PCV7- and PCV10-Vaccinated Otitis-Prone Children in New Zealand Have Similar Pneumococcal and Haemophilus influenzae Densities in Their Nasopharynx and Middle Ear


Abstract
:1. Introduction
2. Methods
2.1. Nasopharyngeal Swabs and Middle Ear Effusion Samples
2.2. DNA Extraction
2.3. Quantitative PCR
2.4. Statistical Analysis
2.5. Ethics
3. Results
3.1. Study Population
3.2. No Difference in Otopathogen Presence in the Nasopharynx of PCV7- and PCV10-Vaccinated Otitis-Prone or Non-Otitis-Prone Children
3.3. PCV10-Vaccinated Otitis-Prone and Non-Otitis-Prone Children Had Similar NTHi Nasopharyngeal Densities to PCV7-Vaccinated Otitis-Prone and Non-Otitis-Prone Children
3.4. PCV10-Vaccinated Otitis-Prone Children Had Higher Pneumococcal Nasopharyngeal Densities than PCV7-Vaccinated Otitis-Prone Children
3.5. Otitis-Prone Children Had Higher Densities of NTHi and S. Pneumoniae in Their Nasopharynx Compared with Non-Otitis-Prone Children
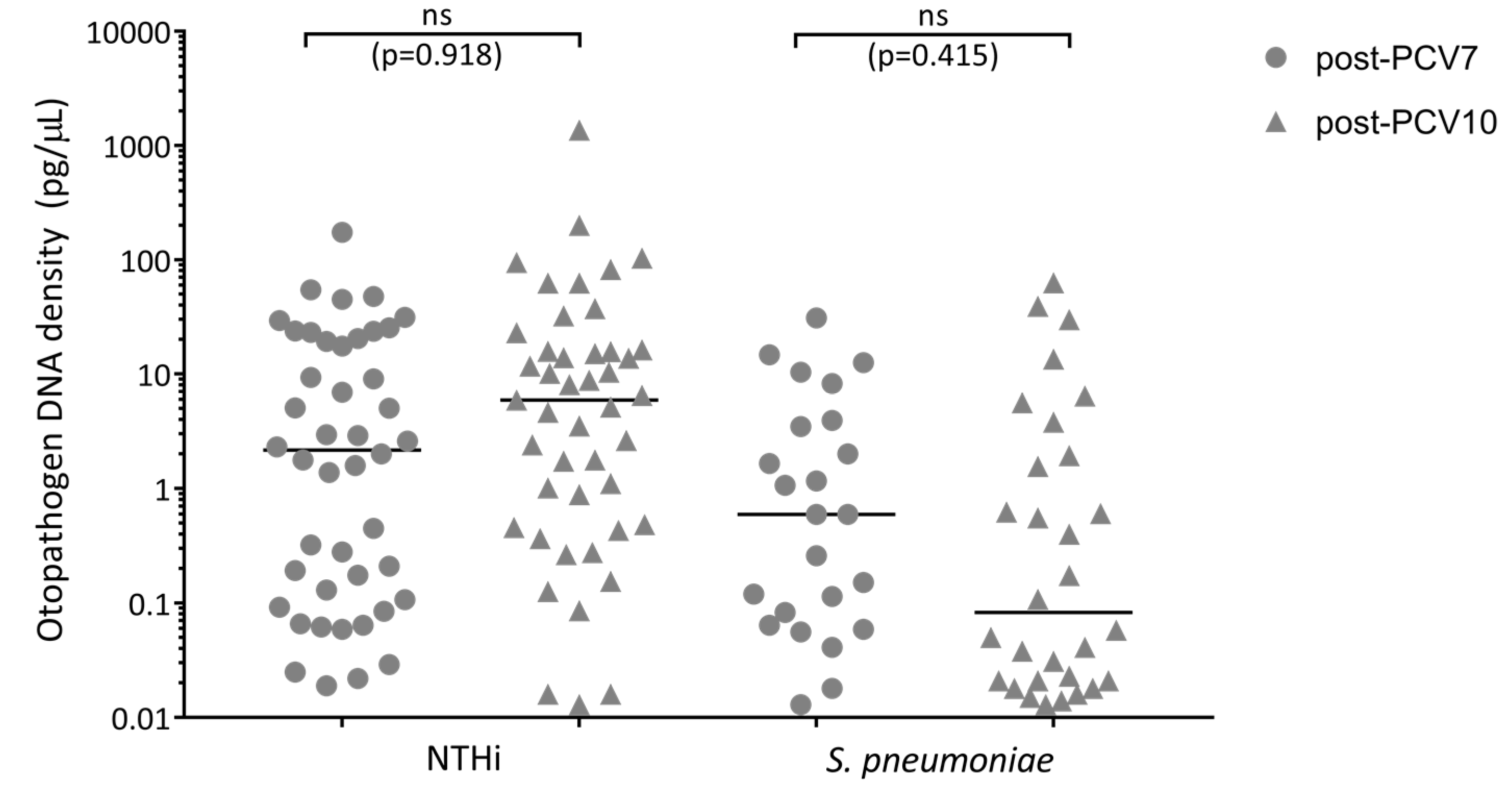
3.6. Otopathogen Density in the Middle Ear of Otitis-Prone Children was Similar between Vaccine Groups
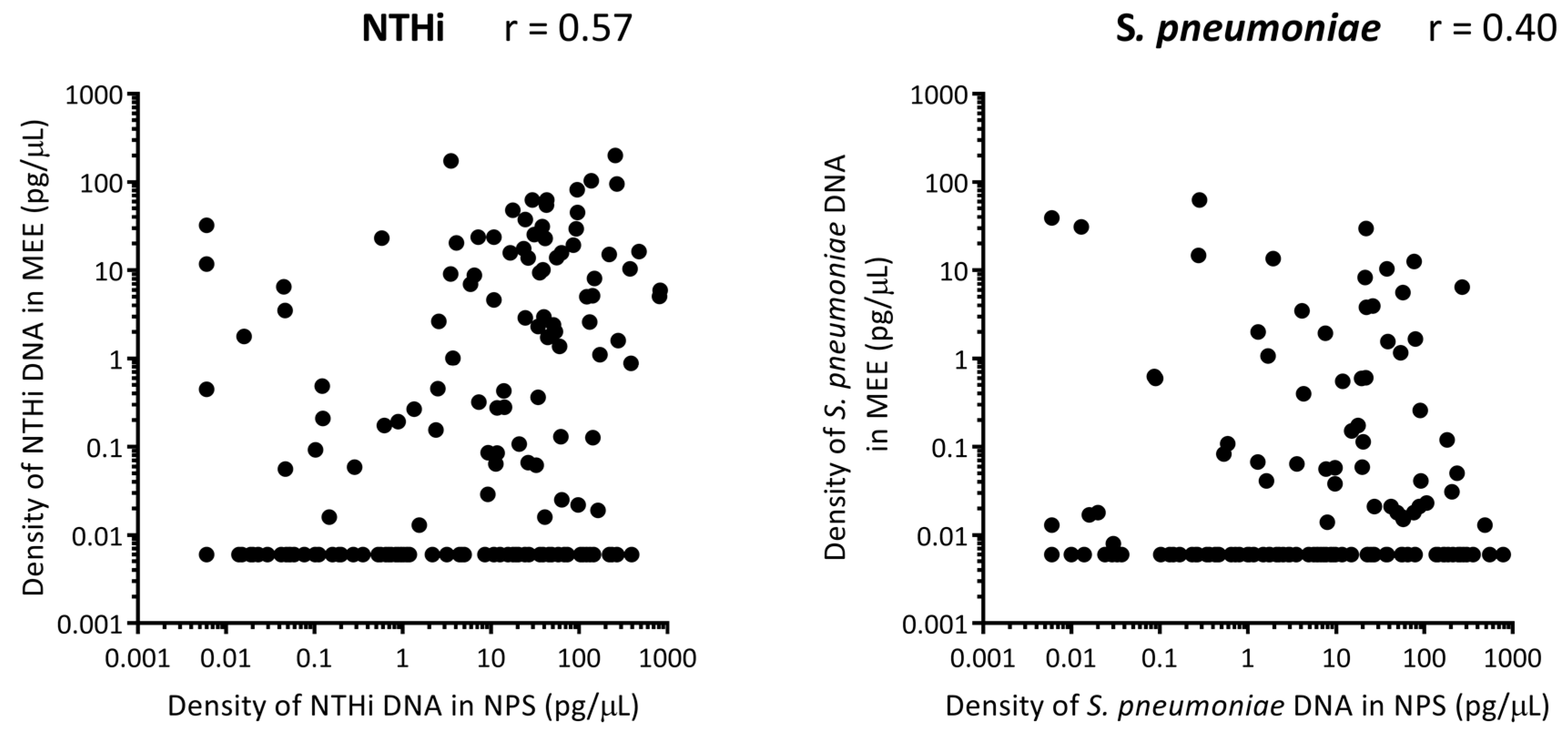
3.7. Correlation between Otopathogen Density in the Nasopharynx and Middle Ear
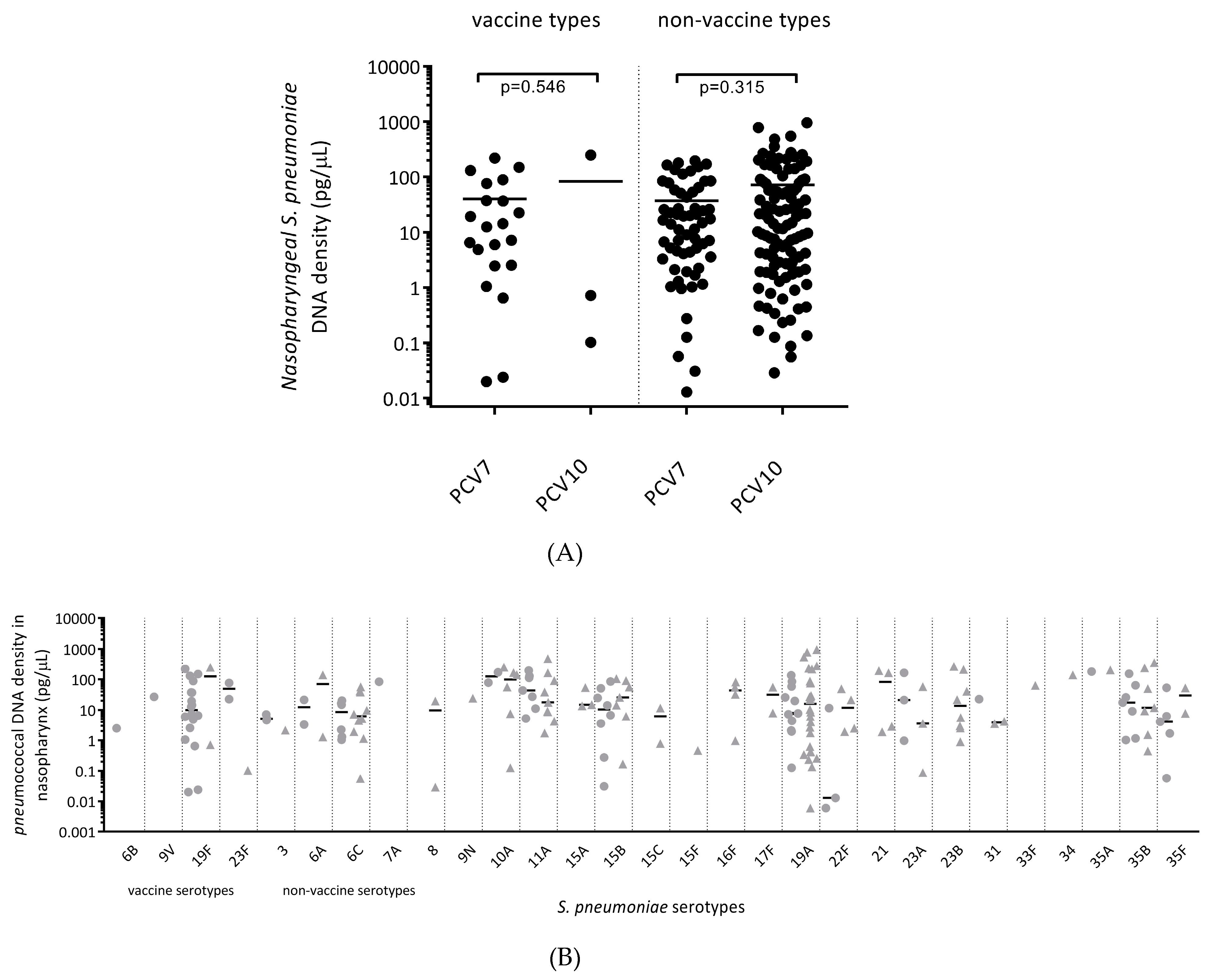
3.8. Carriage Density of Pneumococcal Vaccine and Non-Vaccine Serotypes was Similar between All PCV7- and All PCV10-Vaccinated Children
