
Plant-Produced S1 Subunit Protein of SARS-CoV-2 Elicits Immunogenic Responses in Mice
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of Plant Expression Vector for SARS-CoV-2 S1-Fc
2.2. SARS-CoV-2 S1-Fc Expression in N. benthamiana
2.3. Purification of SARS-CoV-2 S1-Fc Protein
2.4. Quantification of Plant-Produced SARS-CoV-2 S1-Fc Protein
2.5. Mice Immunization with the SARS-CoV-2 S1-Fc Protein
2.6. Evaluation of Immunological Responses in Mice
2.7. Statistical Analysis
3. Results
3.1. SARS-CoV-2 S1-Fc Expression in N. benthamiana
3.2. Purification and Characterization of SARS-CoV-2 S1-Fc from N. Benthamiana Leaves
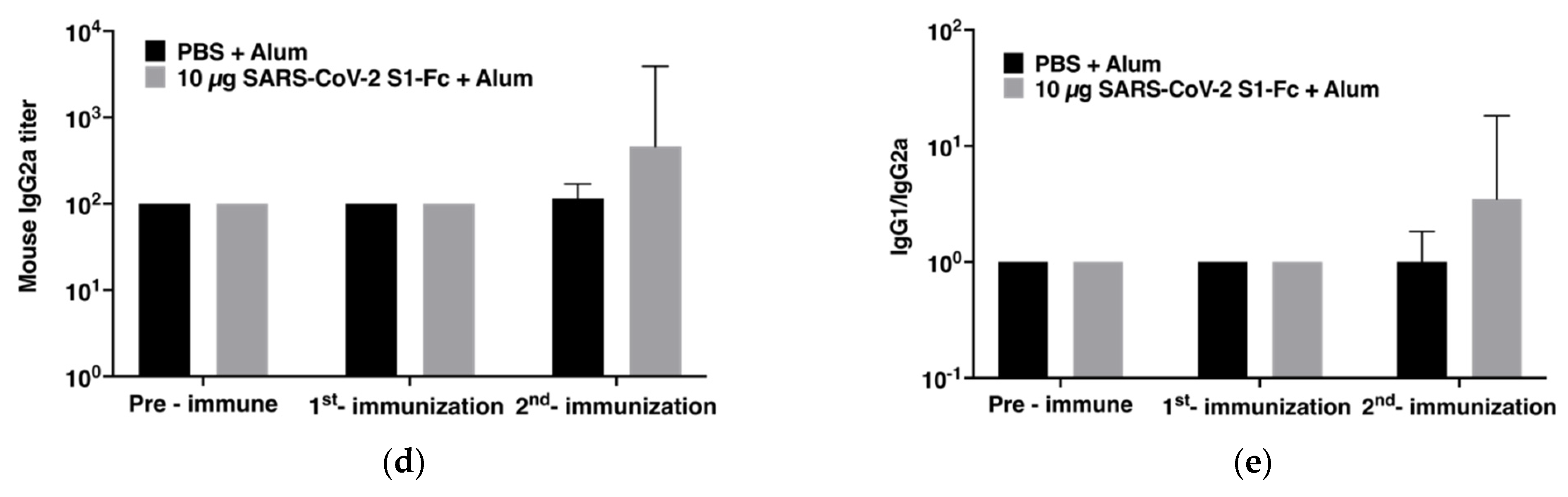
3.3. Immunogenicity in Mice
3.4. IFN-γ ELISpot Assay
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Origin of SARS-CoV-2, 26 March 2020; World Health Organization: Geneva, Switzerland, 2020.
