d-Galactose Decreases Anion Exchange Capability through Band 3 Protein in Human Erythrocytes

 ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Solutions and Chemicals
2.2. Erythrocytes Preparation
2.3. SO42− Uptake Measurement
2.3.1. Control Condition
2.3.2. d-Gal-Treated Erythrocytes
2.4. Oxidative Condition Assessment
2.4.1. Thiobarbituric Acid Reactive Species Determination
2.4.2. Membrane Sulfhydryl Groups Levels Determination
2.4.3. Methemoglobin Levels Determination
2.5. Erythrocytes Membranes Preparation
2.6. SDS-PAGE Preparation and Western Blot Analysis
2.7. Glycated Hemoglobin Measurement (%A1c)
2.8. Experimental Data and Statistics
3. Results
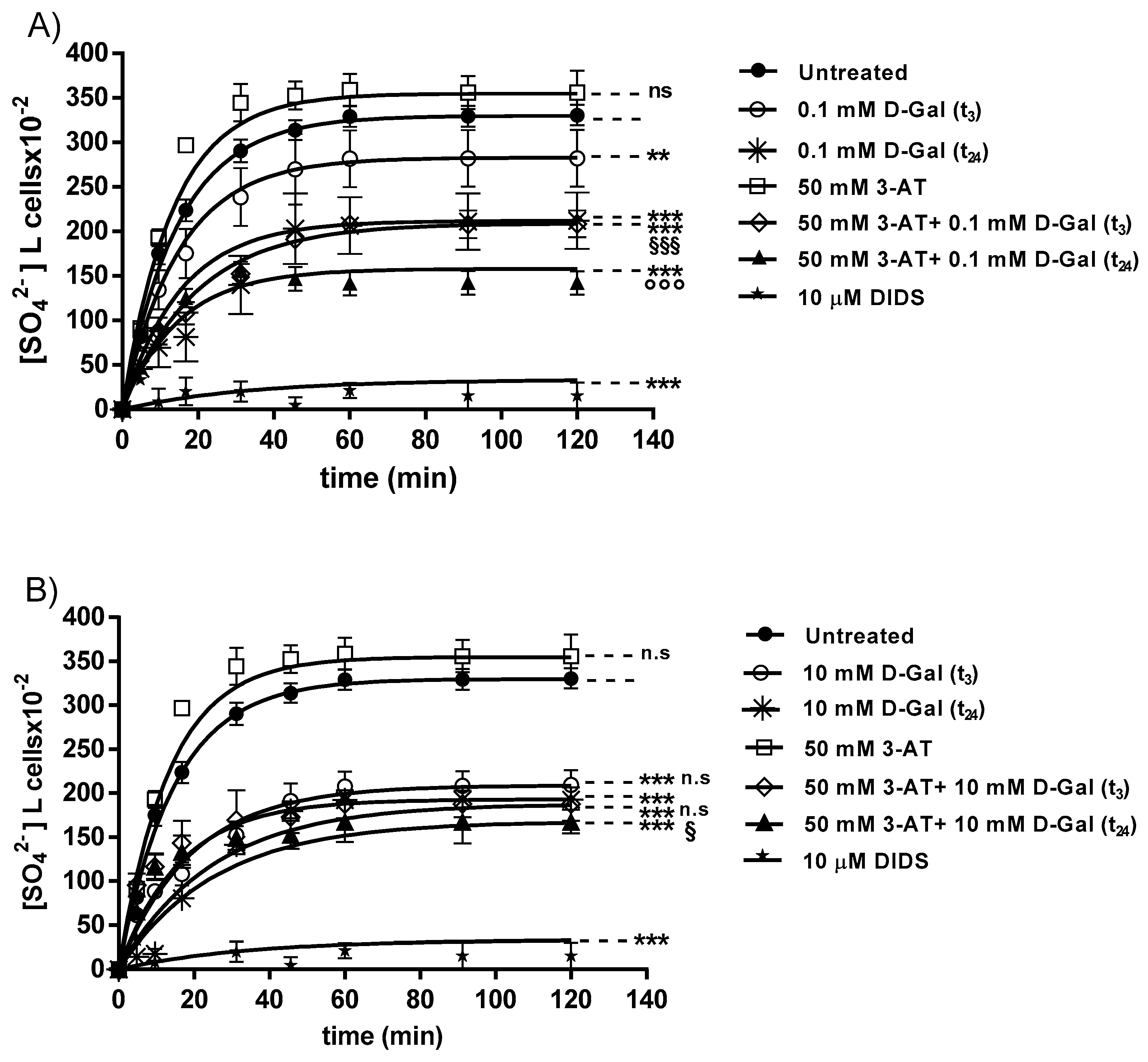
3.1. SO42− Uptake Measurement
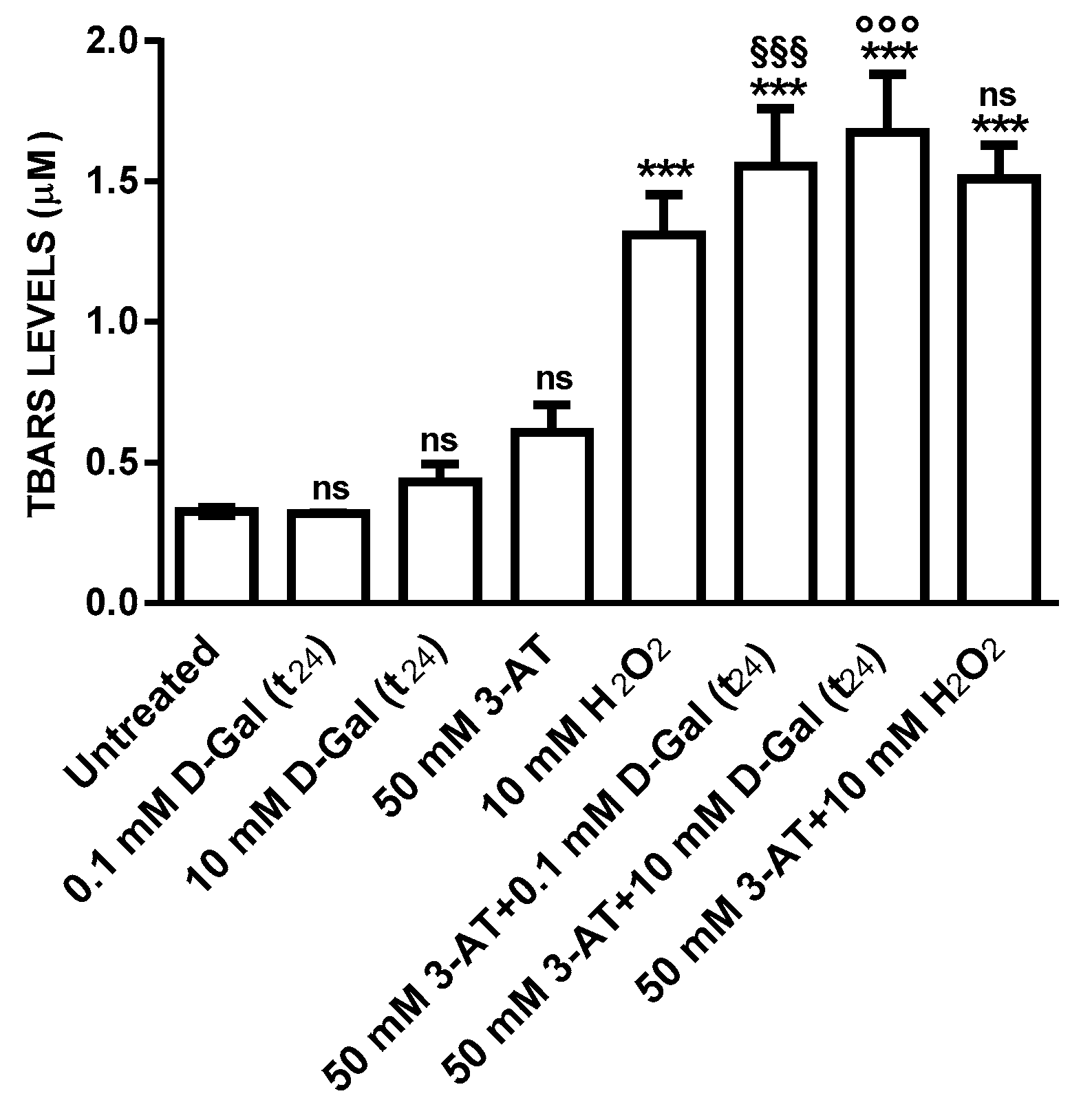
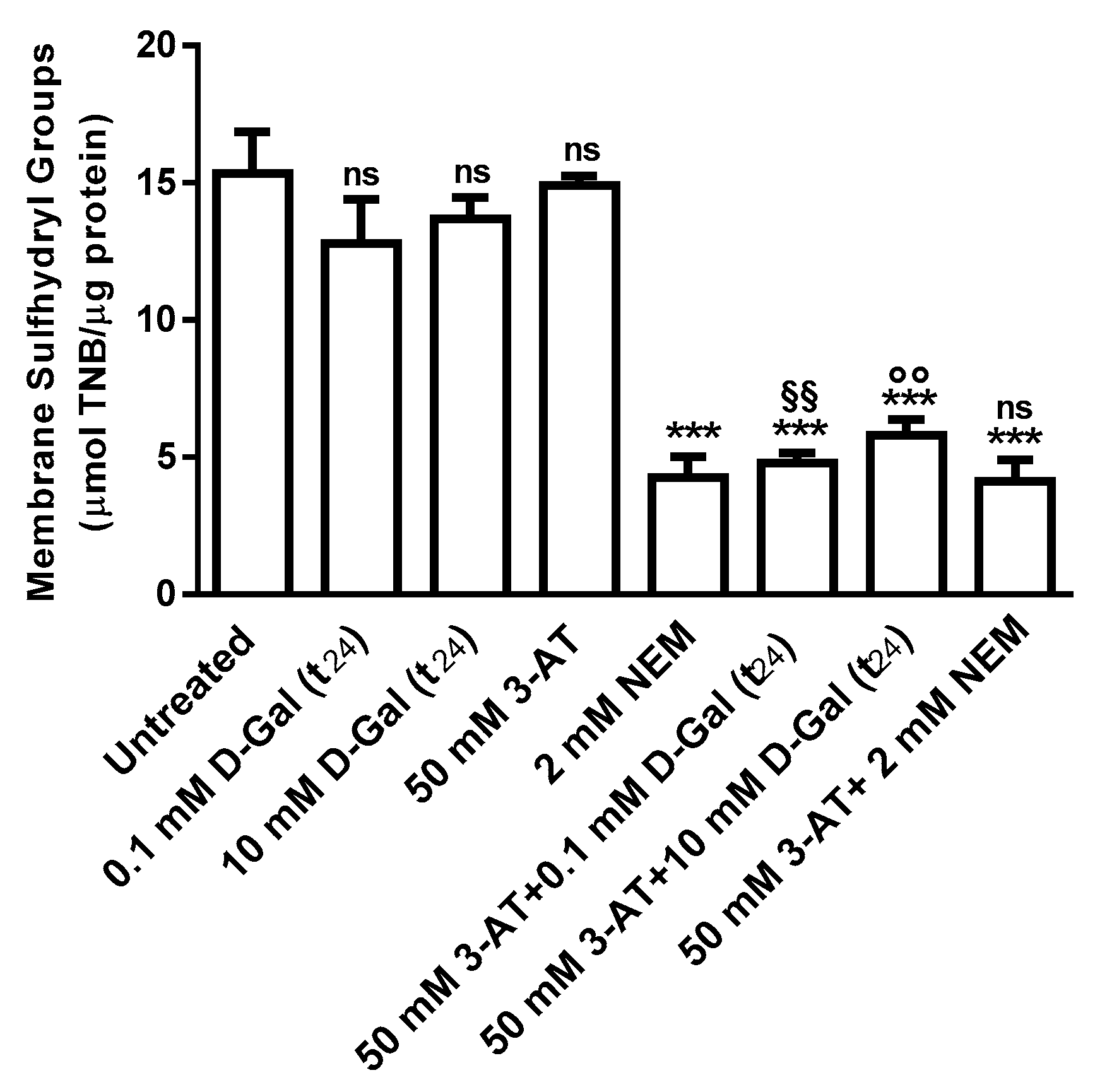
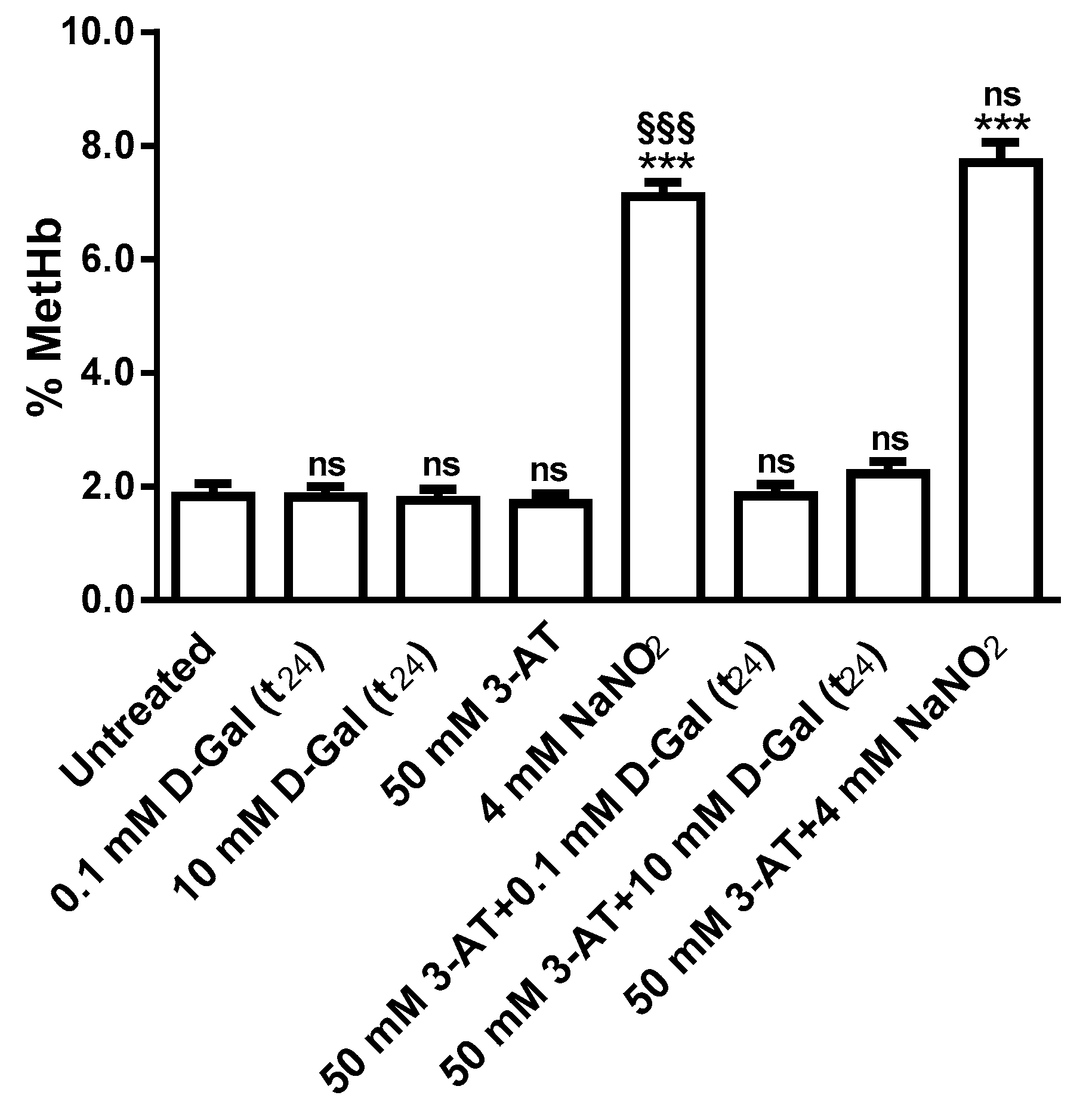
3.2. Oxidative Conditions Assessment
3.2.1. TBARS Levels
3.2.2. Membrane Sulfhydryl Groups Content Measurement
3.2.3. Methemoglobin Measurement
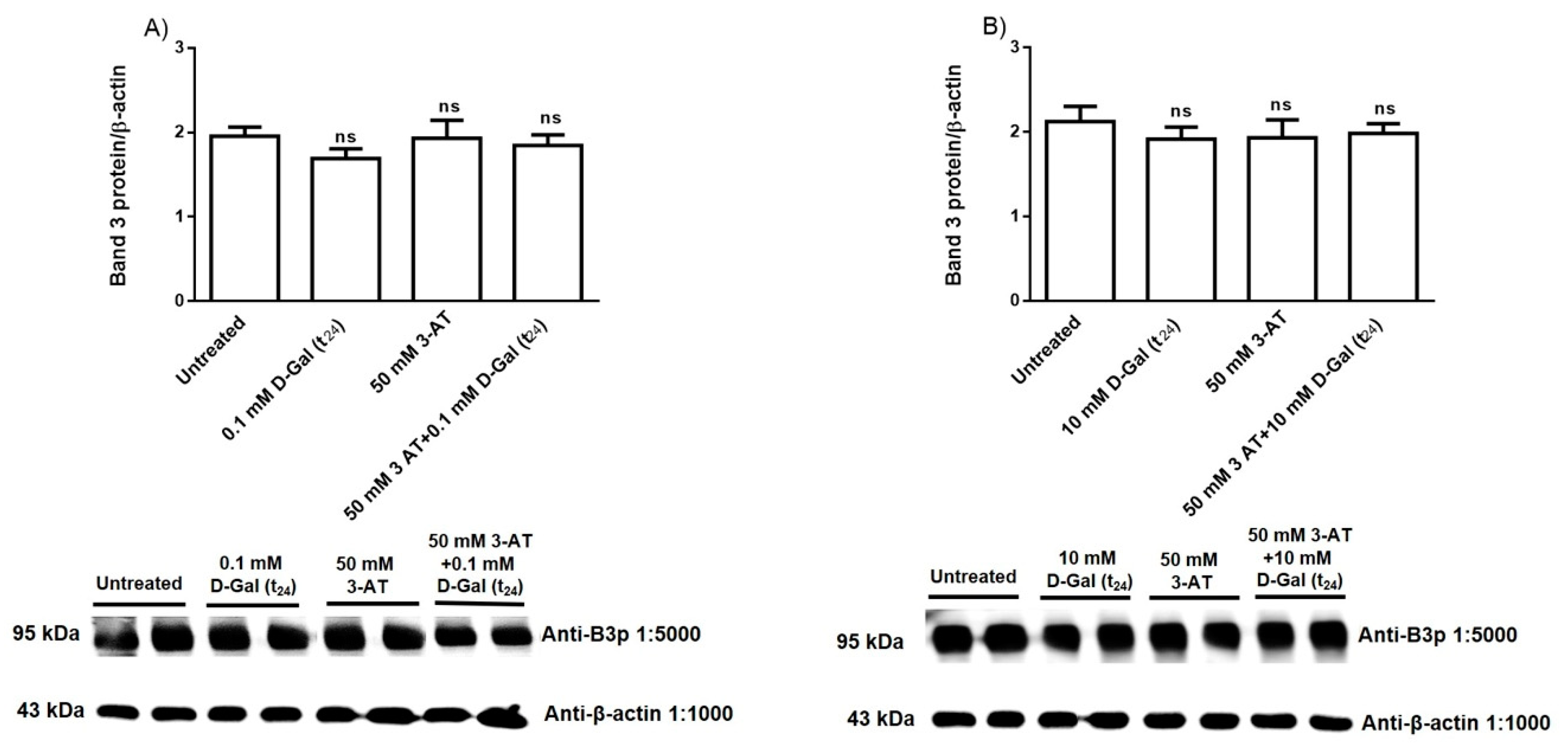
3.3. Band 3 Protein Expression Levels Determination
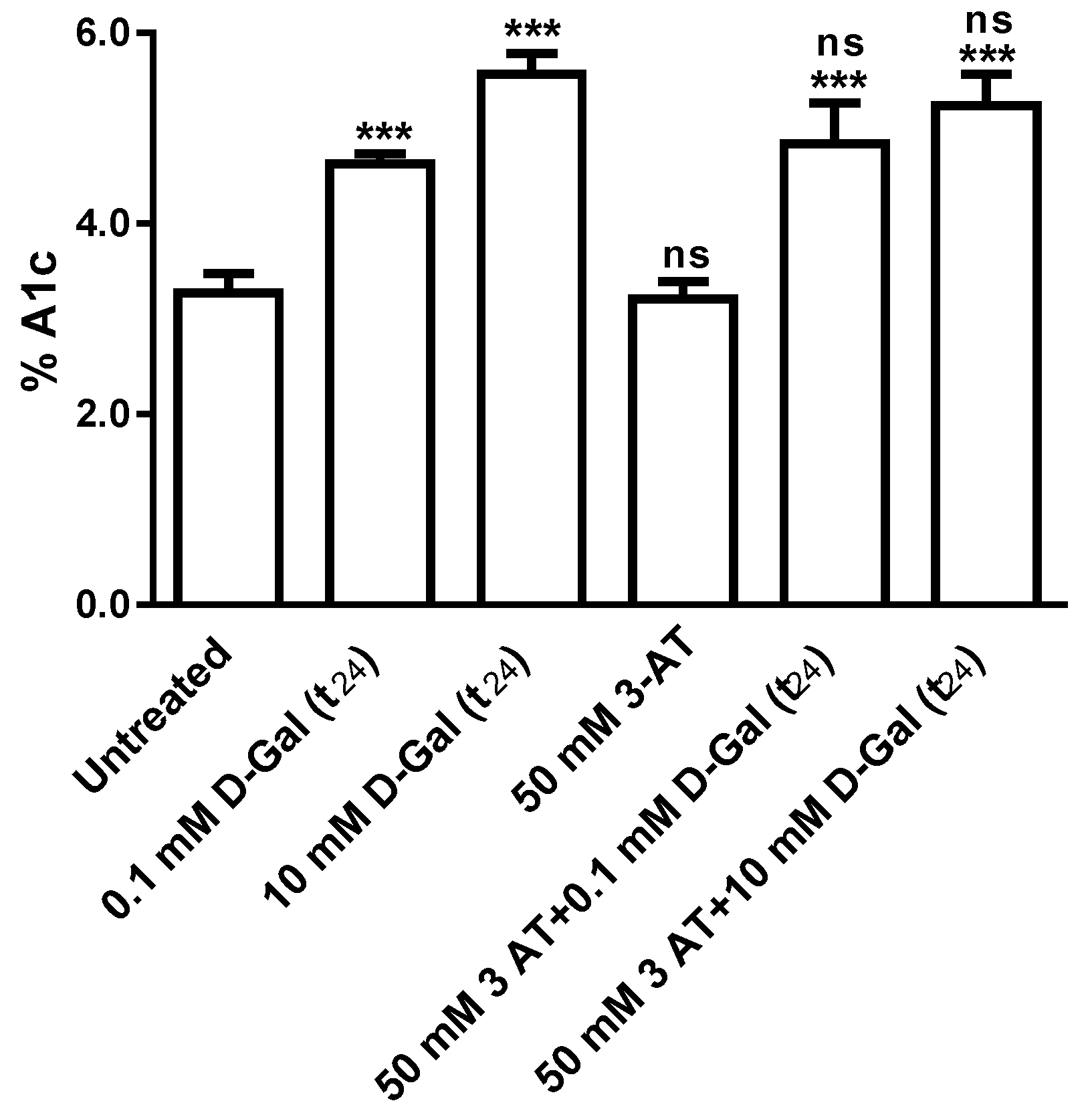
3.4. Glycated Hemoglobin Measurement
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Coelho, A.I.; Berry, G.T.; Rubio-Gozalbo, M.E. Galactose metabolism and health. Curr. Opin. Clin. Nutr. Metab Care 2015, 18, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.M. Classical galactosaemia revisited. J. Inherit Metab. Dis. 2006, 29, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Bossolan, G.; Trindade, C.E.; Barreiros, R.C. Blood galactose and glucose levels in mothers, cord blood, and 48-h-old breast-fed full-term infants. Neonatology 2007, 91, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Schadewaldt, P.; Hammen, H.W.; Loganathan, K.; Bodner-Leidecker, A.; Wendel, U. Analysis of concentration and (13)C enrichment of D-galactose in human plasma. Clin. Chem. 2000, 46, 612–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gitzelmann, R. Galactose-1-phosphate in the pathophysiology of galactosemia. Eur. J. Pediatr. 1995, 154, S45–49. [Google Scholar] [CrossRef] [PubMed]
- Delwing-de Lima, D.; Hennrich, S.B.; Delwing-Dal Magro, D.; Aurelio, J.G.; Serpa, A.P.; Augusto, T.W.; Pereira, N.R. The effect of d-galactose induced oxidative stress on in vitro redox homeostasis in rat plasma and erythrocytes. Biomed. Pharmacother. 2017, 86, 686–693. [Google Scholar] [CrossRef]
