Replication of Hepatitis E Virus (HEV) in Primary Human-Derived Monocytes and Macrophages In Vitro

 , , , and
, , , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
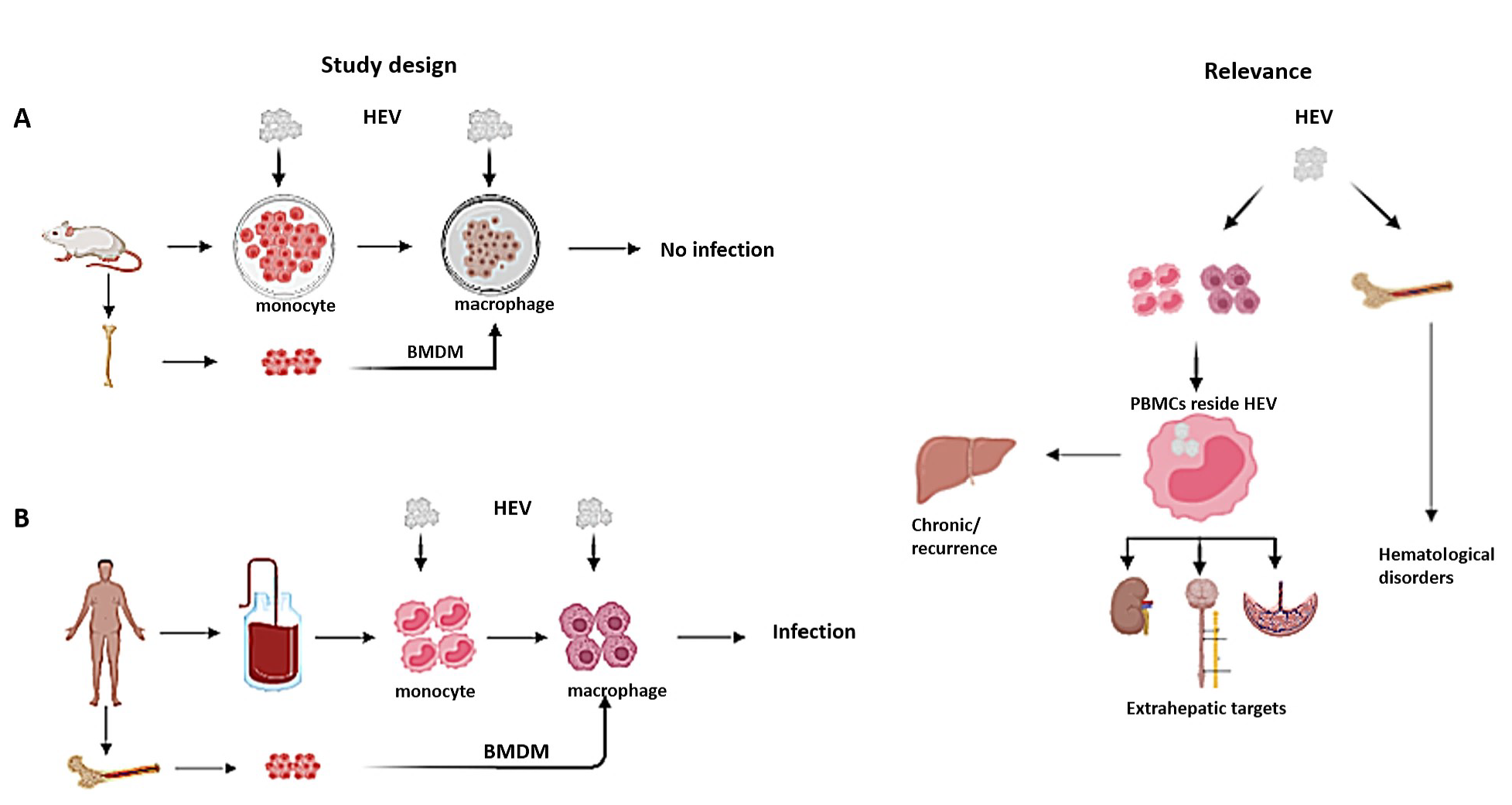
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Animals
2.3. Cell Lines
2.4. HEV Inoculums
2.5. Isolation of Mouse Monocytes and Bone Marrow and Differentiation into Macrophages In Vitro
2.6. Isolation of Human Monocytes and Differentiation into Macrophages In Vitro
2.7. Infection of HMOs and HMACs with HEV Preparations
2.8. Quantification of HEV RNA by qRT-PCR
2.9. Detection of HEV ORF2 Ag and ds-RNA in the Infected Cells
2.10. Monitoring of the Extracellular HEV Capsid Protein by ELISA
2.11. Measurement of the Level of Inflammatory Cytokines Released after HEV Infection
2.12. Statistics
3. Results
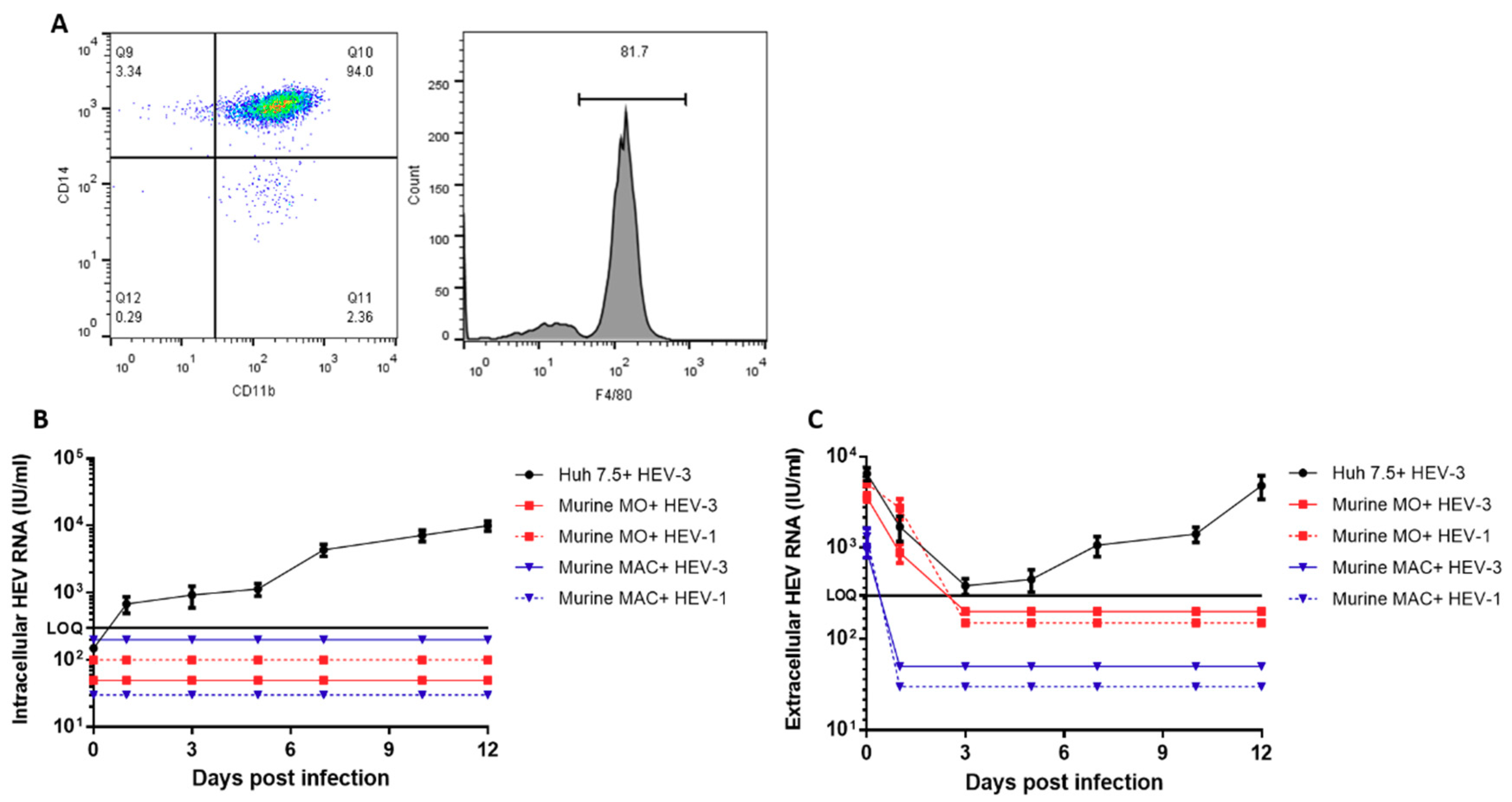
3.1. Murine Monocytes, Macrophages, and BMDMs are Not Susceptible to HEV Infection
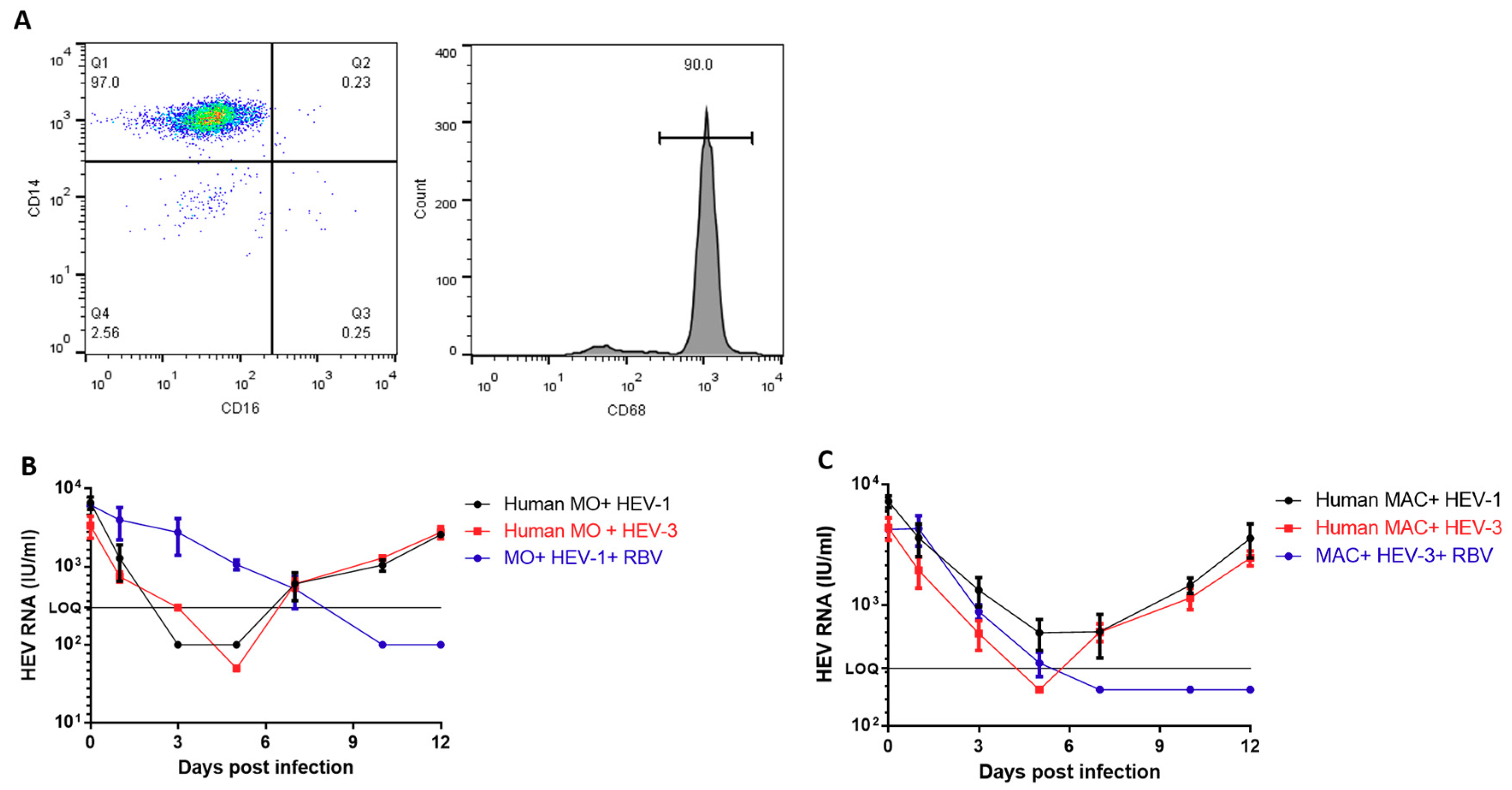
3.2. Human-Derived Monocytes and Macrophages are Susceptible to HEV Infection
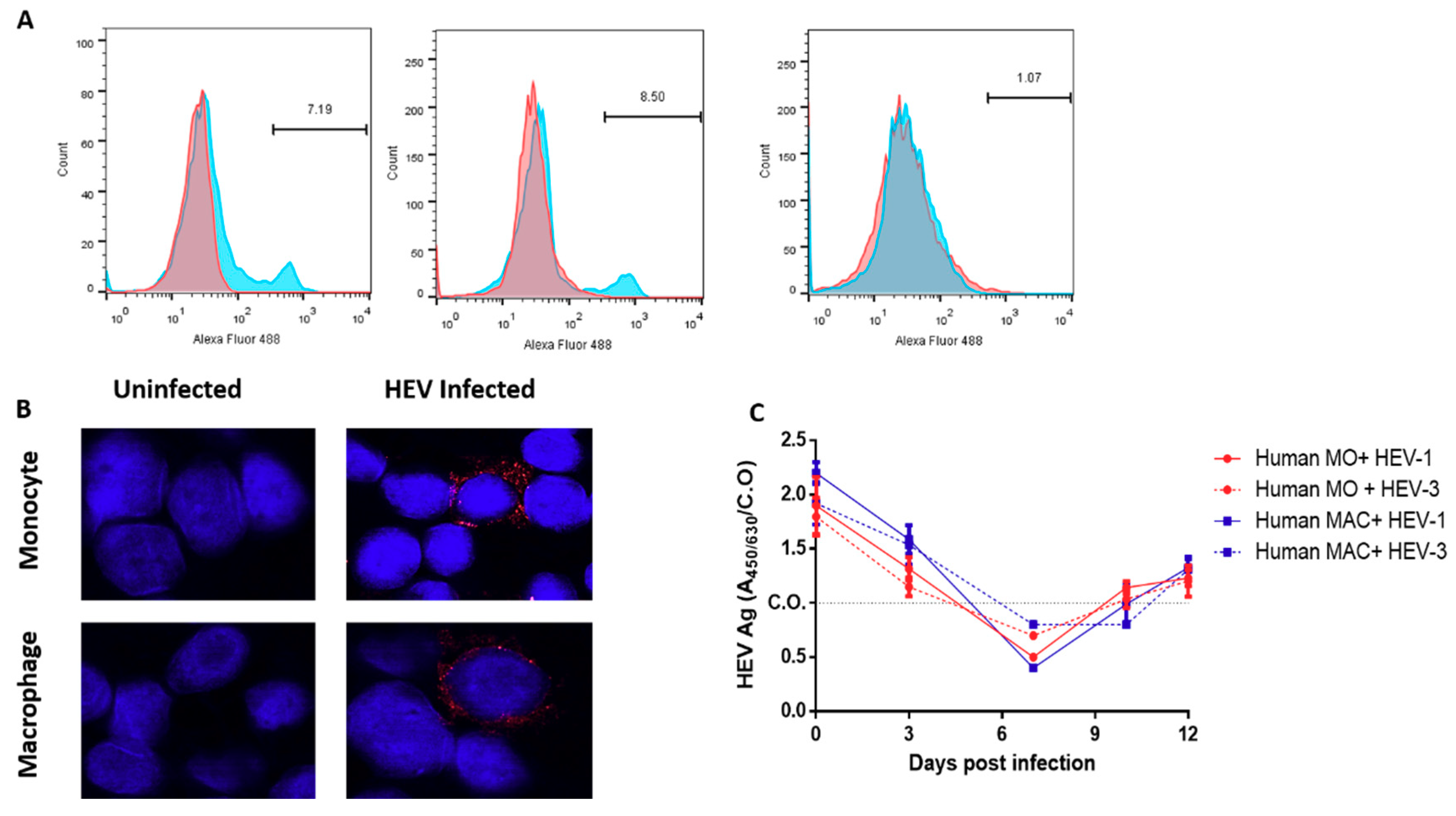
3.3. Detection of the HEV Capsid Protein and dsRNA in the Infected HMOs and HMACs
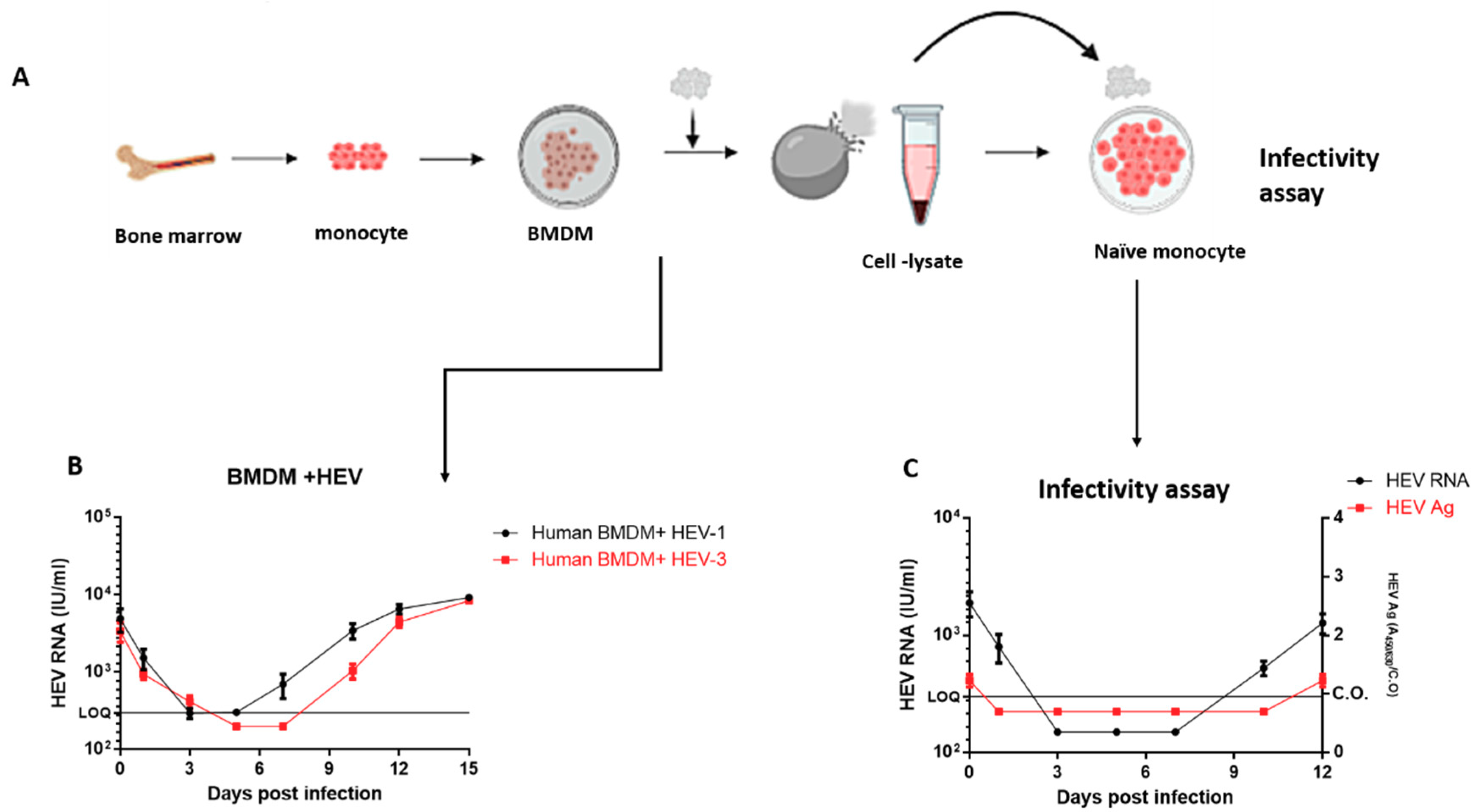
3.4. Replication of HEV in Human Bone Marrow-Derived Macrophages (BMDMs)
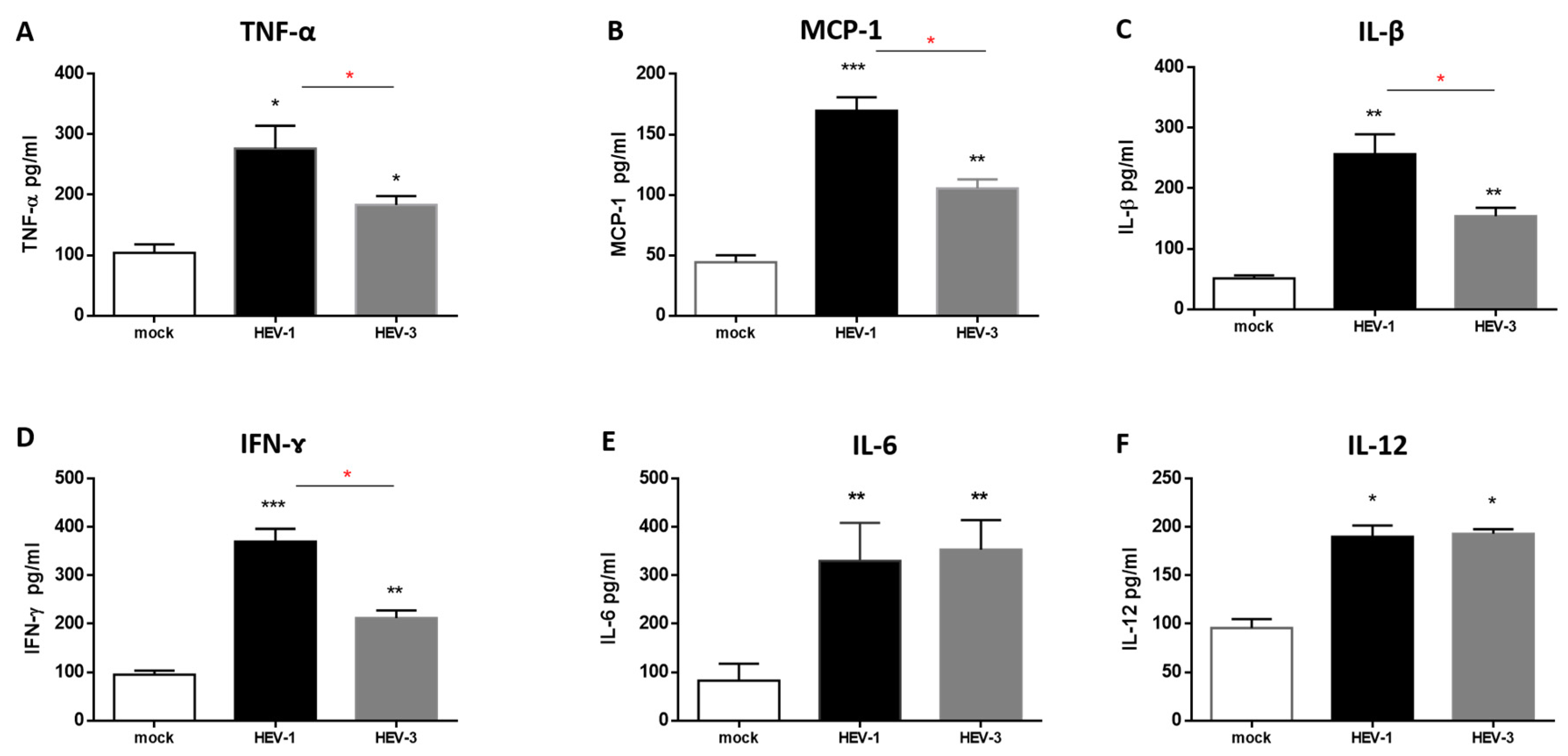
3.5. The Level of Proinflammatory Cytokines Was Elevated in the Supernatant of HEV-Infected HMACs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sayed, I.M.; Vercauteren, K.; Abdelwahab, S.F.; Meuleman, P. The emergence of hepatitis E virus in Europe. Futur. Virol. 2015, 10, 763–778. [Google Scholar] [CrossRef] [Green Version]
- Sayed, I.M.; Vercouter, A.S.; Abdelwahab, S.F.; Vercauteren, K.; Meuleman, P. Is hepatitis E virus an emerging problem in industrialized countries? Hepatology 2015, 62, 1883–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rein, D.B.; Stevens, G.A.; Theaker, J.; Wittenborn, J.S.; Wiersma, S.T. The global burden of hepatitis E virus genotypes 1 and 2 in 2005. Hepatology 2012, 55, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.-J.; Purcell, R.H.; Halbur, P.G.; Lehman, J.R.; Webb, D.M.; Tsareva, T.S.; Haynes, J.S.; Thacker, B.J.; Emerson, S.U. A novel virus in swine is closely related to the human hepatitis E virus. Proc. Natl. Acad. Sci. USA 1997, 94, 9860–9865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavio, N.; Meng, X.-J.; Doceul, V. Zoonotic origin of hepatitis E. Curr. Opin. Virol. 2015, 10, 34–41. [Google Scholar] [CrossRef]