- Banerjee, A.; Kulcsar, K.; Misra, V.; Frieman, M.; Mossman, K. Bats and Coronaviruses. Viruses 2019, 11, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letko, M.; Marzi, A.; Munster, V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat. Microbiol. 2020, 5, 562–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabi, F.A.; Al Zoubi, M.S.; Kasasbeh, G.A.; Salameh, D.M.; Al-Nasser, A.D. SARS-CoV-2 and Coronavirus Disease 2019: What We Know So Far. Pathogens 2020, 9, 231. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Leibowitz, J.L. The structure and functions of coronavirus genomic 3’ and 5’ ends. Virus Res. 2015, 206, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Li, F.; Shi, Z.L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Haynes, B.F.; Corey, L.; Fernandes, P.; Gilbert, P.B.; Hotez, P.J.; Rao, S.; Santos, M.R.; Schuitemaker, H.; Watson, M.; Arvin, A. Prospects for a safe COVID-19 vaccine. Sci. Transl. Med. 2020, 12, eabe0948. [Google Scholar] [CrossRef]
- Li, Y.; Tenchov, R.; Smoot, J.; Liu, C.; Watkins, S.; Zhou, Q. A Comprehensive Review of the Global Efforts on COVID-19 Vaccine Development. ACS Cent. Sci. 2021, 7, 512–533. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, H.; Tian, L.; Pang, Z.; Yang, Q.; Huang, T.; Fan, J.; Song, L.; Tong, Y.; Fan, H. COVID-19 vaccine development: Milestones, lessons and prospects. Signal Transduct. Target. Ther. 2022, 7, 146. [Google Scholar] [CrossRef]
- Dong, Y.; Dai, T.; Wei, Y.; Zhang, L.; Zheng, M.; Zhou, F. A systematic review of SARS-CoV-2 vaccine candidates. Signal Transduct. Target. Ther. 2020, 5, 237. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, C.; Xu, X.F.; Xu, W.; Liu, S.W. Structural and functional properties of SARS-CoV-2 spike protein: Potential antivirus drug development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.F.; Quadeer, A.A.; McKay, M.R. Preliminary Identification of Potential Vaccine Targets for the COVID-19 Coronavirus (SARS-CoV-2) Based on SARS-CoV Immunological Studies. Viruses 2020, 12, 254. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.; Ke, B.; Feng, Q.; Yang, D.; Lian, Q.; Li, Z.; Lu, L.; Ke, C.; Liu, Z.; Liao, G. Construction and immunogenic studies of a mFc fusion receptor binding domain (RBD) of spike protein as a subunit vaccine against SARS-CoV-2 infection. Chem. Commun. 2020, 56, 8683–8686. [Google Scholar] [CrossRef] [PubMed]
- Mamedov, T.; Yuksel, D.; Ilgin, M.; Gurbuzaslan, I.; Gulec, B.; Yetiskin, H.; Uygut, M.A.; Islam Pavel, S.T.; Ozdarendeli, A.; Mammadova, G.; et al. Plant-Produced Glycosylated and In Vivo Deglycosylated Receptor Binding Domain Proteins of SARS-CoV-2 Induce Potent Neutralizing Responses in Mice. Viruses 2021, 13, 1595. [Google Scholar] [CrossRef] [PubMed]
- Siriwattananon, K.; Manopwisedjaroen, S.; Shanmugaraj, B.; Rattanapisit, K.; Phumiamorn, S.; Sapsutthipas, S.; Trisiriwanich, S.; Prompetchara, E.; Ketloy, C.; Buranapraditkun, S.; et al. Plant-Produced Receptor-Binding Domain of SARS-CoV-2 Elicits Potent Neutralizing Responses in Mice and Non-human Primates. Front. Plant Sci. 2021, 12, 682953. [Google Scholar] [CrossRef] [PubMed]
- Brondyk, W.H. Chapter 11 Selecting an Appropriate Method for Expressing a Recombinant Protein. In Methods in Enzymology; Burgess, R.R., Deutscher, M.P., Eds.; Academic Press: New York, NY, USA, 2009; Volume 463, pp. 131–147. [Google Scholar]
- Shanmugaraj, B.; Phoolcharoen, W. Addressing demand for recombinant biopharmaceuticals in the COVID-19 era. Asian Pac. J. Trop. Med. 2021, 14, 49–51. [Google Scholar] [CrossRef]
- Shanmugaraj, B.; I. Bulaon, C.J.; Phoolcharoen, W. Plant Molecular Farming: A Viable Platform for Recombinant Biopharmaceutical Production. Plants 2020, 9, 842. [Google Scholar] [CrossRef]