- Bo-Htay, C.; Palee, S.; Apaijai, N.; Chattipakorn, S.C.; Chattipakorn, N. Effects of d-galactose-induced ageing on the heart and its potential interventions. J. Cell Mol. Med. 2018, 22, 1392–1410. [Google Scholar] [CrossRef] [Green Version]
- Luevano-Contreras, C.; Chapman-Novakofski, K. Dietary advanced glycation end products and aging. Nutrients 2010, 2, 1247–1265. [Google Scholar] [CrossRef] [Green Version]
- Ansari, N.A.; Dash, D. Amadori glycated proteins: Role in production of autoantibodies in diabetes mellitus and effect of inhibitors on non-enzymatic glycation. Aging Dis. 2013, 4, 50–56. [Google Scholar]
- Cebe, T.; Yanar, K.; Atukeren, P.; Ozan, T.; Kuruc, A.I.; Kunbaz, A.; Sitar, M.E.; Mengi, M.; Aydin, M.S.; Esrefoglu, M.; et al. A comprehensive study of myocardial redox homeostasis in naturally and mimetically aged rats. Age (Dordr.) 2014, 36, 9728. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, Y.; Yuan, Y.; Sun, Y.; Qin, Y.; Deng, Z.; Li, H. Protective Effects of Selenium, Vitamin E, and Purple Carrot Anthocyanins on D-Galactose-Induced Oxidative Damage in Blood, Liver, Heart and Kidney Rats. Biol. Trace Elem. Res. 2016, 173, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Doroszko, A.; Radziwon-Balicka, A.; Skomro, R. Novel Approaches for Diagnosing and Management of Cardiovascular Disorders Mediated by Oxidative Stress. Oxid. Med. Cell Longev. 2020, 2020, 7096727. [Google Scholar] [CrossRef] [PubMed]
- Brunelli, E.; La Russa, D.; Pellegrino, D. Impaired Oxidative Status Is Strongly Associated with Cardiovascular Risk Factors. Oxid. Med. Cell Longev. 2017, 2017, 6480145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ton, A.M.M.; Campagnaro, B.P.; Alves, G.A.; Aires, R.; Coco, L.Z.; Arpini, C.M.; Guerra, E.O.T.; Campos-Toimil, M.; Meyrelles, S.S.; Pereira, T.M.C.; et al. Oxidative Stress and Dementia in Alzheimer’s Patients: Effects of Synbiotic Supplementation. Oxid. Med. Cell Longev. 2020, 2020, 2638703. [Google Scholar] [CrossRef]
- Janion, K.; Szczepanska, E.; Nowakowska-Zajdel, E.; Walkiewicz, K.; Strzelczyk, J. Lipid peroxidation and total oxidant/antioxidant status in colorectal cancer patients. J. Biol. Regul. Homeost. Agents 2020, 34, 239–244. [Google Scholar] [CrossRef]
- Zhang, P.; Li, T.; Wu, X.; Nice, E.C.; Huang, C.; Zhang, Y. Oxidative stress and diabetes: Antioxidative strategies. Front. Med. 2002. [Google Scholar] [CrossRef]
- Iglesias-Pedraz, J.M.; Comai, L. Measurements of Hydrogen Peroxide and Oxidative DNA Damage in a Cell Model of Premature Aging. Methods Mol. Biol. 2020, 2144, 245–257. [Google Scholar] [CrossRef]
