Innate Immune Response against Hepatitis C Virus: Targets for Vaccine Adjuvants


Abstract
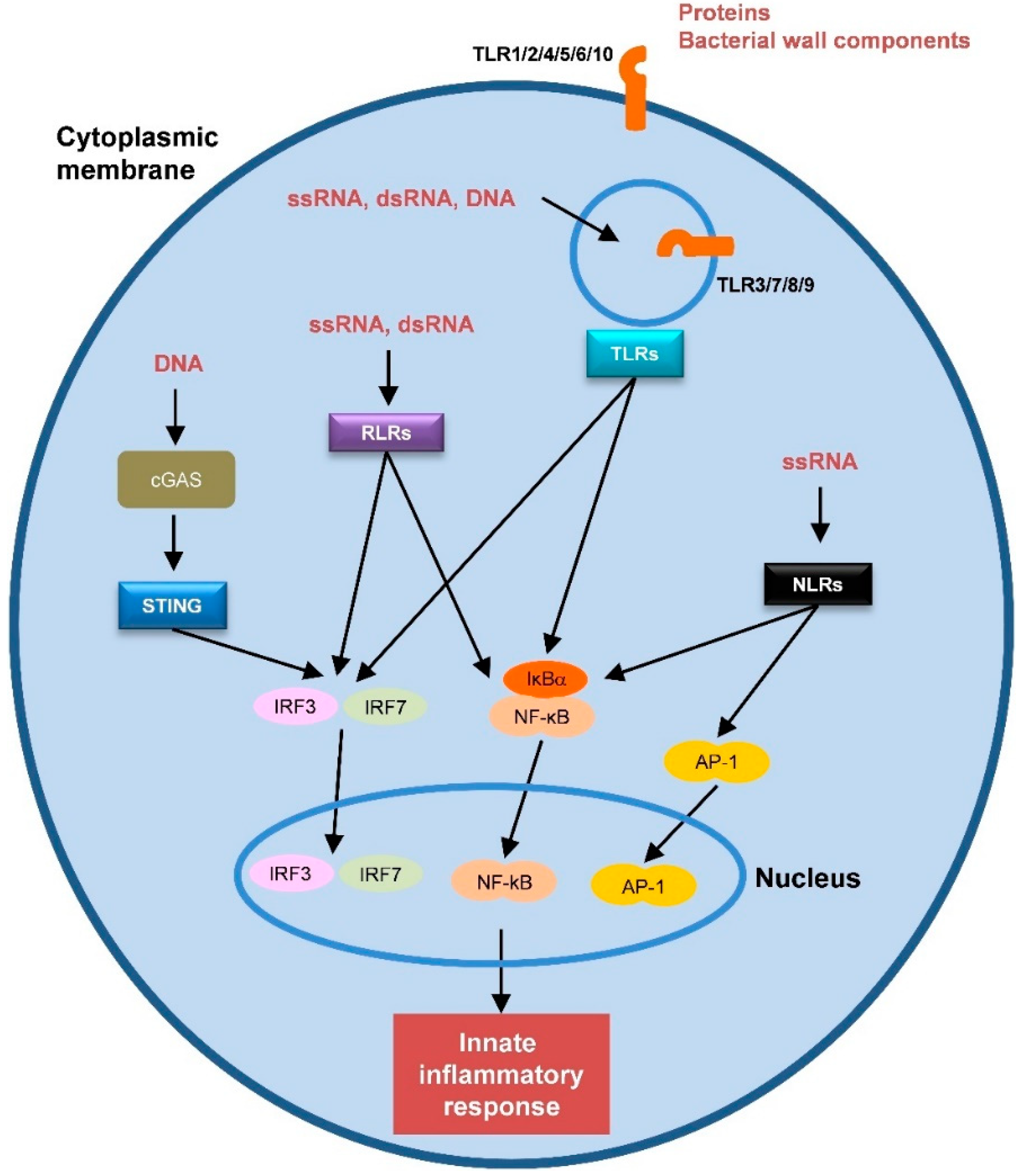
:1. Introduction
1.1. Hepatitis C Virus
1.2. Epidemiology of Hepatitis C
1.3. Antiviral Treatments
1.4. Immune Response and Vaccines
2. Hepatitis C Virus-Host Interaction: The Innate Immune Response
2.1. Hepatic Non-Immune Cells
2.1.1. Hepatocytes
2.1.2. Cholangiocytes
2.1.3. Hepatic Stellate Cells (HSCs)
2.1.4. Liver Sinusoidal Endothelial Cells (LSECs)
2.2. Hepatic Professional Immune Cells
2.2.1. Kupffer Cells (KCs)
2.2.2. Natural Killer (NK) Cells
2.2.3. Dendritic Cells (DCs)
3. Innate Immune Response and Vaccine Adjuvants
3.1. Innate Immunity Is Linked to Adaptive Immunity
3.2. Innate Immunity-Based Adjuvants
3.2.1. Toll-Like Receptors (TLRs)
3.2.2. Other Pattern Recognition Receptors (PRRs)
3.2.3. Combined Adjuvant Formulations: Adjuvant Systems
4. Adjuvants in HCV Vaccines
4.1. HCV Core and NS-Based Vaccines

{kind=link}
| Adjuvant Name | Vaccine Type | Main Stimulated Immune Responses | Trial Stage | Ref |
|---|---|---|---|---|
| hIL-12 | rNS3/core | N/A | Phase 1 completed (NCT03119025) | - |
| IFNL3 | DNA plasmid GLS-6150 (NS3/NS4A, NS4B, NS5A) | T-cell response | C57BL/6 mice Phase 1 completed (NCT02027116) | [150,151] |
| IFNL3 | DNA plasmid GLS-6150 (NS3/NS4A, NS4B, NS5A) | N/A | Phase 1 completed (NCT03674125) | - |
| hIL-12 | DNA plasmid INO-8000 (NS3/NS4A, NS4B, NS5A) | N/A | Phase 1 ongoing (NCT02772003) | - |
| Truncated form of Listeriolysin O | DNA plasmid NS3 | Ab response, T-cell response, Th1 type immunity | BALB/c mice | [155] |
| Hsp27, HR9 and Cady-2 | DNA plasmid hsp27-NS3 + HR9/Protein rhsp27-rNS3 +Cady-2 | Ab response, Th1 type immunity | BALB/c mice | [156] |
