
Pitfalls in the Application of Dispase-Based Keratinocyte Dissociation Assay for In Vitro Analysis of Pemphigus Vulgaris

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Antibodies
- HaCaT spontaneously immortalized human keratinocyte cell line (kindly provided by Prof. P. Boukamp, formerly DKFZ Heidelberg)
- HaSKpw spontaneously immortalized human keratinocyte cell line (kindly provided by Prof. P. Boukamp, formerly DKFZ Heidelberg)
- Primary human keratinocytes (ethical approval: 547/2011BO2)
- AK23 anti-Dsg3 antibody (hybridoma cell supernatants were kindly provided by Prof. R. Tikkanen, Justus Liebig University Giessen, Germany, and purification was performed following Beckert et al. [20])
- Human immunoglobulin isotype control (Thermo Fisher Scientific Inc., Waltham, MA, USA, Cat. no. 02-7102)
2.2. Reagents and Equipment
- 24-Well Cell Culture Multiwell Plate(sterile, with lid) (Greiner AG, Kremsmünster, Austria; Cat. no. 10177380)
- Antibiotics/antimycotics (100×) (Thermo Fisher Scientific Inc., Waltham, MA, USA, Cat. No. 15240062)
- Calcium chloride (CaCl2) (Carl Roth, Karlsruhe, Germany, Cat. No. CN93.1)
- Chelex 100 Chelating Resin, analytical grade, 100–200 mesh, sodium form (Bio-Rad Laboratories Inc., Hercules, CA, USA, Cat. No. 1422832)
- ChemiDoc Imaging System (Bio-Rad Laboratories Inc., Hercules, CA, USA, Cat. No. 1708195)
- CnT-Prime Epithelial Proliferation Medium (CELLnTEC ADVANCED CELL SYSTEMS AG, Bern, Switzerland, Cat. No. CnT-PR)
- Dispase II powder (Thermo Fisher Scientific Inc., Waltham, MA, USA, Cat. No. 17105041)
- Dulbecco’s Modified Eagle Medium (DMEM) (PAN-Biotech GmbH, Aidenbach, Germany, Cat. No. P04-04515)
- Eppendorf Xplorer® electrical pipette (Eppendorf AG, Hamburg, Germany, Cat. No. 4861000040)
- Fetal Bovine Serum (FBS) standard (PAN-Biotech GmbH, Aidenbach, Germany, Cat. No. P30-3306), is equivalent toFetal Calf Serum (FCS)
- Hanks’ Balanced Salt Solution (HBSS) with Ca2+ (Thermo Fisher Scientific Inc., Waltham, MA, USA, Cat. No. 14025-050)
- Syringe filters, ROTILABO® PVDF, 0.22 µm (Carl Roth, Karlsruhe, Germany, Cat. No. P666.1)
- Thiazolyl blue tetrazolium bromide (MTT) (Invitrogen by Thermo Fisher Scientific Inc., Waltham, MA, USA, Cat. No. M6494)
- Whatman® Grade 1 Qualitative Filter Paper (Merck KGaA, Darmstadt, Germany, Cat. No. WHA1001325)
2.3. Reagents Setup
- Chelated FCS: Swell 100 g of Chelex 100 Chelating Resin in 500 mL of dH2O. Adjust the pH to 7.4 with HCl while stirring (pH will take a while to stabilize during titration). Filter through Whatman® Grade 1 Qualitative Filter Paper. Scrape resin slurry into 500 mL of FCS and stir at room temperature for 3 h or at 4 °C overnight. Filter the chelated FCS through Whatman® Grade 1 Qualitative Filter Paper and discard the resin slurry. Filter the chelated FCS through a 0.22 µm filter to sterilize it. Aliquot sterile chelated FCS and store it at −20 °C.
- CnT-Prime Epithelial Proliferation Medium supplemented with 1% antibiotics/antimycotics: 5 mL of antibiotics/antimycotics per 500 mL bottle of medium.
- Dispase solution: dissolve the dispase in HBSS buffer to a final concentration of 2.5 U/mL.
- DMEM supplemented with 10% FCS: 50 mL of FCS per 500 mL bottle of medium.
- DMEM supplemented with 8% chelated FCS: 40 mL of chelated FCS per 500 mL bottle of medium.
- MTT staining solution: a 5 mg/mL working solution is prepared by dissolving MTT in HBSS buffer.
2.4. Software
- ImageJ (Rasband, W.S., ImageJ, US National Institutes of Health, Bethesda, MD, USA, https://imagej.nih.gov/ij/, 1997–2021; accessed on 6 December 2021)
- Image Lab Software (Bio-Rad Laboratories Inc., Hercules, CA, USA, Cat. No. 1708195)
- Microsoft Excel (Microsoft Corporation, Redmond, Washington, DC, USA, version 2016)
2.5. Methods and Protocols
2.5.1. Cell Culture
2.5.2. Dispase-Based Keratinocyte Dissociation Assay in Primary Keratinocytes
Generation of a Cell Monolayer, Time Consideration: 2–3 Days
- Dilute 6 × 105 cells/well in keratinocyte medium (CnT-Prime Epithelial Proliferation Medium) without FCS supplemented with antibiotics/antimycotics. Plate 1 mL of cell suspension per well in a sterile 24-well plate.
