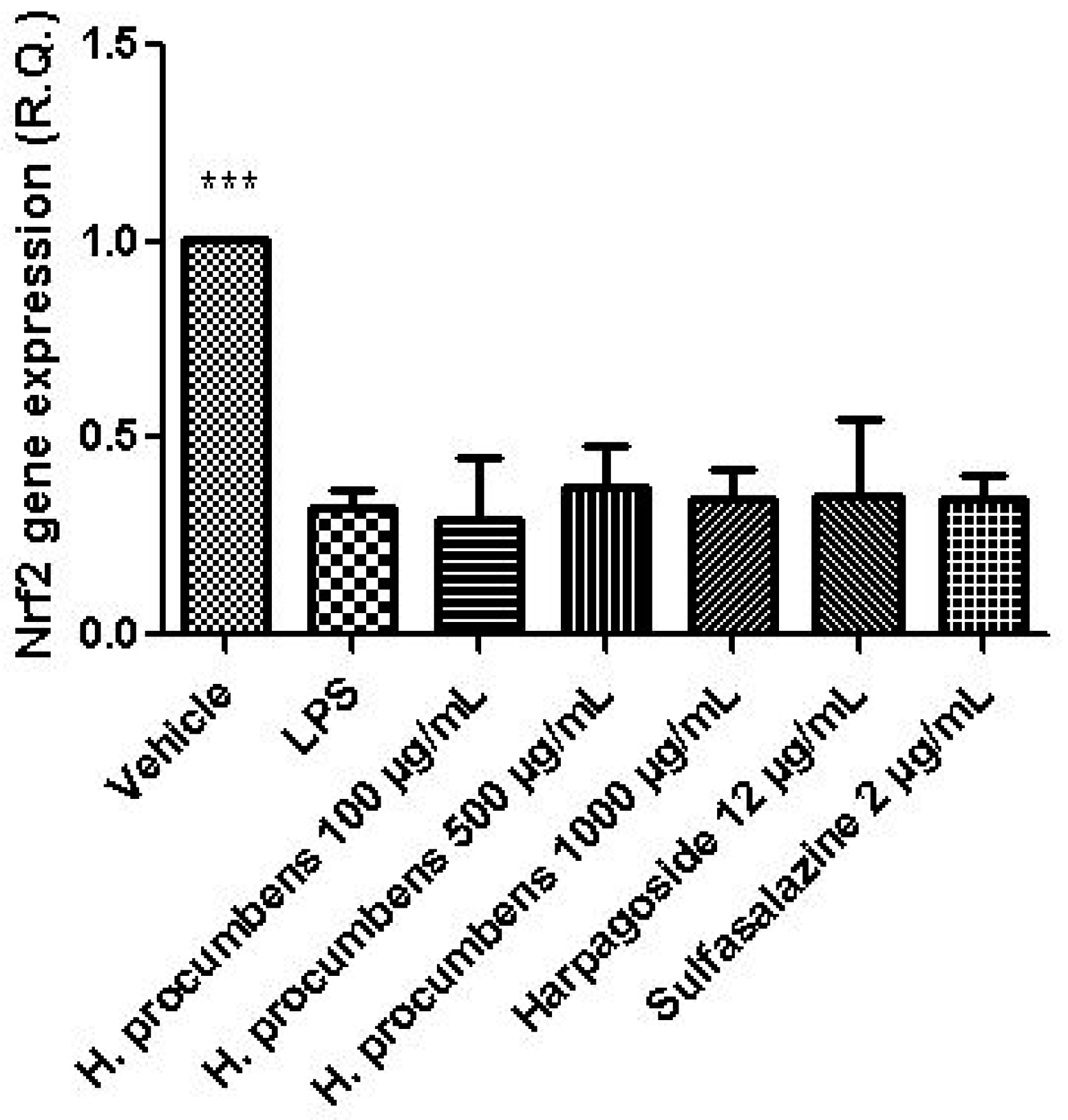
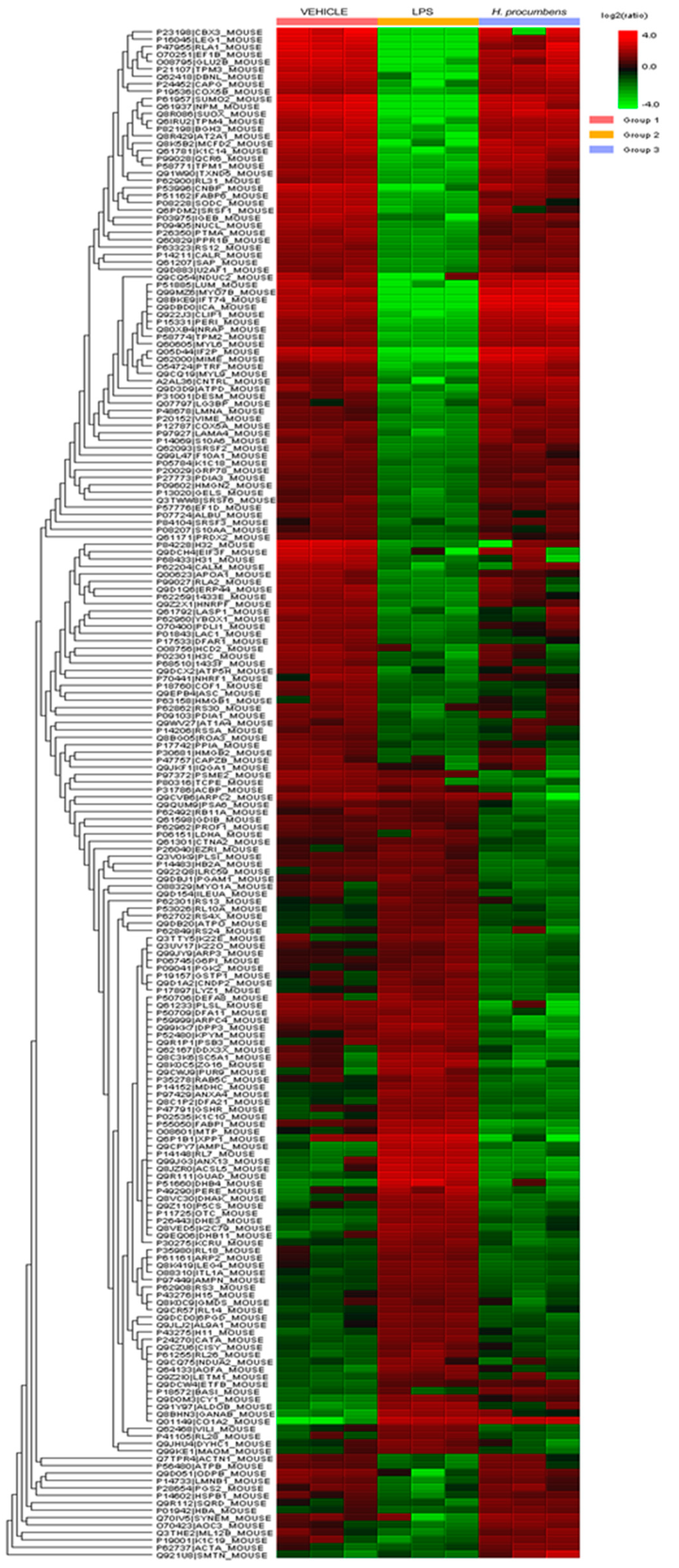
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ARPC4 | actin-related protein 2/3 complex subunit 4 |
| BHT | butylated hydroxytoluene |
| CAPG | macrophage-capping protein |
| CATA | catalase |
| DBNL | drebrin-like protein |
| DEFA8 | α-defensin 8 |
| DEFA11 | α-defensin 11 |
| DPPH | 1,1-diphenyl-2-picrylhydrazyl |
| EZRI | proteins namely ezrin |
| GSHR | glutathione reductase |
| HGMB2 | high mobility group protein B2 |
| 5-HT | 5-hydroxytryptamine, serotonin |
| IBDs | inflammatory bowel diseases |
| IL-6 | interleukin-6 |
| LPS | lipopolysaccharide |
| NFκB | nuclear factor kappa B |
| Nrf2 | nuclear factor erythroid 2-related factor 2 |
| PGE2 | prostaglandin E2 |
| 8-iso-PGF2α | 8-iso-prostaglandin-F2α |
| PLSI | plastin-1 |
| PRDX2 | peroxiredoxin-2 |
| PTMA | prothymosin alpha |
| ROS/RNS | reactive oxygen/nitrogen species |
| SMTN | smoothelin |
| SODC | superoxide dismutase |
| TNFα | tumor necrosis factor α |
References
- Achitei, D.; Ciobica, A.; Balan, G.; Gologan, E.; Stanciu, C.; Stefanescu, G. Different profile of peripheral antioxidant enzymes and lipid peroxidation in active and non-active inflammatory bowel disease patients. Dig. Dis. Sci. 2013, 58, 1244–1249. [Google Scholar] [CrossRef] [PubMed]
- Koutroubakis, I.E.; Malliaraki, N.; Dimoulios, P.D.; Karmiris, K.; Castanas, E.; Kouroumalis, E.A. Decreased total and corrected antioxidant capacity in patients with inflammatory bowel disease. Dig. Dis. Sci. 2004, 49, 1433–1437. [Google Scholar] [CrossRef] [PubMed]
- Rezaie, A.; Parker, R.D.; Abdollahi, M. Oxidative stress and pathogenesis of inflammatory bowel disease: An epiphenomenon or the cause? Dig. Dis. Sci. 2007, 52, 2015–2021. [Google Scholar] [CrossRef] [PubMed]
- Strober, W.; Fuss, I.; Mannon, P. The fundamental basis of inflammatory bowel disease. J. Clin. Investig. 2007, 117, 514–521. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.H.; Moon, W.; Kim, Y.S.; Kim, E.S.; Lee, B.-I.; Jung, Y.; Yoon, Y.S.; Lee, H.; Park, D.I.; Han, D.S. Second Korean guidelines for the management of ulcerative colitis. Intest. Res. 2017, 15, 7. [Google Scholar] [CrossRef]
- Chung, H.-L.; Yue, G.G.-L.; To, K.-F.; Su, Y.-L.; Huang, Y.; Ko, W.-H. Effect of Scutellariae Radix extract on experimental dextran-sulfate sodium-induced colitis in rats. World J. Gastroenterol. WJG 2007, 13, 5605. [Google Scholar] [CrossRef] [Green Version]