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grijalva, C.G.; Nuorti, J.P.; Griffin, M.R. Antibiotic prescription rates for acute respiratory tract infections in US ambulatory settings. JAMA 2009, 302, 758–766. [Google Scholar] [CrossRef] [PubMed]
- McCaig, L.F.; Besser, R.E.; Hughes, J.M. Trends in antimicrobial prescribing rates for children and adolescents. JAMA 2002, 287, 3096–3102. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.O. The burden of otitis media. Vaccine 2000, 19 (Suppl. S1), S2–S8. [Google Scholar] [CrossRef]
- Taylor, P.S.; Faeth, I.; Marks, M.K.; Del Mar, C.B.; Skull, S.A.; Pezzullo, M.L.; Havyatt, S.M.; Coates, H.L. Cost of treating otitis media in Australia. Expert Rev. Pharmacoecon. Outcomes Res. 2009, 9, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Barber, C.; Ille, S.; Vergison, A.; Coates, H. Acute otitis media in young children–what do parents say? Int. J. Pediatr. Otorhinolaryngol. 2014, 78, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Faden, H.; Duffy, L.; Wasielewski, R.; Wolf, J.; Krystofik, D.; Tung, Y. Relationship between nasopharyngeal colonization and the development of otitis media in children. Tonawanda/Williamsville Pediatrics. J. Infect. Dis. 1997, 175, 1440–1445. [Google Scholar] [CrossRef] [PubMed]