- Incubate the cells at 37 °C in 5% CO2 atmosphere. The cells are usually confluent by 24–48 h. Medium should be changed every day.
- When the cells reach confluency, aspirate the medium and replace with a new medium (CnT-Prime Epithelial Proliferation Medium) containing 1.8 mM of CaCl2.
- Incubate the cells at 37 °C in 5% CO2 atmosphere for an additional 24 h.
Stimulation of the Monolayer, Time Consideration: Depending on the Stimulation Procedure
- Aspirate the medium and add the stimulus diluted in medium (CnT-Prime Epithelial Proliferation Medium) without supplemented CaCl2.
Detaching of the Monolayer from the Cell Culture Dish, Time Consideration: Approximately 50 min
- Aspirate the medium and rinse the cell layer with 500 µL of HBSS.
- Add 350 µL of dispase solution to each well.
- Incubate the plate for 30–40 min at 37 °C in 5% CO2 atmosphere. The monolayer should be completely detached from the dish bottom.
- Aspirate dispase solution with a pipette and add 350 µL of HBSS into each well.
- Add 20 µL of MTT staining solution per well and incubate for 10–15 min at 37 °C in 5% CO2 atmosphere.
- Remove the staining solution and add 350 µL of HBSS per well.
- Apply mechanical stress by pipetting with an electrical pipette (settings: Pip, speed 8, 300 µL).
2.5.3. Dispase-Based Keratinocyte Dissociation Assay in HaCaT and HaSKpw Keratinocytes
Generation of a Cell Monolayer, Time Consideration: 1 Day (HaCaT) or 2–3 Days (HaSKpw)
- Dilute 6 × 105 cells/well in DMEM supplemented with:
- -
- Chelated FCS (1.6 mM final concentration of Ca2+) for HaCaT cell culture
- -
- Standard FCS (1.9 mM final concentration of Ca2+) for HaSKpw cell culture
Stimulation of the Monolayer, Time Consideration: Depending on the Stimulation Procedure
- Aspirate the medium and add the stimulus diluted in the respective medium:
- -
- Chelated FCS (1.6 mM final concentration of Ca2+) for HaCaT cells
- -
- Standard FCS (1.9 mM final concentration of Ca2+) for HaSKpw cells
- -
- Either human IgG human isotype control (IgG), 20 µg/mL, or AK23 anti-Dsg3 antibody, 20 µg/mL, at 37 °C in 5% CO2 atmosphere for 4 h for HaCaT cells
- -
- Either human IgG human isotype control (IgG), 30 µg/mL, or AK23 anti-Dsg3 antibody, 30 µg/mL, at 37 °C in 5% CO2 atmosphere for 4 h for HaSKpw cells
Detaching of the Monolayer from the Cell Culture Dish, Time Consideration: Approximately 50 min
- Aspirate the medium and rinse the cell layer with 500 µL of HBSS.
- Add 350 µL of dispase solution to each well.
- Incubate the plate at 37 °C in 5% CO2 atmosphere for:
- -
- Approximately 40 min (HaCaT cells)
- -
- Approximately 30 min (HaSKpw cells)
Application of Mechanical Stress, Time Consideration: Approximately 30 min
- Aspirate dispase solution carefully with a pipette and apply 350 µL of HBSS in each well.
- Add 20 µL of MTT staining solution per well and incubate for 10–15 min at 37 °C in 5% CO2 atmosphere:
- -
- Approximately 10 min for HaCaT monolayer
- -
- Approximately 20 min for HaSKpw monolayer
- Carefully remove the staining solution and add 350 µL of HBSS per well.
- Apply mechanical stress by pipetting with an electrical pipette (settings: Pip, speed 8, 300 µL).