- Hsia, C.H.; Wang, C.H.; Kuo, Y.W.; Ho, Y.J.; Chen, H.L. Fructo-oligosaccharide systemically diminished D-galactose-induced oxidative molecule damages in BALB/cJ. mice. Br. J. Nutr. 2012, 107, 1787–1792. [Google Scholar] [CrossRef] [Green Version]
- Mladenov, M.; Gokik, M.; Hadzi-Petrushev, N.; Gjorgoski, I.; Jankulovski, N. The relationship between antioxidant enzymes and lipid peroxidation in senescent rat erythrocytes. Physiol. Res. 2015, 64, 891–896. [Google Scholar] [CrossRef]
- Song, X.; Bao, M.; Li, D.; Li, Y.M. Advanced glycation in d-galactose induced mouse aging model. Mech. Ageing Dev. 1999, 108, 239–251. [Google Scholar] [CrossRef]
- Haider, S.; Liaquat, L.; Shahzad, S.; Sadir, S.; Madiha, S.; Batool, Z.; Tabassum, S.; Saleem, S.; Naqvi, F.; Perveen, T. A high dose of short term exogenous D-galactose administration in young male rats produces symptoms simulating the natural aging process. Life Sci. 2015, 124, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Inès Birlouez-Aragon, P.S.-M. Marek Morawiec and Majid Shafiezadeh Evidence for a relationship between protein glycation and red blood cell membrane fluidity. Biochem. Bioph. Res. Commun. 1990, 170, 1107–1113. [Google Scholar] [CrossRef]
- Ando, K.; Beppu, M.; Kikugawa, K.; Nagai, R.; Horiuchi, S. Membrane proteins of human erythrocytes are modified by advanced glycation end products during aging in the circulation. Biochem. Biophys. Res. Commun. 1999, 258, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Sompong, W.; Cheng, H.; Adisakwattana, S. Protective Effects of Ferulic Acid on High Glucose-Induced Protein Glycation, Lipid Peroxidation, and Membrane Ion Pump Activity in Human Erythrocytes. PLoS ONE 2015, 10, e0129495. [Google Scholar] [CrossRef] [Green Version]
- Viskupicova, J.; Blaskovic, D.; Galiniak, S.; Soszynski, M.; Bartosz, G.; Horakova, L.; Sadowska-Bartosz, I. Effect of high glucose concentrations on human erythrocytes in vitro. Redox Biol. 2015, 5, 381–387. [Google Scholar] [CrossRef] [Green Version]
- Remigante, A.; Morabito, R.; Marino, A. Natural Antioxidants Beneficial Effects on Anion Exchange through Band 3 Protein in Human Erythrocytes. Antioxidants (Basel) 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Morabito, R.; Romano, O.; La Spada, G.; Marino, A. H2O2-Induced Oxidative Stress Affects SO4 = Transport in Human Erythrocytes. PLoS ONE 2016, 11, e0146485. [Google Scholar] [CrossRef] [Green Version]