- Lenoir, L.; Joubert-Zakeyh, J.; Texier, O.; Lamaison, J.L.; Vasson, M.P.; Felgines, C. Aloysia triphylla infusion protects rats against dextran sulfate sodium-induced colonic damage. J. Sci. Food Agric. 2012, 92, 1570–1572. [Google Scholar] [CrossRef]
- Locatelli, M.; Ferrante, C.; Carradori, S.; Secci, D.; Leporini, L.; Chiavaroli, A.; Leone, S.; Recinella, L.; Orlando, G.; Martinotti, S. Optimization of aqueous extraction and biological activity of Harpagophytum procumbens root on ex vivo rat colon inflammatory model. Phytother. Res. 2017, 31, 937–944. [Google Scholar] [CrossRef]
- Bettenworth, D.; Nowacki, T.M.; Friedrich, A.; Becker, K.; Wessling, J.; Heidemann, J. Crohn’s disease complicated by intestinal infection with methicillin-resistant Staphylococcus aureus. World J. Gastroenterol. WJG 2013, 19, 4418. [Google Scholar] [CrossRef]
- Iguidbashian, J.P.; Parekh, J.D.; Kukrety, S.; Andukuri, V.G. Campylobacter jejuni and Pseudomonas coinfection in the setting of ulcerative colitis. Case Rep. 2018, 2018, bcr-2018. [Google Scholar]
- Iliev, I.D.; Funari, V.A.; Taylor, K.D.; Nguyen, Q.; Reyes, C.N.; Strom, S.P.; Brown, J.; Becker, C.A.; Fleshner, P.R.; Dubinsky, M. Interactions between commensal fungi and the C-type lectin receptor Dectin-1 influence colitis. Science 2012, 336, 1314–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trojanowska, D.; Zwolinska-Wcislo, M.; Tokarczyk, M.; Kosowski, K.; Mach, T.; Budak, A. The role of Candida in inflammatory bowel disease. Estimation of transmission of C. albicans fungi in gastrointestinal tract based on genetic affinity between strains. Med. Sci. Monit. 2010, 16, CR451–CR457. [Google Scholar] [PubMed]
- Zengin, G.; Menghini, L.; Malatesta, L.; De Luca, E.; Bellagamba, G.; Uysal, S.; Aktumsek, A.; Locatelli, M. Comparative study of biological activities and multicomponent pattern of two wild Turkish species: Asphodeline anatolica and Potentilla speciosa. J. Enzym. Inhib. Med. Chem. 2016, 31 (Suppl. S1), 203–208. [Google Scholar] [CrossRef] [Green Version]
- Rodrıguez-Delgado, M.; Malovana, S.; Perez, J.; Borges, T.; Montelongo, F.G. Separation of phenolic compounds by high-performance liquid chromatography with absorbance and fluorimetric detection. J. Chromatogr. A 2001, 912, 249–257. [Google Scholar] [CrossRef]