- Faden, H.; Stanievich, J.; Brodsky, L.; Bernstein, J.; Ogra, P.L. Changes in nasopharyngeal flora during otitis media of childhood. Pediatr. Infect. Dis. J. 1990, 9, 623–626. [Google Scholar] [PubMed]
- Schwartz, R.; Rodriguez, W.J.; Mann, R.; Khan, W.; Ross, S. The nasopharyngeal culture in acute otitis media. A reappraisal of its usefulness. JAMA 1979, 241, 2170–2173. [Google Scholar] [CrossRef]
- Giebink, G.S.; Berzins, I.K.; Marker, S.C.; Schiffman, G. Experimental otitis media after nasal inoculation of Streptococcus pneumoniae and influenza A virus in chinchillas. Infect. Immun. 1980, 30, 445–450. [Google Scholar] [PubMed]
- Eriksson, P.O.; Hellstrom, S. Acute otitis media develops in the rat after intranasal challenge of Streptococcus pneumoniae. Laryngoscope 2003, 113, 2047–2051. [Google Scholar] [CrossRef] [PubMed]
- Stol, K.; van Selm, S.; van den Berg, S.; Bootsma, H.J.; Blokx, W.A.; Graamans, K.; Tonnaer, E.L.; Hermans, P.W. Development of a non-invasive murine infection model for acute otitis media. Microbiology 2009, 155, 4135–4144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichichero, M.; Kaur, R.; Scott, D.A.; Gruber, W.C.; Trammel, J.; Almudevar, A.; Center, K.J. Effectiveness of 13-valent pneumococcal conjugate vaccination for protection against acute otitis media caused by Streptococcus pneumoniae in healthy young children: A prospective observational study. Lancet Child Adolesc. Health 2018, 2, 561–568. [Google Scholar] [CrossRef]