- Kenney, S.P. The Current Host Range of Hepatitis E Viruses. Viruses 2019, 11, 452. [Google Scholar] [CrossRef] [Green Version]
- Domanović, D.; Tedder, R.; Blümel, J.; Zaaijer, H.; Gallian, P.; Niederhauser, C.; Oliveras, S.S.; O’Riordan, J.; Boland, F.; Harritshøj, L.; et al. Hepatitis E and blood donation safety in selected European countries: A shift to screening? Eurosurveillance 2017, 22, 22. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.; Simmonds, P.; Jameel, S.; Emerson, S.U.; Harrison, T.J.; Meng, X.-J.; Okamoto, H.; Van Der Poel, W.H.M.; Purdy, M.A.; members of the International Committee on the Taxonomy of Viruses Hepeviridae Study Group; et al. Consensus proposals for classification of the family Hepeviridae. J. Gen. Virol. 2014, 95, 2223–2232. [Google Scholar] [CrossRef]
- Takahashi, M.; Nishizawa, T.; Sato, H.; Sato, Y.; Jirintai; Nagashima, S.; Okamoto, H. Analysis of the full-length genome of a hepatitis E virus isolate obtained from a wild boar in Japan that is classifiable into a novel genotype. J. Gen. Virol. 2011, 92, 902–908. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.P.; Teng, J.L.; Tsang, A.K.L.; Joseph, M.; Wong, E.Y.; Tang, Y.; Sivakumar, S.; Xie, J.; Bai, R.; et al. New Hepatitis E Virus Genotype in Camels, the Middle East. Emerg. Infect. Dis. 2014, 20, 1044–1048. [Google Scholar] [CrossRef]
- Lee, G.-H.; Tan, B.-H.; Teo, E.C.-Y.; Lim, S.-G.; Dan, Y.-Y.; Wee, A.; Aw, P.P.K.; Zhu, Y.O.; Hibberd, M.L.; Tan, C.-K.; et al. Chronic Infection With Camelid Hepatitis E Virus in a Liver Transplant Recipient Who Regularly Consumes Camel Meat and Milk. Gastroenterology 2016, 150, 355–357.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenney, S.P.; Meng, X.-J. Hepatitis E Virus Genome Structure and Replication Strategy. Cold Spring Harb. Perspect. Med. 2018, 9, a031724. [Google Scholar]