- Capell, T.; Twyman, R.M.; Armario-Najera, V.; Ma, J.K.C.; Schillberg, S.; Christou, P. Potential Applications of Plant Biotechnology against SARS-CoV-2. Trends Plant Sci. 2020, 25, 635–643. [Google Scholar] [CrossRef]
- Fischer, R.; Buyel, J.F. Molecular farming—The slope of enlightenment. Biotechnol. Adv. 2020, 40, 107519. [Google Scholar] [CrossRef]
- Shanmugaraj, B.; Rattanapisit, K.; Manopwisedjaroen, S.; Thitithanyanont, A.; Phoolcharoen, W. Monoclonal Antibodies B38 and H4 Produced in Nicotiana benthamiana Neutralize SARS-CoV-2 in vitro. Front. Plant Sci. 2020, 11, 589995. [Google Scholar] [CrossRef]
- Rattanapisit, K.; Zhang, C.; Siriwattananon, K.; Huang, Z.; Phoolcharoen, W. Plant-Produced Anti-Enterovirus 71 (EV71) Monoclonal Antibody Efficiently Protects Mice Against EV71 Infection. Plants 2019, 8, 560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyriakidis, N.C.; López-Cortés, A.; González, E.V.; Grimaldos, A.B.; Prado, E.O. SARS-CoV-2 vaccines strategies: A comprehensive review of phase 3 candidates. NPJ Vaccines 2021, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Wørzner, K.; Sheward, D.J.; Schmidt, S.T.; Hanke, L.; Zimmermann, J.; McInerney, G.; Karlsson Hedestam, G.B.; Murrell, B.; Christensen, D.; Pedersen, G.K. Adjuvanted SARS-CoV-2 spike protein elicits neutralizing antibodies and CD4 T cell responses after a single immunization in mice. EBioMedicine 2021, 63, 103197. [Google Scholar] [CrossRef]
- Yang, Y.; Du, L. SARS-CoV-2 spike protein: A key target for eliciting persistent neutralizing antibodies. Signal Transduct. Target. Ther. 2021, 6, 95. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Bhatnagar, N.; Jeeva, S.; Oh, J.; Park, B.R.; Shin, C.H.; Wang, B.Z.; Kang, S.M. Immunogenicity and Neutralizing Activity Comparison of SARS-CoV-2 Spike Full-Length and Subunit Domain Proteins in Young Adult and Old-Aged Mice. Vaccines 2021, 9, 316. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Cao, H.; Liu, C. SARS-CoV-2 S1 is superior to the RBD as a COVID-19 subunit vaccine antigen. J. Med. Virol. 2021, 93, 892–898. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Shang, J.; Jiang, S.; Du, L. Subunit Vaccines Against Emerging Pathogenic Human Coronaviruses. Front. Microbiol. 2020, 11, 298. [Google Scholar] [CrossRef]
- Zhang, B.; Shanmugaraj, B.; Daniell, H. Expression and functional evaluation of biopharmaceuticals made in plant chloroplasts. Curr. Opin. Chem. Biol. 2017, 38, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Burnett, M.J.B.; Burnett, A.C. Therapeutic recombinant protein production in plants: Challenges and opportunities. Plants People Planet 2019, 2, 121–132. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Baysal, C.; Lobato Gómez, M.; Huang, X.; Alvarez, D.; Zhu, C.; Armario-Najera, V.; Blanco Perera, A.; Cerda Bennaser, P.; Saba-Mayoral, A.; et al. Contributions of the international plant science community to the fight against infectious diseases in humans—Part 2: Affordable drugs in edible plants for endemic and re-emerging diseases. Plant Biotechnol. J. 2021, 19, 1921–1936. [Google Scholar] [CrossRef]
- Lobato Gómez, M.; Huang, X.; Alvarez, D.; He, W.; Baysal, C.; Zhu, C.; Armario-Najera, V.; Blanco Perera, A.; Cerda Bennasser, P.; Saba-Mayoral, A.; et al. Contributions of the international plant science community to the fight against human infectious diseases—Part 1: Epidemic and pandemic diseases. Plant Biotechnol. J. 2021, 19, 1901–1920. [Google Scholar] [CrossRef] [PubMed]
- Daniell, H.; Singh, N.D.; Mason, H.; Streatfield, S.J. Plant-made vaccine antigens and biopharmaceuticals. Trends Plant Sci. 2009, 14, 669–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govea-Alonso, D.O.; Rybicki, E.; Rosales-Mendoza, S. Plant-Based Vaccines as a Global Vaccination Approach: Current Perspectives. Genet. Eng. Plants A Source Vaccines Against Wide Spread Dis. 2014, 265–280. [Google Scholar] [CrossRef]