- Jardine, A.; Menzies, R.I.; Deeks, S.L.; Patel, M.S.; McIntyre, P.B. The impact of pneumococcal conjugate vaccine on rates of myringotomy with ventilation tube insertion in Australia. Pediatr. Infect. Dis. J. 2009, 28, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Poehling, K.A.; Szilagyi, P.G.; Grijalva, C.G.; Martin, S.W.; LaFleur, B.; Mitchel, E.; Barth, R.D.; Nuorti, J.P.; Griffin, M.R. Reduction of frequent otitis media and pressure-equalizing tube insertions in children after introduction of pneumococcal conjugate vaccine. Pediatrics 2007, 119, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Eskola, J.; Kilpi, T.; Palmu, A.; Jokinen, J.; Haapakoski, J.; Herva, E.; Takala, A.; Kayhty, H.; Karma, P.; Kohberger, R.; et al. Efficacy of a pneumococcal conjugate vaccine against acute otitis media. N. Engl. J. Med. 2001, 344, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Rovers, M.M.; Schilder, A.G.; Zielhuis, G.A.; Rosenfeld, R.M. Otitis media. Lancet 2004, 363, 465–473. [Google Scholar] [CrossRef]
- Roos, R.; Heffernan, H.; Quirke, T. Invasive Pneumococcal Disease in New Zealand, 2015; Institute of Environmental Science and Research Limited: Porirua, New Zealand, 2017.