2.5.4. Application of Mechanical Stress, Quantification of Fragments and Data Processing
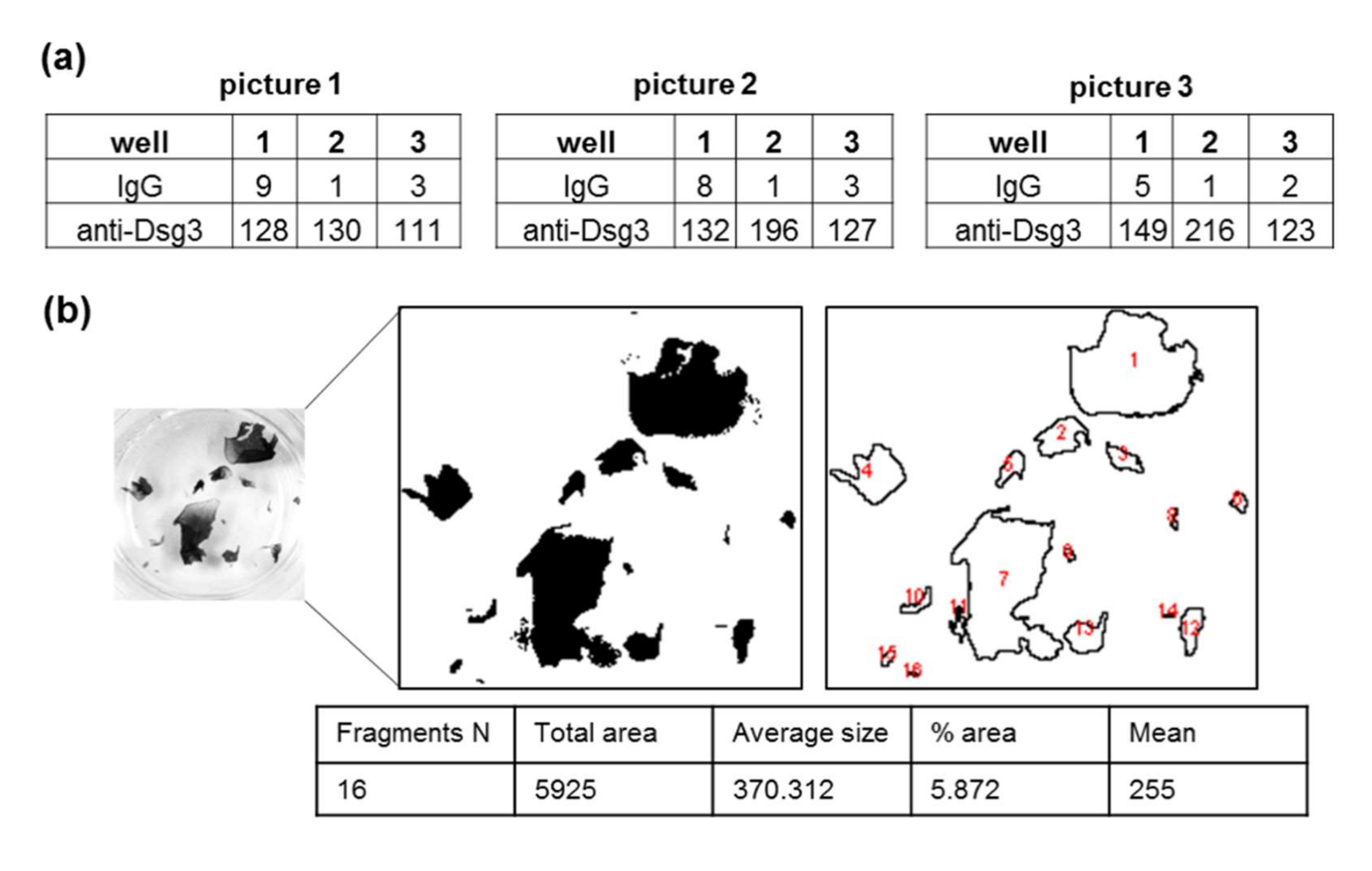
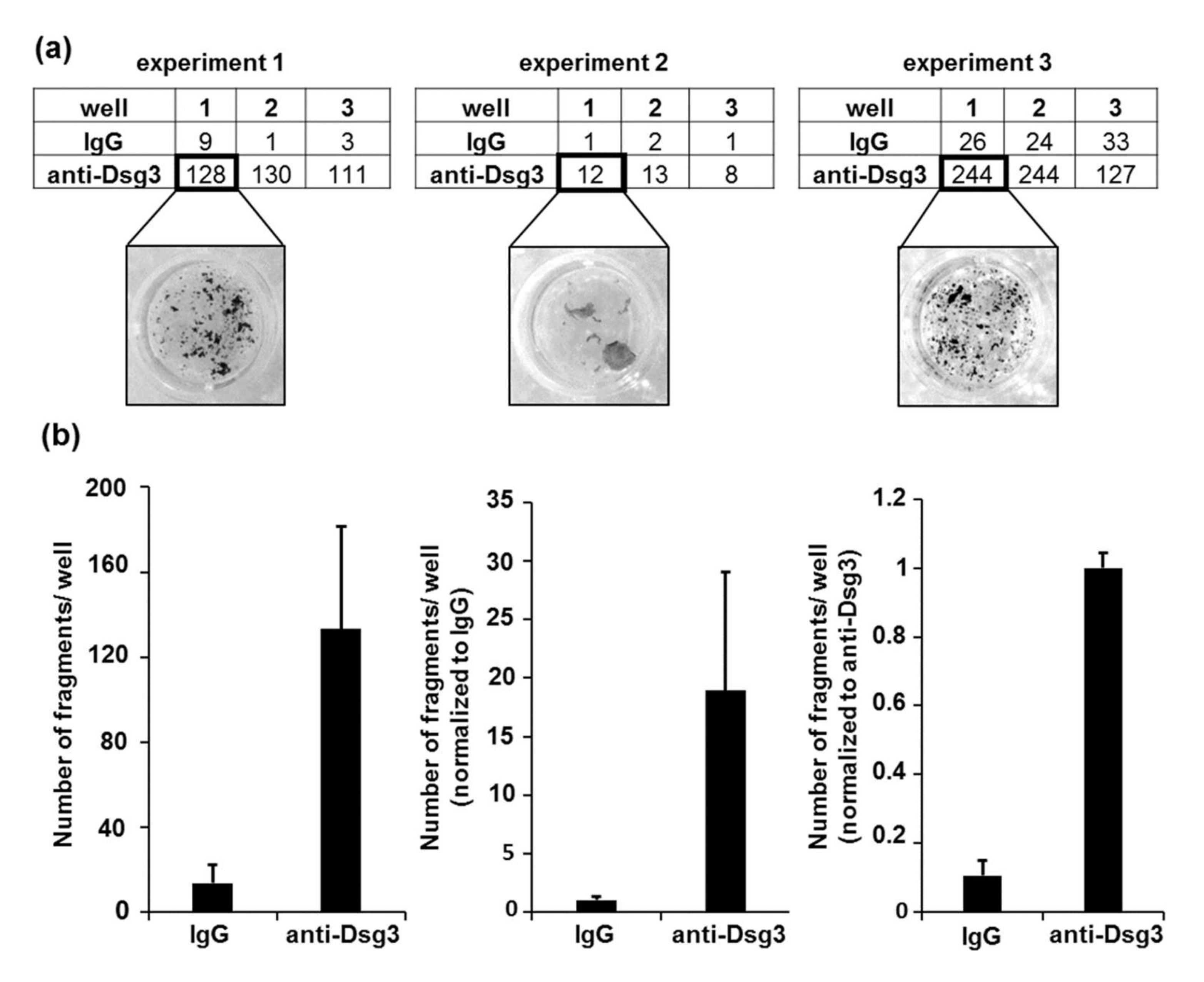
- Set the pipetting conditions by first applying the defined mechanical stress to the replicates within the positive control condition (in this experimental setting, anti-Dsg3-treated samples were used as a control). Take the highest number of pipetting steps necessary to fragment the monolayer between the respective replicates (at least two or more fragments) and apply the same pipetting number to all samples.
- Take at least three pictures of each well (Chemilum Photo Chamber, Image Lab Software).
- 3.
- Quantify fragments with imaging software, i.e., ImageJ software:
- a.
- Open the image with ImageJ.
- b.
- Mark the target area with the desired shape and copy.
- c.
- Create a new file and insert the copied file (Menu: File -> New (choose white background) -> Image -> insert)
- d.
- Adjust the threshold of the image by marking only the cell layer fragments (Menu: Image -> Adjust -> Threshold: adjust)
- e.
- Count the particles (Menu: Analyze -> Analyze Particles: Define the size adapted to the requirements of the individual cell lines (e.g., 5–infinity). Tick: display results, clear results, summarize).
- f.
- Data are evaluated with Microsoft Excel software.
3. Results
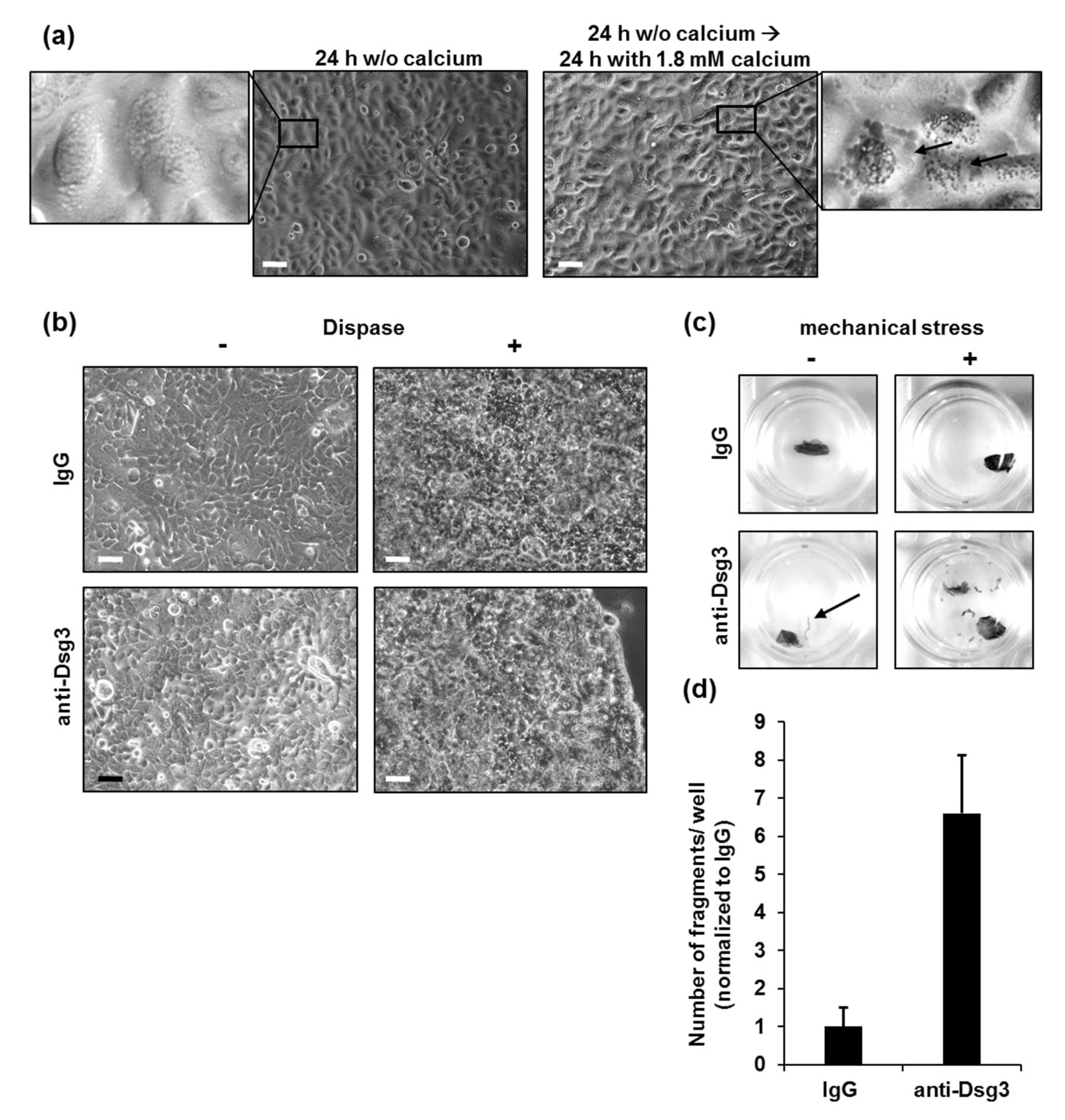
3.1. Conditions for Cultivation of Primary Human Keratinocytes for Dispase-Based Keratinocyte Dissociation Assay
3.2. Conditions for Cultivation of HaCaT Cells for Dispase-Based Keratinocyte Dissociation Assay
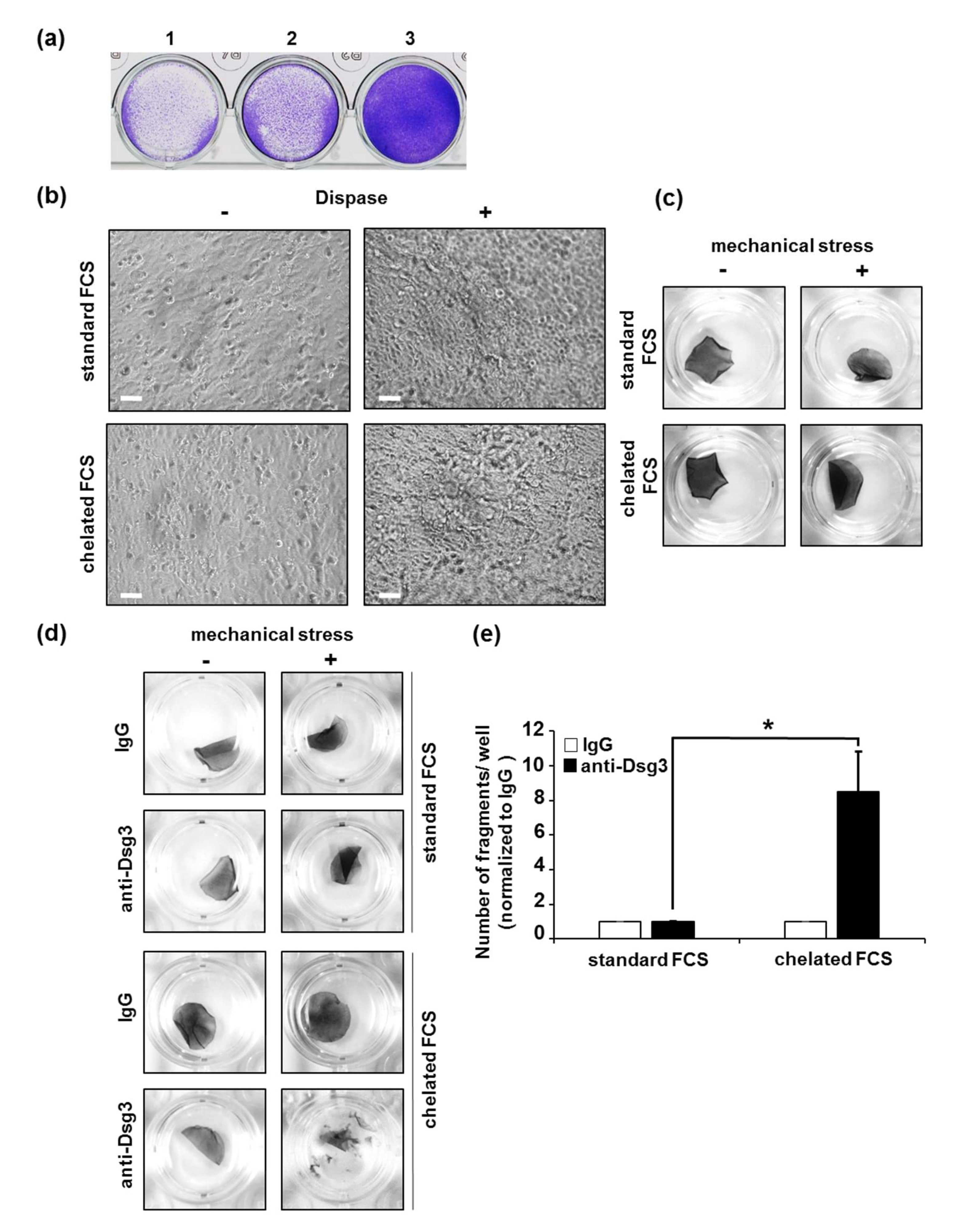
3.3. Conditions for Cultivation of HaSKpw Cells for Dispase-Based Keratinocyte Dissociation Assay
3.4. Application of Mechanical Stress, Quantification of Fragments, and Data Processing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kasperkiewicz, M.; Ellebrecht, C.T.; Takahashi, H.; Yamagami, J.; Zillikens, D.; Payne, A.S.; Amagai, M. Pemphigus. Nat. Rev. Dis. Primers 2017, 3, 17026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porro, A.M.; Seque, C.A.; Ferreira, M.C.C.; Enokihara, M. Pemphigus vulgaris. An. Bras. Dermatol. 2019, 94, 264–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrod, D.; Chidgey, M.; North, A. Desmosomes: Differentiation, development, dynamics and disease. Curr. Opin. Cell. Biol. 1996, 8, 670–678. [Google Scholar] [CrossRef]