- Dalton, H.; Kamar, N.; Baylis, S.A.; Moradpour, D.; Wedemeyer, H.; Negro, F. EASL Clinical Practice Guidelines on hepatitis E virus infection. J. Hepatol. 2018, 68, 1256–1271. [Google Scholar] [CrossRef] [PubMed]
- Dalton, H.; Bendall, R.P.; Keane, F.E.; Tedder, R.S.; Ijaz, S. Persistent Carriage of Hepatitis E Virus in Patients with HIV Infection. N. Engl. J. Med. 2009, 361, 1025–1027. [Google Scholar] [CrossRef]
- Kamar, N.; Selves, J.; Mansuy, J.-M.; Ouezzani, L.; Peron, J.-M.R.; Guitard, J.; Cointault, O.; Esposito, L.; Abravanel, F.; Danjoux, M.; et al. Hepatitis E Virus and Chronic Hepatitis in Organ-Transplant Recipients. N. Engl. J. Med. 2008, 358, 811–817. [Google Scholar] [CrossRef] [Green Version]
- Pischke, S.; Behrendt, P.; Manns, M.P.; Wedemeyer, H. HEV-associated cryoglobulinaemia and extrahepatic manifestations of hepatitis E. Lancet Infect. Dis. 2014, 14, 678–679. [Google Scholar]
- Pischke, S.; Hartl, J.; Pas, S.; Lohse, A.W.; Jacobs, B.C.; Van Der Eijk, A.A. Hepatitis E virus: Infection beyond the liver? J. Hepatol. 2017, 66, 1082–1095. [Google Scholar] [CrossRef] [Green Version]
- El-Mokhtar, M.A.; Othman, E.; Khashbah, M.; Ismael, A.; Ghaliony, M.; Seddik, M.; Sayed, I.M. Evidence of the Extrahepatic Replication of Hepatitis E Virus in Human Endometrial Stromal Cells. Pathogens 2020, 9, 295. [Google Scholar] [CrossRef] [Green Version]
- Wünschmann, S.; Becker, B.; Vallbracht, A. Hepatitis A Virus Suppresses Monocyte-to-Macrophage Maturation In Vitro. J. Virol. 2002, 76, 4350–4356. [Google Scholar] [CrossRef] [Green Version]
- Laskus, T.; Radkowski, M.; Wilkinson, J.; Vargas, H.E.; Rakela, J. The Origin of Hepatitis C Virus Reinfecting Transplanted Livers: Serum-Derived versus Peripheral Blood Mononuclear Cell–Derived Virus. J. Infect. Dis. 2002, 185, 417–421. [Google Scholar]
- Müller, H.M.; Pfaff, E.; Goeser, T.; Kallinowski, B.; Solbach, C.; Theilmann, L. Peripheral blood leukocytes serve as a possible extrahepatic site for hepatitis C virus replication. J. Gen. Virol. 1993, 74, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Radkowski, M.; Kubicka, J.; Kisiel, E.; Cianciara, J.; Nowicki, M.; Rakela, J.; Laskus, T. Detection of active hepatitis C virus and hepatitis G virus/GB virus C replication in bone marrow in human subjects. Blood 2000, 95, 3986–3989. [Google Scholar] [PubMed]