- van Zwieten, R.; Verhoeven, A.J.; Roos, D. Inborn defects in the antioxidant systems of human red blood cells. Free Radic. Biol. Med. 2014, 67, 377–386. [Google Scholar] [CrossRef]
- Reithmeier, R.A.; Casey, J.R.; Kalli, A.C.; Sansom, M.S.; Alguel, Y.; Iwata, S. Band 3, the human red cell chloride/bicarbonate anion exchanger (AE1, SLC4A1), in a structural context. Biochim. Biophys. Acta 2016, 1858, 1507–1532. [Google Scholar] [CrossRef]
- Hatae, H.; Inaka, K.; Okamura, R.; Furubayashi, N.; Kamo, M.; Kobayashi, T.; Abe, Y.; Iwata, S.; Hamasaki, N. Crystallization of Human Erythrocyte Band 3, the anion exchanger, at the International Space Station “KIBO”. Anal. Biochem. 2018, 559, 91–93. [Google Scholar] [CrossRef]
- Romano, L.; Peritore, D.; Simone, E.; Sidoti, A.; Trischitta, F.; Romano, P. Chloride-sulphate exchange chemically measured in human erythrocyte ghosts. Cell Mol. Biol. (Noisy-le-grand) 1998, 44, 351–355. [Google Scholar]
- Jennings, M.L. Proton fluxes associated with erythrocyte membrane anion exchange. J. Membr. Biol. 1976, 28, 187–205. [Google Scholar] [CrossRef] [PubMed]
- Morabito, R.; Remigante, A.; Spinelli, S.; Vitale, G.; Trichilo, V.; Loddo, S.; Marino, A. High Glucose Concentrations Affect Band 3 Protein in Human Erythrocytes. Antioxidants (Basel) 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Morabito, R.; Remigante, A.; Di Pietro, M.L.; Giannetto, A.; La Spada, G.; Marino, A. SO4(=) uptake and catalase role in preconditioning after H2O2-induced oxidative stress in human erythrocytes. Pflugers Arch. 2017, 469, 235–250. [Google Scholar] [CrossRef]
- Morabito, R.; Remigante, A.; Cordaro, M.; Trichilo, V.; Loddo, S.; Dossena, S.; Marino, A. Impact of acute inflammation on Band 3 protein anion exchange capability in human erythrocytes. Arch. Physiol. Biochem. 2020. [Google Scholar] [CrossRef]
- Pantaleo, A.; Ferru, E.; Pau, M.C.; Khadjavi, A.; Mandili, G.; Matte, A.; Spano, A.; De Franceschi, L.; Pippia, P.; Turrini, F. Band 3 Erythrocyte Membrane Protein Acts as Redox Stress Sensor Leading to Its Phosphorylation by p (72) Syk. Oxid. Med. Cell Longev. 2016, 2016, 6051093. [Google Scholar] [CrossRef] [Green Version]
- Margoliash, E.; Novogrodsky, A.; Schejter, A. Irreversible reaction of 3-amino-1:2:4-triazole and related inhibitors with the protein of catalase. Biochem. J. 1960, 74, 339–348. [Google Scholar] [CrossRef] [Green Version]
- Romano, L.; Passow, H. Characterization of anion transport system in trout red blood cell. Am. J. Physiol. 1984, 246, C330–338. [Google Scholar] [CrossRef]