- Vesikari, T.; Wysocki, J.; Chevallier, B.; Karvonen, A.; Czajka, H.; Arsene, J.P.; Lommel, P.; Dieussaert, I.; Schuerman, L. Immunogenicity of the 10-valent pneumococcal non-typeable Haemophilus influenzae protein D conjugate vaccine (PHiD-CV) compared to the licensed 7vCRM vaccine. Pediatr. Infect. Dis. J. 2009, 28, S66–S76. [Google Scholar] [CrossRef]
- Chevallier, B.; Vesikari, T.; Brzostek, J.; Knuf, M.; Bermal, N.; Aristegui, J.; Borys, D.; Cleerbout, J.; Lommel, P.; Schuerman, L. Safety and reactogenicity of the 10-valent pneumococcal non-typeable Haemophilus influenzae protein D conjugate vaccine (PHiD-CV) when coadministered with routine childhood vaccines. Pediatr. Infect. Dis. J. 2009, 28, S109–S118. [Google Scholar] [CrossRef]
- Leach, A.J.; Wigger, C.; Hare, K.; Hampton, V.; Beissbarth, J.; Andrews, R.; Chatfield, M.; Smith-Vaughan, H.; Morris, P.S. Reduced middle ear infection with non-typeable Haemophilus influenzae, but not Streptococcus pneumoniae, after transition to 10-valent pneumococcal non-typeable H. influenzae protein D conjugate vaccine. BMC Pediatr. 2015, 15, 162. [Google Scholar] [CrossRef]
- Van den Bergh, M.R.; Spijkerman, J.; Swinnen, K.M.; Francois, N.A.; Pascal, T.G.; Borys, D.; Schuerman, L.; Ijzerman, E.P.; Bruin, J.P.; van der Ende, A.; et al. Effects of the 10-valent pneumococcal nontypeable Haemophilus influenzae protein D-conjugate vaccine on nasopharyngeal bacterial colonization in young children: A randomized controlled trial. Clin. Infect. Dis. 2013, 56, e30–e39. [Google Scholar] [CrossRef]
- Vesikari, T.; Forsten, A.; Seppa, I.; Kaijalainen, T.; Puumalainen, T.; Soininen, A.; Traskine, M.; Lommel, P.; Schoonbroodt, S.; Hezareh, M.; et al. Effectiveness of the 10-Valent Pneumococcal Nontypeable Haemophilus influenzae Protein D-Conjugated Vaccine (PHiD-CV) Against Carriage and Acute Otitis Media-A Double-Blind Randomized Clinical Trial in Finland. J. Pediatr. Infect. Dis. Soc. 2016, 5, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Van Dongen, T.M.; van der Heijden, G.J.; van Zon, A.; Bogaert, D.; Sanders, E.A.; Schilder, A.G. Evaluation of concordance between the microorganisms detected in the nasopharynx and middle ear of children with otitis media. Pediatr. Infect. Dis. J. 2013, 32, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Smith-Vaughan, H.; Byun, R.; Nadkarni, M.; Jacques, N.A.; Hunter, N.; Halpin, S.; Morris, P.S.; Leach, A.J. Measuring nasal bacterial load and its association with otitis media. BMC Ear Nose Throat Disord. 2006, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Michel, L.V.; Kaur, R.; Zavorin, M.; Pryharski, K.; Khan, M.N.; LaClair, C.; O’Neil, M.; Xu, Q.; Pichichero, M.E. Intranasal coinfection model allows for assessment of protein vaccines against nontypeable Haemophilus influenzae in mice. J. Med. Microbiol. 2018, 67, 1527–1532. [Google Scholar] [CrossRef] [PubMed]
- Mills, N.; Best, E.J.; Murdoch, D.; Souter, M.; Neeff, M.; Anderson, T.; Salkeld, L.; Ahmad, Z.; Mahadevan, M.; Barber, C.; et al. What is behind the ear drum? The microbiology of otitis media and the nasopharyngeal flora in children in the era of pneumococcal vaccination. J. Paediatr. Child Health 2015, 51, 300–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Best, E.J.; Walls, T.; Souter, M.; Neeff, M.; Anderson, T.; Salkeld, L.; Ahmad, Z.; Mahadevan, M.; Walker, C.; Murdoch, D.; et al. Pneumococcal vaccine impact on otitis media microbiology: A New Zealand cohort study before and after the introduction of PHiD-CV10 vaccine. Vaccine 2016, 34, 3840–3847. [Google Scholar] [CrossRef] [PubMed]
- Wiertsema, S.P.; Kirkham, L.A.; Corscadden, K.J.; Mowe, E.N.; Bowman, J.M.; Jacoby, P.; Francis, R.; Vijayasekaran, S.; Coates, H.L.; Riley, T.V.; et al. Predominance of nontypeable Haemophilus influenzae in children with otitis media following introduction of a 3+0 pneumococcal conjugate vaccine schedule. Vaccine 2011, 29, 5163–5170. [Google Scholar] [CrossRef] [PubMed]