- Waschke, J. The desmosome and pemphigus. Histochem. Cell. Biol. 2008, 130, 21–54. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.L.; Najor, N.A.; Green, K.J. Desmosomes: Regulators of cellular signaling and adhesion in epidermal health and disease. Cold Spring Harb Perspect Med. 2014, 4, a015297. [Google Scholar] [CrossRef] [Green Version]
- Green, K.J.; Simpson, C.L. Desmosomes: New perspectives on a classic. J. Invest. Dermatol. 2007, 127, 2499–2515. [Google Scholar] [CrossRef] [Green Version]
- Kottke, M.D.; Delva, E.; Kowalczyk, A.P. The desmosome: Cell science lessons from human diseases. J. Cell Sci. 2006, 119, 797–806. [Google Scholar] [CrossRef] [Green Version]
- Amagai, M.; Koch, P.J.; Nishikawa, T.; Stanley, J.R. Pemphigus vulgaris antigen (desmoglein 3) is localized in the lower epidermis, the site of blister formation in patients. J. Invest. Dermatol. 1996, 106, 351–355. [Google Scholar] [CrossRef]
- Mahoney, M.G.; Hu, Y.; Brennan, D.; Bazzi, H.; Christiano, A.M.; Wahl, J.K., 3rd. Delineation of diversified desmoglein distribution in stratified squamous epithelia: Implications in diseases. Exp. Dermatol. 2006, 15, 101–109. [Google Scholar] [CrossRef]
- Heupel, W.M.; Zillikens, D.; Drenckhahn, D.; Waschke, J. Pemphigus vulgaris IgG directly inhibit desmoglein 3-mediated transinteraction. J. Immunol. 2008, 181, 1825–1834. [Google Scholar] [CrossRef] [Green Version]
- Vielmuth, F.; Spindler, V.; Waschke, J. Atomic Force Microscopy Provides New Mechanistic Insights into the Pathogenesis of Pemphigus. Front. Immunol. 2018, 9, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilgram, G.F.; Caulfield, J.B.; Lever, W.F. An electron microscopic study of acantholysis in pemphigus vulgaris. J. Invest. Dermatol. 1961, 36, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Spindler, V.; Waschke, J. Pemphigus-A Disease of Desmosome Dysfunction Caused by Multiple Mechanisms. Front. Immunol. 2018, 9, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calautti, E.; Cabodi, S.; Stein, P.L.; Hatzfeld, M.; Kedersha, N.; Paolo Dotto, G. Tyrosine phosphorylation and src family kinases control keratinocyte cell-cell adhesion. J. Cell Biol. 1998, 141, 1449–1465. [Google Scholar] [CrossRef]