- Jessen, F.; Sjoholm, C.; Hoffmann, E.K. Identification of the anion exchange protein of Ehrlich cells: A kinetic analysis of the inhibitory effects of 4,4′-diisothiocyano-2,2′-stilbene-disulfonic acid (DIDS) and labeling of membrane proteins with 3H-DIDS. J. Membr. Biol. 1986, 92, 195–205. [Google Scholar] [CrossRef]
- Mendanha, S.A.; Anjos, J.L.; Silva, A.H.; Alonso, A. Electron paramagnetic resonance study of lipid and protein membrane components of erythrocytes oxidized with hydrogen peroxide. Braz. J. Med. Biol. Res. 2012, 45, 473–481. [Google Scholar] [CrossRef] [Green Version]
- Sokolowska, M.; Oleszek, A.; Wlodek, L. Protective effect of alpha-keto acids on the oxidative hemolysis. Pol. J. Pharmacol. 1999, 51, 429–434. [Google Scholar]
- Aksenov, M.Y.; Markesbery, W.R. Changes in thiol content and expression of glutathione redox system genes in the hippocampus and cerebellum in Alzheimer’s disease. Neurosci. Lett. 2001, 302, 141–145. [Google Scholar] [CrossRef]
- Morabito, R.; Falliti, G.; Geraci, A.; Spada, G.L.; Marino, A. Curcumin Protects -SH Groups and Sulphate Transport after Oxidative Damage in Human Erythrocytes. Cell Physiol. Biochem. 2015, 36, 345–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naoum, P.C.R.; Radispiel, J.; Magaly da Silva, M. Spectrometric measurement of methemoglobin without interference of chemical or enzymatic reagents. Rev. Bras. Hematol. Hemoter. 2004, 26, 19–22. [Google Scholar]
- Morabito, R.; Remigante, A.; Cavallaro, M.; Taormina, A.; La Spada, G.; Marino, A. Anion exchange through band 3 protein in canine leishmaniasis at different stages of disease. Pflugers Arch. 2017, 469, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Zavodnik, I.B.; Lapshina, E.A.; Rekawiecka, K.; Zavodnik, L.B.; Bartosz, G.; Bryszewska, M. Membrane effects of nitrite-induced oxidation of human red blood cells. Biochim. Biophys. Acta 1999, 1421, 306–316. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Yeung, Y.G.; Stanley, E.R. A solution for stripping antibodies from polyvinylidene fluoride immunoblots for multiple reprobing. Anal. Biochem. 2009, 389, 89–91. [Google Scholar] [CrossRef] [Green Version]
- Cebe, T.; Atukeren, P.; Yanar, K.; Kuruc, A.I.; Ozan, T.; Kunbaz, A.; Sitar, M.E.; Mirmaroufizibandeh, R.; Aydin, S.; Cakatay, U. Oxidation scrutiny in persuaded aging and chronological aging at systemic redox homeostasis level. Exp. Gerontol. 2014, 57, 132–140. [Google Scholar] [CrossRef]