- Bakaletz, L.O.; Tallan, B.M.; Hoepf, T.; DeMaria, T.F.; Birck, H.G.; Lim, D.J. Frequency of fimbriation of nontypable Haemophilus influenzae and its ability to adhere to chinchilla and human respiratory epithelium. Infect. Immun. 1988, 56, 331–335. [Google Scholar] [Green Version]
- De Gier, C.; Pickering, J.L.; Richmond, P.C.; Thornton, R.B.; Kirkham, L.A. Duplex Quantitative PCR Assay for Detection of Haemophilus influenzae That Distinguishes Fucose- and Protein D-Negative Strains. J. Clin. Microbiol. 2016, 54, 2380–2383. [Google Scholar] [CrossRef]
- Best, E.J.; Mills, N.; Kate, G.; Hinds, J.; Anderson, T.; Murdoch, D.; Walls, T. Microarray detection of pneumococcal serotypes in middle ear fluid in children with established ear disease. In Proceedings of the 11th International Symposium on Pneumococci and Pneumococcal Diseases (ISPPD 2018), Melbourne, Australia, 15–19 April 2018. [Google Scholar]
- Robbins, J.B.; Austrian, R.; Lee, C.J.; Rastogi, S.C.; Schiffman, G.; Henrichsen, J.; Makela, P.H.; Broome, C.V.; Facklam, R.R.; Tiesjema, R.H.; et al. Considerations for formulating the second-generation pneumococcal capsular polysaccharide vaccine with emphasis on the cross-reactive types within groups. J. Infect. Dis. 1983, 148, 1136–1159. [Google Scholar] [CrossRef]
- Poolman, J.; Frasch, C.; Nurkka, A.; Kayhty, H.; Biemans, R.; Schuerman, L. Impact of the conjugation method on the immunogenicity of Streptococcus pneumoniae serotype 19F polysaccharide in conjugate vaccines. Clin. Vaccine Immunol. 2011, 18, 327–336. [Google Scholar] [CrossRef]
- Lewnard, J.A.; Givon-Lavi, N.; Tahtinen, P.A.; Dagan, R. Pneumococcal Phenotype and Interaction with Nontypeable Haemophilus influenzae as Determinants of Otitis Media Progression. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [PubMed]
- Petousis-Harris, H.; Howe, A.S.; Paynter, J.; Turner, N.; Griffin, J. Pneumococcal conjugate vaccines turning the tide on inequity—A retrospective cohort study of New Zealand children born 2006–2015. Clin. Infect. Dis. 2018. [Google Scholar] [CrossRef] [PubMed]
- Corscadden, K.J.; Kirkham, L.A.; Thornton, R.B.; Vijayasekaran, S.; Coates, H.L.; Richmond, P.C.; Wiertsema, S.P.; Vijayasekaran, S.; Coates, H.; Thornton, R.B.; et al. High pneumococcal serotype specific IgG, IgG1 and IgG2 levels in serum and the middle ear of children with recurrent acute otitis media receiving ventilation tubes. Vaccine 2013, 31, 1393–1399. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, L.S.; Wiertsema, S.P.; Corscadden, K.J.; Mateus, T.; Mullaney, G.L.; Zhang, G.; Richmond, P.C.; Thornton, R.B. Otitis-Prone Children Produce Functional Antibodies to Pneumolysin and Pneumococcal Polysaccharides. Clin. Vaccine Immunol. 2017, 24. [Google Scholar] [CrossRef]
- Menon, V.J.; Corscadden, K.J.; Fuery, A.; Thornton, R.B.; Kirkham, L.A.; Richmond, P.C.; Wiertsema, S.P. Children with otitis media mount a pneumococcal serotype specific serum IgG and IgA response comparable to healthy controls after pneumococcal conjugate vaccination. Vaccine 2012, 30, 3136–3144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornton, R.B.; Kirkham, L.S.; Corscadden, K.J.; Coates, H.L.; Vijayasekaran, S.; Hillwood, J.; Toster, S.; Edminston, P.; Zhang, G.; Keil, A.; et al. No evidence for impaired humoral immunity to pneumococcal proteins in Australian Aboriginal children with otitis media. Int. J. Pediatr. Otorhinolaryngol. 2017, 92, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Thornton, R.B.; Kirkham, L.S.; Corscadden, K.J.; Wiertsema, S.P.; Fuery, A.; Jones, B.J.; Coates, H.L.; Vijayasekaran, S.; Zhang, G.; Keil, A.; et al. Australian Aboriginal Children with Otitis Media Have Reduced Antibody Titers to Specific Nontypeable Haemophilus influenzae Vaccine Antigens. Clin. Vaccine Immunol. 2017, 24. [Google Scholar] [CrossRef]