- Coluccio, C.; Begini, P.; Marzano, A.; Pellicelli, A.; Imperatrice, B.; Anania, G.; Fave, G.D.; Marignani, M. Hepatitis B in patients with hematological diseases: An update. World J. Hepatol. 2017, 9, 1043–1053. [Google Scholar] [CrossRef] [PubMed]
- Bazerbachi, F.; Haffar, S.; Garg, S.K.; Lake, J.R. Extra-hepatic manifestations associated with hepatitis E virus infection: A comprehensive review of the literature. Gastroenterol. Rep. 2015, 4, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Ippagunta, S.K.; Naik, S.; Jameel, S.; Ramana, K.N.S.; Aggarwal, R.; Kn, S.R. Viral RNA but no evidence of replication can be detected in the peripheral blood mononuclear cells of hepatitis E virus-infected patients. J. Viral Hepat. 2010, 18, 668–672. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.E.; El-Badawy, O.; Afifi, N.A.; Eldin, A.S.; Hassan, E.A.; Halby, H.M.; El-Mokhtar, M.A. Role of T-Helper 9 Cells in Chronic Hepatitis C-Infected Patients. Viruses 2018, 10, 341. [Google Scholar] [CrossRef] [Green Version]
- El-Mokhtar, M.A.; Elgendy, S.; El-Rehim, A.S.E.-D.A.; Hassan, E.A.; Hasan, A.A.A.; Hameed, M.A.; Sayed, D.; Salama, E. Hepatitis C Virus Affects Tuberculosis-Specific T Cells in HIV-Negative Patients. Viruses 2020, 12, 101. [Google Scholar] [CrossRef] [Green Version]
- Franz, S.; Hoeve, M.A.; Wickert, S.; Janko, C.; Dransfield, I. Clearance of apo Nph induces an immunosuppressive response in pro-inflammatory type-1 and anti-inflammatory type-2 MPhi. Autoimmunity 2009, 42, 275–277. [Google Scholar]
- Behrendt, P.; Drave, S.; Debing, Y.; Todt, D.; Friesland, M.; Engelmann, M.; Walter, S.; Wedemeyer, H.; Neyts, J.; Steinmann, E. Extra-Hepatic Replication and Infection of Hepatitis E Virus in Neuronal-Derived Cells. J. Hepatol. 2016, 64, S375. [Google Scholar] [CrossRef]
- Gouilly, J.; Chen, Q.; Siewiera, J.; Cartron, G.; Levy, C.; Dubois, M.; Al-Daccak, R.; Izopet, J.; Jabrane-Ferrat, N.; El Costa, H. Genotype specific pathogenicity of hepatitis E virus at the human maternal-fetal interface. Nat. Commun. 2018, 9, 4748. [Google Scholar]