- Recinella, L.; Chiavaroli, A.; Orlando, G.; Menghini, L.; Ferrante, C.; Di Cesare Mannelli, L.; Ghelardini, C.; Brunetti, L.; Leone, S. Protective Effects Induced by Two Polyphenolic Liquid Complexes from Olive (Olea europaea, mainly Cultivar Coratina) Pressing Juice in Rat Isolated Tissues Challenged with LPS. Molecules 2019, 24, 3002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiavaroli, A.; Recinella, L.; Ferrante, C.; Locatelli, M.; Carradori, S.; Macchione, N.; Zengin, G.; Leporini, L.; Leone, S.; Martinotti, S. Crocus sativus, Serenoa repens and Pinus massoniana extracts modulate inflammatory response in isolated rat prostate challenged with LPS. J. Biol. Regul. Homeost. Agents 2017, 31, 531–541. [Google Scholar] [PubMed]
- Ferrante, C.; Orlando, G.; Recinella, L.; Leone, S.; Chiavaroli, A.; Di, C.N.; Shohreh, R.; Manippa, F.; Ricciuti, A.; Vacca, M. Central apelin-13 administration modulates hypothalamic control of feeding. J. Biol. Regul. Homeost. Agents 2016, 30, 883–888. [Google Scholar]
- Leone, S.; Chiavaroli, A.; Shohreh, R.; Ferrante, C.; Ricciuti, A.; Manippa, F.; Recinella, L.; Di Nisio, C.; Orlando, G.; Salvatori, R. Increased locomotor and thermogenic activity in mice with targeted ablation of the GHRH gene. Growth Horm. IGF Res. 2015, 25, 80–84. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- di Giacomo, V.; Ferrante, C.; Ronci, M.; Cataldi, A.; Di Valerio, V.; Rapino, M.; Recinella, L.; Chiavaroli, A.; Leone, S.; Vladimir-Knežević, S. Multiple pharmacological and toxicological investigations on Tanacetum parthenium and Salix alba extracts: Focus on potential application as anti-migraine agents. Food Chem. Toxicol. 2019, 133, 110783. [Google Scholar] [CrossRef]
- Distler, A.M.; Kerner, J.; Peterman, S.M.; Hoppel, C.L. A targeted proteomic approach for the analysis of rat liver mitochondrial outer membrane proteins with extensive sequence coverage. Anal. Biochem. 2006, 356, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, C.; Recinella, L.; Ronci, M.; Menghini, L.; Brunetti, L.; Chiavaroli, A.; Leone, S.; Di Iorio, L.; Carradori, S.; Tirillini, B. Multiple pharmacognostic characterization on hemp commercial cultivars: Focus on inflorescence water extract activity. Food Chem. Toxicol. 2019, 125, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, C.; Recinella, L.; Ronci, M.; Orlando, G.; Di Simone, S.; Brunetti, L.; Chiavaroli, A.; Leone, S.; Politi, M.; Tirillini, B. Protective effects induced by alcoholic Phlomis fruticosa and Phlomis herba-venti extracts in isolated rat colon: Focus on antioxidant, anti-inflammatory, and antimicrobial activities in vitro. Phytother. Res. 2019, 33, 2387–2400. [Google Scholar] [CrossRef] [PubMed]