- Caldelari, R.; de Bruin, A.; Baumann, D.; Suter, M.M.; Bierkamp, C.; Balmer, V.; Muller, E. A central role for the armadillo protein plakoglobin in the autoimmune disease pemphigus vulgaris. J. Cell Biol. 2001, 153, 823–834. [Google Scholar] [CrossRef] [Green Version]
- Huen, A.C.; Park, J.K.; Godsel, L.M.; Chen, X.; Bannon, L.J.; Amargo, E.V.; Hudson, T.Y.; Mongiu, A.K.; Leigh, I.M.; Kelsell, D.P.; et al. Intermediate filament-membrane attachments function synergistically with actin-dependent contacts to regulate intercellular adhesive strength. J. Cell Biol. 2002, 159, 1005–1017. [Google Scholar] [CrossRef] [Green Version]
- Ishii, K.; Harada, R.; Matsuo, I.; Shirakata, Y.; Hashimoto, K.; Amagai, M. In vitro keratinocyte dissociation assay for evaluation of the pathogenicity of anti-desmoglein 3 IgG autoantibodies in pemphigus vulgaris. J Invest. Dermatol. 2005, 124, 939–946. [Google Scholar] [CrossRef] [Green Version]
- Payne, A.S.; Ishii, K.; Kacir, S.; Lin, C.; Li, H.; Hanakawa, Y.; Tsunoda, K.; Amagai, M.; Stanley, J.R.; Siegel, D.L. Genetic and functional characterization of human pemphigus vulgaris monoclonal autoantibodies isolated by phage display. J. Clin. Invest. 2005, 115, 888–899. [Google Scholar] [CrossRef] [Green Version]
- Pavez Lorie, E.; Stricker, N.; Plitta-Michalak, B.; Chen, I.P.; Volkmer, B.; Greinert, R.; Jauch, A.; Boukamp, P.; Rapp, A. Characterisation of the novel spontaneously immortalized and invasively growing human skin keratinocyte line HaSKpw. Sci. Rep. 2020, 10, 15196. [Google Scholar] [CrossRef]
- Beckert, B.; Panico, F.; Pollmann, R.; Eming, R.; Banning, A.; Tikkanen, R. Immortalized Human hTert/KER-CT Keratinocytes a Model System for Research on Desmosomal Adhesion and Pathogenesis of Pemphigus Vulgaris. Int J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Feoktistova, M.; Geserick, P.; Kellert, B.; Dimitrova, D.P.; Langlais, C.; Hupe, M.; Cain, K.; MacFarlane, M.; Hacker, G.; Leverkus, M. cIAPs block Ripoptosome formation, a RIP1/caspase-8 containing intracellular cell death complex differentially regulated by cFLIP isoforms. Mol. Cell 2011, 43, 449–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennings, H.; Michael, D.; Cheng, C.; Steinert, P.; Holbrook, K.; Yuspa, S.H. Calcium regulation of growth and differentiation of mouse epidermal cells in culture. Cell 1980, 19, 245–254. [Google Scholar] [CrossRef]