- Knegendorf, L.; Drave, S.A.; Thi, V.L.D.; Debing, Y.; Brown, R.J.P.; Vondran, F.; Resner, K.; Friesland, M.; Khera, T.; Engelmann, M.; et al. Hepatitis E virus replication and interferon responses in human placental cells. Hepatol. Commun. 2018, 2, 173–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayed, I.M.; Verhoye, L.; Cocquerel, L.; Abravanel, F.; Foquet, L.; Montpellier, C.; Debing, Y.; Farhoudi, A.; Wychowski, C.; Dubuisson, J.D.; et al. Study of hepatitis E virus infection of genotype 1 and 3 in mice with humanised liver. Gut 2016, 66, 920–929. [Google Scholar] [CrossRef] [PubMed]
- Sayed, I.M.; Foquet, L.; Verhoye, L.; Abravanel, F.; Farhoudi, A.; Leroux-Roels, G.; Izopet, J.; Meuleman, P. Transmission of hepatitis E virus infection to human-liver chimeric FRG mice using patient plasma. Antivir. Res. 2017, 141, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Sayed, I.M.; Meuleman, P. Murine Tissues of Human Liver Chimeric Mice Are Not Susceptible to Hepatitis E Virus Genotypes 1 and 3. J. Infect. Dis. 2017, 216, 919–920. [Google Scholar] [CrossRef]
- Sayed, I.M.; Elkhawaga, A.A.; El-Mokhtar, M.A. Circulation of hepatitis E virus (HEV) and/or HEV-like agent in non-mixed dairy farms could represent a potential source of infection for Egyptian people. Int. J. Food Microbiol. 2020, 317, 108479. [Google Scholar] [CrossRef]
- Sayed, I.M.; Verhoye, L.; Montpellier, C.; Abravanel, F.; Izopet, J.; Cocquerel, L.; Meuleman, P. Study of hepatitis E virus ORF2 antigen kinetics in human-liver chimeric mice and its impact on HEV diagnosis. J. Infect. Dis. 2019, 220, 811–819. [Google Scholar] [CrossRef]
- Dieterich, D.T.; Spivak, J.L. Hematologic disorders associated with hepatitis C virus infection and their management. Clin. Infect. Dis. 2003, 37, 533–541. [Google Scholar] [CrossRef]
- Gallian, P.; Pouchol, E.; Djoudi, R.; Lhomme, S.; Mouna, L.; Gross, S.; Bierling, P.; Assal, A.; Kamar, N.; Mallet, V.; et al. Transfusion-Transmitted Hepatitis E Virus Infection in France. Transfus. Med. Rev. 2019, 33, 146–153. [Google Scholar] [CrossRef]
- Hewitt, P.E.; Ijaz, S.; Brailsford, S.R.; Brett, R.; Dicks, S.; Haywood, B.; Kennedy, I.; Kitchen, A.; Patel, P.; Poh, J.; et al. Hepatitis E virus in blood components: A prevalence and transmission study in southeast England. Lancet 2014, 384, 1766–1773. [Google Scholar] [CrossRef] [Green Version]