- Brandileone, M.C.; Zanella, R.C.; Almeida, S.C.G.; Brandao, A.P.; Ribeiro, A.F.; Carvalhanas, T.M.P.; Sato, H.; Andrade, A.L.; Verani, J.R.; Pneumococcal Carriage Study, G. Effect of 10-valent pneumococcal conjugate vaccine on nasopharyngeal carriage of Streptococcus pneumoniae and Haemophilus influenzae among children in Sao Paulo, Brazil. Vaccine 2016, 34, 5604–5611. [Google Scholar] [CrossRef]
- Prymula, R.; Kriz, P.; Kaliskova, E.; Pascal, T.; Poolman, J.; Schuerman, L. Effect of vaccination with pneumococcal capsular polysaccharides conjugated to Haemophilus influenzae-derived protein D on nasopharyngeal carriage of Streptococcus pneumoniae and H. influenzae in children under 2 years of age. Vaccine 2009, 28, 71–78. [Google Scholar] [CrossRef]
- Baggett, H.C.; Watson, N.L.; Deloria Knoll, M.; Brooks, W.A.; Feikin, D.R.; Hammitt, L.L.; Howie, S.R.C.; Kotloff, K.L.; Levine, O.S.; Madhi, S.A.; et al. Density of Upper Respiratory Colonization With Streptococcus pneumoniae and Its Role in the Diagnosis of Pneumococcal Pneumonia Among Children Aged <5 Years in the PERCH Study. Clin. Infect. Dis. 2017, 64, S317–S327. [Google Scholar] [CrossRef]
- Bhuiyan, M.U.; Snelling, T.L.; West, R.; Lang, J.; Rahman, T.; Granland, C.; de Gier, C.; Borland, M.L.; Thornton, R.B.; Kirkham, L.S.; et al. The contribution of viruses and bacteria to community-acquired pneumonia in vaccinated children: A case-control study. Thorax 2018, 0, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bennett, K.E.; Haggard, M.P. Accumulation of factors influencing children’s middle ear disease: Risk factor modelling on a large population cohort. J. Epidemiol. Community Health 1998, 52, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Rovers, M.M.; Zielhuis, G.A.; Ingels, K.; van der Wilt, G.J. Day-care and otitis media in young children: A critical overview. Eur. J. Pediatr. 1999, 158, 1–6. [Google Scholar] [PubMed]
- Rovers, M.M.; de Kok, I.M.; Schilder, A.G. Risk factors for otitis media: An international perspective. Int. J. Pediatr. Otorhinolaryngol. 2006, 70, 1251–1256. [Google Scholar] [CrossRef] [PubMed]
- Education, N.Z.M.o. 20 Hours ECE (Early Childhood Education). Available online: https://parents.education.govt.nz/early-learning/early-childhood-education/20-hours-ece-2/ (accessed on 1 November 2018).
- Hare, K.M.; Smith-Vaughan, H.C.; Leach, A.J. Viability of respiratory pathogens cultured from nasopharyngeal swabs stored for up to 12 years at −70 degrees C in skim milk tryptone glucose glycerol broth. J. Microbiol. Methods 2011, 86, 364–367. [Google Scholar] [CrossRef] [PubMed]
- Kaijalainen, T.; Palmu, A. Long-term survival of Streptococcus pneumoniae, Haemophilus influenzae and Moraxella catarrhalis as isolates and in nasopharyngeal specimens in frozen STGG storage medium. J. Microbiol. Methods 2015, 114, 38–39. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Detected Species | Primer/Probe a | Sequence (5’ to 3’) | Concentration in Reaction Mix | Cycling Conditions | Ref. |
|---|---|---|---|---|---|---|
| fucP | H. influenzae | fucP fwd | GCCGCTTCTGAGGCTGG | 1000 nM | 50 °C for 2 min and 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 sec and 60 °C for 60 sec. | [26] |
| fucP rev | AACGACATTACCAATCCGATGG | 1000 nM | ||||
| fucP probe | 6FAM-TCCATTACTGTTTGAAATAC-MGBNFQ | 1000 nM | ||||
| hpd#3 | H. influenzae | hpd3 fwd | GGTTAAATATGCCGATGGTGTTG | 1000 nM | 50 °C for 2 min and 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 sec and 60 °C for 60 sec. | [27] |
| hpd3 rev | TGCATCTTTACGCACGGTGTA | 1000 nM | ||||