- Kayali, R.; Cakatay, U.; Tekeli, F. Male rats exhibit higher oxidative protein damage than females of the same chronological age. Mech. Ageing Dev. 2007, 128, 365–369. [Google Scholar] [CrossRef]
- Cakatay, U.; Kayali, R.; Uzun, H. Relation of plasma protein oxidation parameters and paraoxonase activity in the ageing population. Clin. Exp. Med. 2008, 8, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Yelinova, V.; Glazachev, Y.; Khramtsov, V.; Kudryashova, L.; Rykova, V.; Salganik, R. Studies of human and rat blood under oxidative stress: Changes in plasma thiol level, antioxidant enzyme activity, protein carbonyl content, and fluidity of erythrocyte membrane. Biochem. Biophys. Res. Commun. 1996, 221, 300–303. [Google Scholar] [CrossRef] [PubMed]
- Welbourn, E.M.; Wilson, M.T.; Yusof, A.; Metodiev, M.V.; Cooper, C.E. The mechanism of formation, structure and physiological relevance of covalent hemoglobin attachment to the erythrocyte membrane. Free Radic. Biol. Med. 2017, 103, 95–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luthra, M.G.; Kim, H.D. Effects of calcium and soluble cytoplasmic activator protein (calmodulin) on various states of (Ca2+ + Mg2+)-ATPase activity in isolated membranes of human red cells. Biochim. Biophys. Acta 1980, 600, 467–479. [Google Scholar] [CrossRef]
- Rifkind, J.M.; Nagababu, E. Hemoglobin redox reactions and red blood cell aging. Antioxid. Redox Signal 2013, 18, 2274–2283. [Google Scholar] [CrossRef] [Green Version]
- Morabito, R.; Remigante, A.; Bagnato, G.; Neal, R.W.; Sciortino, D.; D’Angelo, T.; Iannelli, F.; Iannelli, F.; Cordova, F.; Cirillo, M.; et al. Band 3 Protein Function and Oxidative Stress in Erythrocytes from Systemic Sclerosis Patients with Interstitial Lung Disease. Eur. J. Clin. Biomed. Sci. 2017, 3, 80–84. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.C.; Liu, J.H.; Wu, R.Y. Establishment of the mimetic aging effect in mice caused by d-galactose. Biogerontology 2003, 4, 15–18. [Google Scholar] [CrossRef]
- Roy, S.S.; Sen, G.; Biswas, T. Role of sulfhydryl groups in band 3 in the inhibition of phosphate transport across erythrocyte membrane in visceral leishmaniasis. Arch. Biochem. Biophys. 2005, 436, 121–127. [Google Scholar] [CrossRef]
- Morabito, R.; Remigante, A.; Marino, A. Protective Role of Magnesium against Oxidative Stress on SO4(=) Uptake through Band 3 Protein in Human Erythrocytes. Cell Physiol. Biochem. 2019, 52, 1292–1308. [Google Scholar] [CrossRef]
- Morabito, R.; Remigante, A.; Marino, A. Melatonin Protects Band 3 Protein in Human Erythrocytes against H2O2-Induced Oxidative Stress. Molecules 2019, 24. [Google Scholar] [CrossRef] [Green Version]