- Sayed, I.M.; Elkhawaga, A.A.; El-Mokhtar, M.A. In vivo models for studying Hepatitis E virus infection; Updates and applications. Virus Res. 2019, 274, 197765. [Google Scholar] [CrossRef]
- Sayed, I.M.; Meuleman, P. Updates in Hepatitis E virus (HEV) field; lessons learned from human liver chimeric mice. Rev. Med Virol. 2019, 30, e2086. [Google Scholar] [CrossRef] [PubMed]
- Targett-Adams, P.; Boulant, S.; Mclauchlan, J. Visualization of Double-Stranded RNA in Cells Supporting Hepatitis C Virus RNA Replication. J. Virol. 2007, 82, 2182–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stollar, B.; Stollar, V. Immunofluorescent demonstration of double-stranded RNA in the cytoplasm of sindbis virus-infected cells. Virology 1970, 42, 276–280. [Google Scholar] [CrossRef]
- Lee, J.; Marshall, J.; Bowden, D. Characterization of Rubella Virus Replication Complexes Using Antibodies to Double-Stranded RNA. Virology 1994, 200, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Comar, C.E.; Goldstein, S.A.; Li, Y.; Yount, B.; Baric, R.S.; Weiss, S.R.; Perlman, S.; Hogue, B. Antagonism of dsRNA-Induced Innate Immune Pathways by NS4a and NS4b Accessory Proteins during MERS Coronavirus Infection. mBio 2019, 10, e00319-19. [Google Scholar]
- Uchida, L.; Espada-Murao, L.A.; Takamatsu, Y.; Okamoto, K.; Hayasaka, D.; Yu, F.; Nabeshima, T.; Buerano, C.C.; Morita, K. The dengue virus conceals double-stranded RNA in the intracellular membrane to escape from an interferon response. Sci. Rep. 2014, 4, 7395. [Google Scholar]
- Feng, Q.; Hato, S.V.; Langereis, M.; Zoll, J.; Virgen-Slane, R.; Peisley, A.; Hur, S.; Semler, B.L.; Van Rij, R.P.; Van Kuppeveld, F.J. MDA5 Detects the Double-Stranded RNA Replicative Form in Picornavirus-Infected Cells. Cell Rep. 2012, 2, 1187–1196. [Google Scholar]
- Debes, J.D.; Groothuismink, Z.M.; Doukas, M.; De Man, R.A.; Boonstra, A. Immune dissociation during acute hepatitis E infection. Int. J. Infect. Dis. 2019, 87, 39–42. [Google Scholar] [CrossRef] [Green Version]
- Purcell, R.H.; Engle, R.E.; Govindarajan, S.; Herbert, R.; Claire, M.S.; Elkins, W.R.; Cook, A.; Shaver, C.; Beauregard, M.; Swerczek, J.; et al. Pathobiology of hepatitis E: Lessons learned from primate models. Emerg. Microbes Infect. 2013, 2, e9. [Google Scholar] [CrossRef]