- Charan, J.; Kantharia, N. How to calculate sample size in animal studies? J. Pharmacol. Pharmacother. 2013, 4, 303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menghini, L.; Recinella, L.; Leone, S.; Chiavaroli, A.; Cicala, C.; Brunetti, L.; Vladimir-Knežević, S.; Orlando, G.; Ferrante, C. Devil’s claw (Harpagophytum procumbens) and chronic inflammatory diseases: A concise overview on preclinical and clinical data. Phytother. Res. 2019, 33, 2152–2162. [Google Scholar] [CrossRef] [PubMed]
- Menghini, L.; Ferrante, C.; Leporini, L.; Recinella, L.; Chiavaroli, A.; Leone, S.; Pintore, G.; Vacca, M.; Orlando, G.; Brunetti, L. An hydroalcoholic chamomile extract modulates inflammatory and immune response in HT29 cells and isolated rat colon. Phytother. Res. 2016, 30, 1513–1518. [Google Scholar] [CrossRef]
- Banaganapalli, B.; Mulakayala, C.; Gowsia, D.; Mulakayala, N.; Pulaganti, M.; Shaik, N.A.; Anuradha, C.; Rao, R.M.; Al-Aama, J.Y.; Chitta, S.K. Synthesis and biological activity of new resveratrol derivative and molecular docking: Dynamics studies on NFkB. Appl. Biochem. Biotechnol. 2013, 171, 1639–1657. [Google Scholar] [CrossRef]
- Chiavaroli, A.; Brunetti, L.; Orlando, G.; Recinella, L.; Ferrante, C.; Leone, S.; Di Michele, P.; Di Nisio, C.; Vacca, M. Resveratrol inhibits isoprostane production in young and aged rat brain. J. Biol. Regul. Homeost. Agents 2010, 24, 441. [Google Scholar]
- Shanmuganathan, S.; Angayarkanni, N. Chebulagic acid Chebulinic acid and Gallic acid, the active principles of Triphala, inhibit TNFα induced pro-angiogenic and pro-inflammatory activities in retinal capillary endothelial cells by inhibiting p38, ERK and NFkB phosphorylation. Vasc. Pharmacol. 2018, 108, 23–35. [Google Scholar] [CrossRef]
- Fiebich, B.L.; Muñoz, E.; Rose, T.; Weiss, G.; McGregor, G.P. Molecular targets of the antiinflammatory Harpagophytum procumbens (devil’s claw): Inhibition of TNFα and COX-2 gene expression by preventing activation of AP-1. Phytother. Res. 2012, 26, 806–811. [Google Scholar] [CrossRef]