- Schillberg, S.; Raven, N.; Spiegel, H.; Rasche, S.; Buntru, M. Critical Analysis of the Commercial Potential of Plants for the Production of Recombinant Proteins. Front. Plant Sci. 2019, 10, 720. [Google Scholar] [CrossRef] [Green Version]
- Rosales-Mendoza, S.; Márquez-Escobar, V.A.; González-Ortega, O.; Nieto-Gómez, R.; Arévalo-Villalobos, J.I. What Does Plant-Based Vaccine Technology Offer to the Fight against COVID-19? Vaccines 2020, 8, 183. [Google Scholar] [CrossRef] [Green Version]
- Maharjan, P.M.; Choe, S. Plant-Based COVID-19 Vaccines: Current Status, Design, and Development Strategies of Candidate Vaccines. Vaccines 2021, 9, 992. [Google Scholar] [CrossRef]
- Rattanapisit, K.; Yusakul, G.; Shanmugaraj, B.; Kittirotruji, K.; Suwatsrisakul, P.; Prompetchara, E.; Taychakhoonavud, S.; Phoolcharoen, W. Plant-produced recombinant SARS-CoV-2 receptor-binding domain; an economical, scalable biomaterial source for COVID-19 diagnosis. Biomater. Transl. 2021, 2, 43–49. [Google Scholar] [CrossRef]
- Shanmugaraj, B.; Siriwattananon, K.; Malla, A.; Phoolcharoen, W. Potential for Developing Plant-Derived Candidate Vaccines and Biologics against Emerging Coronavirus Infections. Pathogens 2021, 10, 1051. [Google Scholar] [CrossRef]
- Shanmugaraj, B.; Khorattanakulchai, N.; Phoolcharoen, W. Chapter 12—SARS-CoV-2 Vaccines: Current Trends and Prospects of Developing Plant-Derived Vaccines. In Biomedical Innovations to Combat COVID-19; Rosales-Mendoza, S., Comas-Garcia, M., Gonzalez-Ortega, O., Eds.; Academic Press: New York, NY, USA, 2022; pp. 213–229. [Google Scholar]
- Ortega-Berlanga, B.; Pniewski, T. Plant-Based Vaccines in Combat against Coronavirus Diseases. Vaccines 2022, 10, 138. [Google Scholar] [CrossRef]
- Shanmugaraj, B.; Khorattanakulchai, N.; Panapitakkul, C.; Malla, A.; Im-erbsin, R.; Inthawong, M.; Sunyakumthorn, P.; Hunsawong, T.; Klungthong, C.; Reed, M.C.; et al. Preclinical evaluation of a plant-derived SARS-CoV-2 subunit vaccine: Protective efficacy, immunogenicity, safety, and toxicity. Vaccine 2022, 40, 4440–4452. [Google Scholar] [CrossRef]
- Yao, J.; Weng, Y.; Dickey, A.; Wang, K.Y. Plants as Factories for Human Pharmaceuticals: Applications and Challenges. Int. J. Mol. Sci. 2015, 16, 28549–28565. [Google Scholar] [CrossRef] [PubMed]
- Awate, S.; Babiuk, L.A.; Mutwiri, G. Mechanisms of action of adjuvants. Front. Immunol. 2013, 4, 114. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panapitakkul, C.; Khorattanakulchai, N.; Rattanapisit, K.; Srisangsung, T.; Shanmugaraj, B.; Buranapraditkun, S.; Ketloy, C.; Prompetchara, E.; Phoolcharoen, W. Plant-Produced S1 Subunit Protein of SARS-CoV-2 Elicits Immunogenic Responses in Mice. Vaccines 2022, 10, 1961. https://doi.org/10.3390/vaccines10111961
Panapitakkul C, Khorattanakulchai N, Rattanapisit K, Srisangsung T, Shanmugaraj B, Buranapraditkun S, Ketloy C, Prompetchara E, Phoolcharoen W. Plant-Produced S1 Subunit Protein of SARS-CoV-2 Elicits Immunogenic Responses in Mice. Vaccines. 2022; 10(11):1961. https://doi.org/10.3390/vaccines10111961
Chicago/Turabian StylePanapitakkul, Chalisa, Narach Khorattanakulchai, Kaewta Rattanapisit, Theerakarn Srisangsung, Balamurugan Shanmugaraj, Supranee Buranapraditkun, Chutitorn Ketloy, Eakachai Prompetchara, and Waranyoo Phoolcharoen. 2022. "Plant-Produced S1 Subunit Protein of SARS-CoV-2 Elicits Immunogenic Responses in Mice" Vaccines 10, no. 11: 1961. https://doi.org/10.3390/vaccines10111961
APA StylePanapitakkul, C., Khorattanakulchai, N., Rattanapisit, K., Srisangsung, T., Shanmugaraj, B., Buranapraditkun, S., Ketloy, C., Prompetchara, E., & Phoolcharoen, W. (2022). Plant-Produced S1 Subunit Protein of SARS-CoV-2 Elicits Immunogenic Responses in Mice. Vaccines, 10(11), 1961. https://doi.org/10.3390/vaccines10111961