- Umbayev, B.; Askarova, S.; Almabayeva, A.; Saliev, T.; Masoud, A.R.; Bulanin, D. Galactose-Induced Skin Aging: The Role of Oxidative Stress. Oxid. Med. Cell Longev. 2020, 2020, 7145656. [Google Scholar] [CrossRef] [PubMed]
- Walder, J.A.; Chatterjee, R.; Steck, T.L.; Low, P.S.; Musso, G.F.; Kaiser, E.T.; Rogers, P.H.; Arnone, A. The interaction of hemoglobin with the cytoplasmic domain of band 3 of the human erythrocyte membrane. J. Biol. Chem. 1984, 259, 10238–10246. [Google Scholar] [PubMed]
- Kenawy, S.; Hegazy, R.; Hassan, A.; El-Shenawy, S.; Gomaa, N.; Zaki, H.; Attia, A. Involvement of insulin resistance in D-galactose-induced age-related dementia in rats: Protective role of metformin and saxagliptin. PLoS ONE 2017, 12, e0183565. [Google Scholar] [CrossRef] [PubMed]
- Faust, R.G. Monosaccharide penetration into human red blood cells by an altered diffusion mechanism. J. Cell Comp. Physiol. 1960, 56, 103–121. [Google Scholar] [CrossRef] [PubMed]
- Ledesma-Osuna, A.I.; Ramos-Clamont, G.; Vazquez-Moreno, L. Characterization of bovine serum albumin glycated with glucose, galactose and lactose. Acta Biochim. Pol. 2008, 55, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Sathiyapriya, V.; Bobby, Z.; Vinod Kumar, S.; Selvaraj, N.; Parthibane, V.; Gupta, S. Evidence for the role of lipid peroxides on glycation of hemoglobin and plasma proteins in non-diabetic asthma patients. Clin. Chim. Acta 2006, 366, 299–303. [Google Scholar] [CrossRef]
- Bartosz, G.; Tannert, C.; Fried, R.; Leyko, W. Superoxide dismutase activity decreases during erythrocyte aging. Experientia 1978, 34, 1464. [Google Scholar] [CrossRef]
- Arai, K.; Maguchi, S.; Fujii, S.; Ishibashi, H.; Oikawa, K.; Taniguchi, N. Glycation and inactivation of human Cu-Zn-superoxide dismutase. Identification of the in vitro glycated sites. J. Biol. Chem. 1987, 262, 16969–16972. [Google Scholar]
- Jafarnejad, A.; Bathaie, S.Z.; Nakhjavani, M.; Hassan, M.Z. Investigation of the mechanisms involved in the high-dose and long-term acetyl salicylic acid therapy of type I diabetic rats. J. Pharmacol. Exp. Ther. 2008, 324, 850–857. [Google Scholar] [CrossRef] [Green Version]
- Rao, G.N.; Lardis, M.P.; Cotlier, E. Acetylation of lens crystallins: A possible mechanism by which aspirin could prevent cataract formation. Biochem. Biophys. Res. Commun. 1985, 128, 1125–1132. [Google Scholar] [CrossRef]