- Lin, Y.-C.; Huang, D.-Y.; Chu, C.-L.; Lin, W.-W. Anti-inflammatory actions of Syk inhibitors in macrophages involve non-specific inhibition of toll-like receptors-mediated JNK signaling pathway. Mol. Immunol. 2010, 47, 1569–1578. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Ng, S.C. The gut microbiota in the pathogenesis and therapeutics of inflammatory bowel disease. Front. Microbiol. 2018, 9, 2247. [Google Scholar] [CrossRef] [PubMed]
- Nazareth, N.; Magro, F.; Machado, E.; Ribeiro, T.G.; Martinho, A.; Rodrigues, P.; Alves, R.; Macedo, G.N.; Gracio, D.; Coelho, R. Prevalence of Mycobacteriumavium subsp. paratuberculosis and Escherichia coli in blood samples from patients with inflammatory bowel disease. Med. Microbiol. Immunol. 2015, 204, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Soares, S.; Abreu, P.; Jesus, L.; Brito, L.; Bernardo-Filho, M. Protective effect of an aqueous extract of Harpagophytum procumbens upon Escherichia coli strains submitted to the lethal action of stannous chloride. Cell. Mol. Biol. 2007, 53, OL923–OL927. [Google Scholar]
- Weckesser, S.; Engel, K.; Simon-Haarhaus, B.; Wittmer, A.; Pelz, K.; Schempp, C.Á. Screening of plant extracts for antimicrobial activity against bacteria and yeasts with dermatological relevance. Phytomedicine 2007, 14, 508–516. [Google Scholar] [CrossRef]
- Betts, J.W.; Wareham, D.W.; Haswell, S.J.; Kelly, S.M. Antifungal synergy of theaflavin and epicatechin combinations against Candida albicans. J. Microbiol. Biotechnol. 2013, 23, 1322–1326. [Google Scholar] [CrossRef] [Green Version]
- Bottari, N.B.; Lopes, L.Q.S.; Pizzuti, K.; dos Santos Alves, C.F.; Corrêa, M.S.; Bolzan, L.P.; Zago, A.; de Almeida Vaucher, R.; Boligon, A.A.; Giongo, J.L. Antimicrobial activity and phytochemical characterization of Carya illinoensis. Microb. Pathog. 2017, 104, 190–195. [Google Scholar] [CrossRef]
- de Camargo, A.C.; Regitano-d’Arce, M.A.B.; Rasera, G.B.; Canniatti-Brazaca, S.G.; do Prado-Silva, L.; Alvarenga, V.O.; Sant’Ana, A.S.; Shahidi, F. Phenolic acids and flavonoids of peanut by-products: Antioxidant capacity and antimicrobial effects. Food Chem. 2017, 237, 538–544. [Google Scholar] [CrossRef]
- Orlando, G.; Zengin, G.; Ferrante, C.; Ronci, M.; Recinella, L.; Senkardes, I.; Gevrenova, R.; Zheleva-Dimitrova, D.; Chiavaroli, A.; Leone, S. Comprehensive Chemical Profiling and Multidirectional Biological Investigation of Two Wild Anthemis Species (Anthemis tinctoria var. Pallida and A. cretica subsp. tenuiloba): Focus on Neuroprotective Effects. Molecules 2019, 24, 2582. [Google Scholar] [CrossRef] [Green Version]