- Penn, E.J.; Burdett, I.D.; Hobson, C.; Magee, A.I.; Rees, D.A. Structure and assembly of desmosome junctions: Biosynthesis and turnover of the major desmosome components of Madin-Darby canine kidney cells in low calcium medium. J. Cell Biol. 1987, 105, 2327–2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartle, E.I.; Urner, T.M.; Raju, S.S.; Mattheyses, A.L. Desmoglein 3 Order and Dynamics in Desmosomes Determined by Fluorescence Polarization Microscopy. Biophys. J. 2017, 113, 2519–2529. [Google Scholar] [CrossRef] [Green Version]
- Watt, F.M.; Mattey, D.L.; Garrod, D.R. Calcium-induced reorganization of desmosomal components in cultured human keratinocytes. J. Cell Biol. 1984, 99, 2211–2215. [Google Scholar] [CrossRef]
- Gutowska-Owsiak, D.; de La Serna, J.B.; Fritzsche, M.; Naeem, A.; Podobas, E.I.; Leeming, M.; Colin-York, H.; O’Shaughnessy, R.; Eggeling, C.; Ogg, G.S. Orchestrated control of filaggrin-actin scaffolds underpins cornification. Cell Death Dis. 2018, 9, 412. [Google Scholar] [CrossRef]
- Fujisaki, H.; Futaki, S.; Yamada, M.; Sekiguchi, K.; Hayashi, T.; Ikejima, T.; Hattori, S. Respective optimal calcium concentrations for proliferation on type I collagen fibrils in two keratinocyte line cells, HaCaT and FEPE1L-8. Regen. Ther. 2018, 8, 73–79. [Google Scholar] [CrossRef]
- Lichti, U.; Anders, J.; Yuspa, S.H. Isolation and short-term culture of primary keratinocytes, hair follicle populations and dermal cells from newborn mice and keratinocytes from adult mice for in vitro analysis and for grafting to immunodeficient mice. Nat. Protoc. 2008, 3, 799–810. [Google Scholar] [CrossRef]
- Walter, E.; Vielmuth, F.; Wanuske, M.T.; Seifert, M.; Pollmann, R.; Eming, R.; Waschke, J. Role of Dsg1- and Dsg3-Mediated Signaling in Pemphigus Autoantibody-Induced Loss of Keratinocyte Cohesion. Front. Immunol. 2019, 10, 1128. [Google Scholar] [CrossRef] [Green Version]