| hpd3 probe b | HEX-TTGTGTACACTCCGT/ZEN/TGGTAAAAGAACTTGCAC-3C6 | 1000 nM | ||||
| lytA | S. pneumoniae | lytA fwd | ACGCAATCTAGCAGATGAAGCA | 200 nM | 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 sec and 60 °C for 60 sec. | [28] |
| lytA rev | TCGTGCGTTTTAATTCCAGCT | 200 nM | ||||
| lytA probe | 6FAM-TGCCGAAAACGCTTGATACAG-GGAG-BHQ1 | 200 nM |
| PCV7 Vaccine Group | PCV10 Vaccine Group | |||||
|---|---|---|---|---|---|---|
| Sample Demographics | Cases | Controls | p | Cases | Controls | p |
| Total number | 147 | 70 | 178 | 62 | ||
| Median age in months (interquartile range) | 21.34 (16.54–26.63) | 18.82 (12.86–24.8) | 0.031 | 24.17 (19.05–28.81) | 21.11 (14.7–29.08) | 0.043 |
| Male gender (%) | 92 (63%) | 51 (73%) | 0.136 | 122 (69%) | 34 (55%) | 0.051 |
| Day care attendance (%) | 93 (63%) | 28 (40%) | 0.002 | 144 (81%) | 28 (45%) | 0.0001 |
| Antibiotics in last month (%) | 74 (50%) | 29 (41%) | 0.201 | 78 (44%) | 18 (29%) | 0.171 |
| Ethnicity (%) | ||||||
| European | 104 (71%) | 45 (64%) | 0.337 | 103 (58%) | 30 (48%) | 0.196 |
| Māori | 21 (14%) | 5 (7%) | 0.130 | 45 (25%) | 14 (23%) | 0.671 |
| Pacific Island | 19 (13%) | 13 (19%) | 0.273 | 27 (15%) | 9 (15%) | 0.901 |
| Other/unknown | 3 (2%) | 7 (10%) | 0.014 | 3 (2%) | 9 (15%) | 0.0001 |
| Otitis-Prone Children (Cases) | Non-Otitis-Prone (Controls) | |||||
|---|---|---|---|---|---|---|
| Sample Demographics | PCV7 | PCV10 | p | PCV7 | PCV10 | p |
| Total number | 147 | 178 | 70 | 62 | ||
| Median age in months (interquartile range) | 21.34 (16.54–26.63) | 24.17 (19.05–28.81) | 0.007 | 18.82 (12.86–24.8) | 21.11 (14.7–29.08) | 0.183 |
| Male gender (%) | 92 (63%) | 122 (69%) | 0.260 | 51 (73%) | 34 (55%) | 0.031 |
| Day care attendance (%) | 93 (63%) | 144 (81%) | 0.001 | 28 (40%) | 28 (45%) | 0.270 |
| Antibiotics in last month (%) | 74 (50%) | 78 (44%) | 0.176 | 29 (41%) | 18 (29%) | 0.325 |
| Ethnicity (%) | ||||||
| European | 104 (71%) | 103 (58%) | 0.016 | 45(64%) | 30 (48%) | 0.066 |
| Māori | 21 (14%) | 45 (25%) | 0.014 | 5 (7%) | 14 (23%) | 0.012 |
| Pacific Island | 19 (13%) | 27 (15%) | 0.564 | 13 (19%) | 9 (15%) | 0.533 |
| Other/unknown | 3 (2%) | 3 (2%) | 0.813 | 7 (10%) | 9 (15%) | 0.428 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Gier, C.; Granland, C.M.; Pickering, J.L.; Walls, T.; Bhuiyan, M.; Mills, N.; Richmond, P.C.; Best, E.J.; Thornton, R.B.; Kirkham, L.-A.S. PCV7- and PCV10-Vaccinated Otitis-Prone Children in New Zealand Have Similar Pneumococcal and Haemophilus influenzae Densities in Their Nasopharynx and Middle Ear. Vaccines 2019, 7, 14. https://doi.org/10.3390/vaccines7010014
de Gier C, Granland CM, Pickering JL, Walls T, Bhuiyan M, Mills N, Richmond PC, Best EJ, Thornton RB, Kirkham L-AS. PCV7- and PCV10-Vaccinated Otitis-Prone Children in New Zealand Have Similar Pneumococcal and Haemophilus influenzae Densities in Their Nasopharynx and Middle Ear. Vaccines. 2019; 7(1):14. https://doi.org/10.3390/vaccines7010014
Chicago/Turabian Stylede Gier, Camilla, Caitlyn M. Granland, Janessa L. Pickering, Tony Walls, Mejbah Bhuiyan, Nikki Mills, Peter C. Richmond, Emma J. Best, Ruth B. Thornton, and Lea-Ann S. Kirkham. 2019. "PCV7- and PCV10-Vaccinated Otitis-Prone Children in New Zealand Have Similar Pneumococcal and Haemophilus influenzae Densities in Their Nasopharynx and Middle Ear" Vaccines 7, no. 1: 14. https://doi.org/10.3390/vaccines7010014
APA Stylede Gier, C., Granland, C. M., Pickering, J. L., Walls, T., Bhuiyan, M., Mills, N., Richmond, P. C., Best, E. J., Thornton, R. B., & Kirkham, L. -A. S. (2019). PCV7- and PCV10-Vaccinated Otitis-Prone Children in New Zealand Have Similar Pneumococcal and Haemophilus influenzae Densities in Their Nasopharynx and Middle Ear. Vaccines, 7(1), 14. https://doi.org/10.3390/vaccines7010014