- Detienne, G.; De Haes, W.; Mergan, L.; Edwards, S.L.; Temmerman, L.; Van Bael, S. Beyond ROS clearance: Peroxiredoxins in stress signaling and aging. Ageing Res. Rev. 2018, 44, 33–48. [Google Scholar] [CrossRef]
- Rashid, K.; Sinha, K.; Sil, P.C. An update on oxidative stress-mediated organ pathophysiology. Food Chem. Toxicol. 2013, 62, 584–600. [Google Scholar] [CrossRef] [PubMed]
- Karachaliou, C.-E.; Kalbacher, H.; Voelter, W.; Tsitsilonis, O.E.; Livaniou, E. In Vitro immunodetection of prothymosin alpha in normal and pathological conditions. Curr. Med. Chem. 2019. [Google Scholar] [CrossRef]
- Rocha-Perugini, V.; Gordon-Alonso, M.; Sánchez-Madrid, F. Role of Drebrin at the immunological synapse. In Drebrin; Springer: Berlin/Heidelberg, Germany, 2017; pp. 271–280. [Google Scholar]
- Wang, G.-H.; Wang, J.-J.; Yue, B.; Du, X.; Du, H.-H.; Zhang, M.; Hu, Y.-H. High mobility group box 2 of black rockfish Sebastes schlegelii: Gene cloning, immunoregulatory properties and antibacterial effect. Fish Shellfish Immunol. 2019, 84, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Chen, J.; Ding, Q.; Yang, S.; Wang, J.; Yu, H.; Lin, J. Function of the macrophage-capping protein in colorectal carcinoma. Oncol. Lett. 2017, 14, 5549–5555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimm-Günter, E.-M.S.; Revenu, C.; Ramos, S.; Hurbain, I.; Smyth, N.; Ferrary, E.; Louvard, D.; Robine, S.; Rivero, F. Plastin 1 binds to keratin and is required for terminal web assembly in the intestinal epithelium. Mol. Biol. Cell 2009, 20, 2549–2562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaguchi, K.; Yoshida, S.; Hatano, R.; Asano, S. Pathophysiological roles of ezrin/radixin/moesin proteins. Biol. Pharm. Bull. 2017, 40, 381–390. [Google Scholar] [CrossRef] [Green Version]
- Petchampai, N.; Sunyakumthorn, P.; Guillotte, M.L.; Verhoeve, V.I.; Banajee, K.H.; Kearney, M.T.; Macaluso, K.R. Novel identification of Dermacentor variabilis Arp2/3 complex and its role in rickettsial infection of the arthropod vector. PLoS ONE 2014, 9, e93768. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Table 1 | Total Phenols | Total Flavonoids | ||
|---|---|---|---|---|
| mg/g Extract | SD | mg/g Extract | SD | |
| H. procumbens | 65.7 | 6.9 | 5.4 | 1.9 |