- Setzer, S.V.; Calkins, C.C.; Garner, J.; Summers, S.; Green, K.J.; Kowalczyk, A.P. Comparative analysis of armadillo family proteins in the regulation of a431 epithelial cell junction assembly, adhesion and migration. J. Invest. Dermatol. 2004, 123, 426–433. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, R.P.; Amargo, E.V.; Somasundaram, A.; Simpson, C.L.; Prakriya, M.; Denning, M.F.; Green, K.J. The calcium ATPase SERCA2 regulates desmoplakin dynamics and intercellular adhesive strength through modulation of PKCα signaling. FASEB J. 2011, 25, 990–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, M.D.; Kang, T.J.; Lee, C.H.; Lee, A.Y.; Noh, M. HaCaT Keratinocytes and Primary Epidermal Keratinocytes Have Different Transcriptional Profiles of Cornified Envelope-Associated Genes to T Helper Cell Cytokines. Biomol. Ther. 2012, 20, 171–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayflick, L. The Limited in Vitro Lifetime of Human Diploid Cell Strains. Exp. Cell Res. 1965, 37, 614–636. [Google Scholar] [CrossRef]
- Kaur, G.; Dufour, J.M. Cell lines: Valuable tools or useless artifacts. Spermatogenesis 2012, 2, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Boukamp, P.; Popp, S.; Altmeyer, S.; Hulsen, A.; Fasching, C.; Cremer, T.; Fusenig, N.E. Sustained nontumorigenic phenotype correlates with a largely stable chromosome content during long-term culture of the human keratinocyte line HaCaT. Genes Chromosomes Cancer 1997, 19, 201–214. [Google Scholar] [CrossRef]
- O’Keefe, E.J.; Briggaman, R.A.; Herman, B. Calcium-induced assembly of adherens junctions in keratinocytes. J. Cell Biol. 1987, 105, 807–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, J.E.; Jensen, P.J.; Wheelock, M.J. Cadherin function is required for human keratinocytes to assemble desmosomes and stratify in response to calcium. J. Invest. Dermatol. 1994, 102, 870–877. [Google Scholar] [CrossRef] [Green Version]
- Elias, P.M.; Nau, P.; Hanley, K.; Cullander, C.; Crumrine, D.; Bench, G.; Sideras-Haddad, E.; Mauro, T.; Williams, M.L.; Feingold, K.R. Formation of the epidermal calcium gradient coincides with key milestones of barrier ontogenesis in the rodent. J. Invest. Dermatol. 1998, 110, 399–404. [Google Scholar] [CrossRef] [Green Version]
- Elias, P.; Ahn, S.; Brown, B.; Crumrine, D.; Feingold, K.R. Origin of the epidermal calcium gradient: Regulation by barrier status and role of active vs passive mechanisms. J. Invest. Dermatol. 2002, 119, 1269–1274. [Google Scholar] [CrossRef] [Green Version]