- Shukla, P.; Nguyen, H.T.; Torian, U.; Engle, R.E.; Faulk, K.; Dalton, H.; Bendall, R.P.; Keane, F.E.; Purcell, R.H.; Emerson, S.U. Cross-species infections of cultured cells by hepatitis E virus and discovery of an infectious virus–host recombinant. Proc. Natl. Acad. Sci. USA 2011, 108, 2438–2443. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.T.; Shukla, P.; Torian, U.; Faulk, K.; Emerson, S.U. Hepatitis E Virus Genotype 1 Infection of Swine Kidney Cells In Vitro Is Inhibited at Multiple Levels. J. Virol. 2013, 88, 868–877. [Google Scholar] [PubMed] [Green Version]
- Navaneethan, U.; Al Mohajer, M.; Shata, M.T.M. Hepatitis E and pregnancy: Understanding the pathogenesis. Liver Int. 2008, 28, 1190–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, A.W.; White, R.; Reed, E.; Short, M.; Zhang, Y.; Fuerst, T.R.; Lanford, R.E. In VitroPropagation and Production of Hepatitis E Virus fromin Vivo-Infected Primary Macaque Hepatocytes. Virology 1996, 215, 1–9. [Google Scholar] [PubMed] [Green Version]
- Tam, A.W.; White, R.; Yarbough, P.O.; Murphy, B.J.; McAtee, C.; Lanford, R.E.; Fuerst, T.R. In VitroInfection and Replication of Hepatitis E Virus in Primary Cynomolgus Macaque Hepatocytes. Virology 1997, 238, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Fu, R.M.; Decker, C.C.; Thi, V.L.D. Cell Culture Models for Hepatitis E Virus. Viruses 2019, 11, 608. [Google Scholar] [CrossRef] [Green Version]
- Williams, T.P.E.; Kasorndorkbua, C.; Halbur, P.G.; Haqshenas, G.; Guenette, D.K.; Toth, T.E.; Meng, X.-J. Evidence of Extrahepatic Sites of Replication of the Hepatitis E Virus in a Swine Model. J. Clin. Microbiol. 2001, 39, 3040–3046. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; An, J.; She, R.; Shi, R.; Hao, W.; Soomro, M.; Yuan, X.; Yang, J.; Wang, J. Detection of Genotype 4 Swine Hepatitis E Virus in Systemic Tissues in Cross-Species Infected Rabbits. PLoS ONE 2017, 12, e0171277. [Google Scholar]
- Saravanabalaji, S.; Tripathy, A.S.; Dhoot, R.R.; Chadha, M.S.; Kakrani, A.L.; Arankalle, V.A. Viral load, antibody titers and recombinant open reading frame 2 protein-induced TH1/TH2 cytokines and cellular immune responses in self-limiting and fulminant hepatitise. Intervirology 2009, 52, 78–85. [Google Scholar] [CrossRef]