- Jaeschke, H. Glutathione disulfide formation and oxidant stress during acetaminophen-induced hepatotoxicity in mice in vivo: The protective effect of allopurinol. J. Pharmacol. Exp. Ther. 1990, 255, 935–941. [Google Scholar] [PubMed]
- Niki, E.; Traber, M.G. A history of vitamin E. Ann. Nutr. Metab. 2012, 61, 207–212. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rate Constant (min−1) | Time (min) | n | SO4 2− Amount Trapped after a 45 min Incubation in SO4 2− Medium [SO4 2−] L cells x10−2 | |
|---|---|---|---|---|
| untreated (control) | 0.067 ± 0.001 | 14.92 | 8 | 313.81 ± 11.09 |
| 0.1 mM d-Gal (t3) | 0.066 ± 0.001 ** | 15.15 | 6 | 269.76 ± 39.74 ** |
| 0.1 mM d-Gal (t24) | 0.065 ± 0.001 *** | 15.38 | 6 | 203.16 ± 39.74 *** |
| 10 mM d-Gal (t3) | 0.060 ± 0.001 *** | 16.66 | 6 | 191.26 ± 19.90 *** |
| 10 mM d-Gal (t24) 50 mM 3-AT | 0.051 ± 0.001 ***,ns 0.068 ± 0.001 ns | 19.60 14.70 | 6 6 | 179.97 ± 7.23 *** 343.43 ± 12.23 ns |
| 50 mM 3-AT + 0.1 mM d-Gal (t3) | 0.065 ± 0.001 ***,§§§ | 15.38 | 6 | 146.85 ± 18.46 §§§ |
| 50 mM 3-AT + 0.1 mM d-Gal (t24) | 0.066 ± 0.001 ***,°°° | 15.15 | 6 | 191.26 ± 15.21 °°° |
| 50 mM 3-AT + 10 mM d-Gal (t3) | 0.040 ± 0.001 ***, ns | 25.00 | 6 | 172.85 ± 18 ns |
| 50 mM 3-AT + 10 mM d-Gal (t24) | 0.038 ± 0.001 ***,§ | 26.31 | 6 | 152.85 ± 15.87 § |
| 10 µM DIDS | 0.018 ± 0.001 *** | 55.5 | 10 | 4.75 ± 8.50 *** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Remigante, A.; Morabito, R.; Spinelli, S.; Trichilo, V.; Loddo, S.; Sarikas, A.; Dossena, S.; Marino, A. d-Galactose Decreases Anion Exchange Capability through Band 3 Protein in Human Erythrocytes. Antioxidants 2020, 9, 689. https://doi.org/10.3390/antiox9080689
Remigante A, Morabito R, Spinelli S, Trichilo V, Loddo S, Sarikas A, Dossena S, Marino A. d-Galactose Decreases Anion Exchange Capability through Band 3 Protein in Human Erythrocytes. Antioxidants. 2020; 9(8):689. https://doi.org/10.3390/antiox9080689
Chicago/Turabian StyleRemigante, Alessia, Rossana Morabito, Sara Spinelli, Vincenzo Trichilo, Saverio Loddo, Antonio Sarikas, Silvia Dossena, and Angela Marino. 2020. "d-Galactose Decreases Anion Exchange Capability through Band 3 Protein in Human Erythrocytes" Antioxidants 9, no. 8: 689. https://doi.org/10.3390/antiox9080689
APA StyleRemigante, A., Morabito, R., Spinelli, S., Trichilo, V., Loddo, S., Sarikas, A., Dossena, S., & Marino, A. (2020). d-Galactose Decreases Anion Exchange Capability through Band 3 Protein in Human Erythrocytes. Antioxidants, 9(8), 689. https://doi.org/10.3390/antiox9080689