- Micallef, L.; Belaubre, F.; Pinon, A.; Jayat-Vignoles, C.; Delage, C.; Charveron, M.; Simon, A. Effects of extracellular calcium on the growth-differentiation switch in immortalized keratinocyte HaCaT cells compared with normal human keratinocytes. Exp. Dermatol. 2009, 18, 143–151. [Google Scholar] [CrossRef]
- Cirillo, N.; Lanza, A.; Prime, S.S. Induction of hyper-adhesion attenuates autoimmune-induced keratinocyte cell-cell detachment and processing of adhesion molecules via mechanisms that involve PKC. Exp. Cell Res. 2010, 316, 580–592. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, R.P.; Green, K.J. Desmoplakin regulates desmosome hyperadhesion. J. Invest. Dermatol. 2012, 132, 482–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spindler, V.; Endlich, A.; Hartlieb, E.; Vielmuth, F.; Schmidt, E.; Waschke, J. The extent of desmoglein 3 depletion in pemphigus vulgaris is dependent on Ca(2+)-induced differentiation: A role in suprabasal epidermal skin splitting? Am. J. Pathol. 2011, 179, 1905–1916. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmidt, M.F.; Feoktistova, M.; Panayotova-Dimitrova, D.; Eichkorn, R.A.; Yazdi, A.S. Pitfalls in the Application of Dispase-Based Keratinocyte Dissociation Assay for In Vitro Analysis of Pemphigus Vulgaris. Vaccines 2022, 10, 208. https://doi.org/10.3390/vaccines10020208
Schmidt MF, Feoktistova M, Panayotova-Dimitrova D, Eichkorn RA, Yazdi AS. Pitfalls in the Application of Dispase-Based Keratinocyte Dissociation Assay for In Vitro Analysis of Pemphigus Vulgaris. Vaccines. 2022; 10(2):208. https://doi.org/10.3390/vaccines10020208
Chicago/Turabian StyleSchmidt, Morna F., Maria Feoktistova, Diana Panayotova-Dimitrova, Ramona A. Eichkorn, and Amir S. Yazdi. 2022. "Pitfalls in the Application of Dispase-Based Keratinocyte Dissociation Assay for In Vitro Analysis of Pemphigus Vulgaris" Vaccines 10, no. 2: 208. https://doi.org/10.3390/vaccines10020208
APA StyleSchmidt, M. F., Feoktistova, M., Panayotova-Dimitrova, D., Eichkorn, R. A., & Yazdi, A. S. (2022). Pitfalls in the Application of Dispase-Based Keratinocyte Dissociation Assay for In Vitro Analysis of Pemphigus Vulgaris. Vaccines, 10(2), 208. https://doi.org/10.3390/vaccines10020208