| Phenolic Content of H. procumbens Extract | mg/g Extract | SD |
|---|---|---|
| Gallic acid | 9.74 | 0.88 |
| Catechin | 2.90 | 0.35 |
| Epicatechin | 3.03 | 0.18 |
| Resveratrol | 3.33 | 0.33 |
| Antitadical Activity | DPPH | β-Carotene/Linoleic Acid | ||
|---|---|---|---|---|
| IC50 µg/mL | SD | IC50 µg/mL | SD | |
| BHT | 2.5 | 0.32 | ||
| Trolox | 4.07 | 0.44 | 4.03 | 0.57 |
| H. procumbens | 121.01 | 16.6 | 16.8 | 2.05 |
| Table 4 | MIC (µg/mL)* | |
|---|---|---|
| H. procumbens | Ciprofloxacin | |
| E. coli (clinical isolate) | 11.80 (9.37–18.75) | <0.12 |
| P. aeruginosa (clinical isolate) | 188.98 (150–300) | <0.12 |
| S. aureus (ATCC 6538) | >300 | 0.98 |
| Table 5 | MIC (µg/mL)* | |
|---|---|---|
| H. procumbens | Fluconazole | |
| C. albicans (YEPGA 6183) | 11.80 (9.37–18.75) | 2 |
| C. tropicalis (YEPGA 6184) | 5.89 (4.68–9.37) | 4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Recinella, L.; Chiavaroli, A.; Ronci, M.; Menghini, L.; Brunetti, L.; Leone, S.; Tirillini, B.; Angelini, P.; Covino, S.; Venanzoni, R.; et al. Multidirectional Pharma-Toxicological Study on Harpagophytum procumbens DC. ex Meisn.: An IBD-Focused Investigation. Antioxidants 2020, 9, 168. https://doi.org/10.3390/antiox9020168
Recinella L, Chiavaroli A, Ronci M, Menghini L, Brunetti L, Leone S, Tirillini B, Angelini P, Covino S, Venanzoni R, et al. Multidirectional Pharma-Toxicological Study on Harpagophytum procumbens DC. ex Meisn.: An IBD-Focused Investigation. Antioxidants. 2020; 9(2):168. https://doi.org/10.3390/antiox9020168
Chicago/Turabian StyleRecinella, Lucia, Annalisa Chiavaroli, Maurizio Ronci, Luigi Menghini, Luigi Brunetti, Sheila Leone, Bruno Tirillini, Paola Angelini, Stefano Covino, Roberto Venanzoni, and et al. 2020. "Multidirectional Pharma-Toxicological Study on Harpagophytum procumbens DC. ex Meisn.: An IBD-Focused Investigation" Antioxidants 9, no. 2: 168. https://doi.org/10.3390/antiox9020168
APA StyleRecinella, L., Chiavaroli, A., Ronci, M., Menghini, L., Brunetti, L., Leone, S., Tirillini, B., Angelini, P., Covino, S., Venanzoni, R., Zengin, G., Simone, S. D., Ciferri, M. C., Giacomo, V. d., Cataldi, A., Rapino, M., Valerio, V. D., Orlando, G., & Ferrante, C. (2020). Multidirectional Pharma-Toxicological Study on Harpagophytum procumbens DC. ex Meisn.: An IBD-Focused Investigation. Antioxidants, 9(2), 168. https://doi.org/10.3390/antiox9020168