- Kumar, A.; Devi, S.G.; Kar, P.; Agarwal, S.; Husain, S.A.; Gupta, R.K.; Sharma, S. Association of cytokines in hepatitis E with pregnancy outcome. Cytokine 2014, 65, 95–104. [Google Scholar] [CrossRef]
- Taherkhani, R.; Farshadpour, F.; Makvandi, M.; Memari, H.R.; Samarbafzadeh, A.R.; Sharifi, N.; Naeimi, B.; Tajbakhsh, S.; Akbarzadeh, S. Cytokine Profiles and Cell Proliferation Responses to Truncated ORF2 Protein in Iranian Patients Recovered from Hepatitis E Infection. J. Trop. Med. 2015, 2015, 523560. [Google Scholar]
- Srivastava, R.; Aggarwal, R.; Jameel, S.; Puri, P.; Gupta, V.K.; Ramesh, V.; Bhatia, S.; Naik, S. Cellular immune responses in acute hepatitis E virus infection to the viral open reading frame 2 protein. Viral Immunol. 2007, 20, 56–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayed, I.M. Hepatic Immune Response against Hepatitis E Virus Genotype 1 Infection among Animal Models. J. Infect. Dis. 2019, 220, 1082–1083. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sayed, I.M.; Seddik, M.I.; Gaber, M.A.; Saber, S.H.; Mandour, S.A.; El-Mokhtar, M.A. Replication of Hepatitis E Virus (HEV) in Primary Human-Derived Monocytes and Macrophages In Vitro. Vaccines 2020, 8, 239. https://doi.org/10.3390/vaccines8020239
Sayed IM, Seddik MI, Gaber MA, Saber SH, Mandour SA, El-Mokhtar MA. Replication of Hepatitis E Virus (HEV) in Primary Human-Derived Monocytes and Macrophages In Vitro. Vaccines. 2020; 8(2):239. https://doi.org/10.3390/vaccines8020239
Chicago/Turabian StyleSayed, Ibrahim M., Mohamed Ismail Seddik, Marwa A. Gaber, Saber H. Saber, Sahar A. Mandour, and Mohamed A. El-Mokhtar. 2020. "Replication of Hepatitis E Virus (HEV) in Primary Human-Derived Monocytes and Macrophages In Vitro" Vaccines 8, no. 2: 239. https://doi.org/10.3390/vaccines8020239
APA StyleSayed, I. M., Seddik, M. I., Gaber, M. A., Saber, S. H., Mandour, S. A., & El-Mokhtar, M. A. (2020). Replication of Hepatitis E Virus (HEV) in Primary Human-Derived Monocytes and Macrophages In Vitro. Vaccines, 8(2), 239. https://doi.org/10.3390/vaccines8020239