| Hp91 or hsp20 | DNA plasmid NS3 | Ab response, Th1 type immunity | BALB/c mice | [157] |
| Lenalidomide | DNA plasmid NS3 | T-cell response, Innate response | BALB/c mice | [158] |
| Stork HBcAg | DNA plasmid NS3/4A | Memory T-cell response | C57BL/6J mice | [159] |
| PADRE, lipopeptide from Neisseria meningiditis and IL-2 | rNS3, rNS4A/B, rNS5A | T-cell response, Th1 type immunity | BALB/c mice | [160] |
| G2 dendrimer | rNS3 | Ab response, T-cell response | BALB/c mice | [161] |
| Multihydroxylated fullerene (C60(OH)22) | rNS3/core | Ab response, T-cell response | BALB/c mice | [162] |
| Sodium polyprenyl phosphate | rNS3, rNS5B | Ab response, Th1 type immunity | DBA/2J mice | [163] |
| CAF09 | A mixture of 62 overlapping 20-mer peptides NS3 | T-cell response | CB6F1 mice | [164] |
| CAF09 | A mixture of six overlapping p7 peptides | Ab response, T-cell response | CB6F1, C57BL/6, and BALB/c mice | [165] |
| Neisseria meningitidis serogroup B outer membrane vesicles | rNS3/core | Ab response, Th1/Th2/Th17 type immunity | BALB/c mice | [166] |
| β-defensin | rNS3/4A, rNS5A, rNS5B | N/A | Immunoinformatics approaches | [167] |
4.2. HCV E1E2-Based Vaccines
| Adjuvant Name | Vaccine Type | Main Stimulated Immune Responses | Trial Stage | Ref |
|---|---|---|---|---|
| MF59 | rE1E2 | Ab response, CD4+ T-cell response | C57BL/6J mice Rhesus macaques Phase 1 completed | [174,175,176,177,178] |
| c-di-AMP STING or archaeosomes | rE1E2 | Ab response, CD4+ T-cell response, Th1 type immunity | CB6F1 mice | [179] |
| SLA (Enc) or (Adm)-based archaeosomes | rE1E2 | Ab response, CD4+ T-cell response, Th1 type immunity | C57Bl/6 x BALB/ c F1 mice | [180] |
| IMX313P | DNA plasmid expressing sE1/sE2 | Ab response | BALB/c mice | [184] |
| R4Pam2Cys or E8Pam2Cys | Quadrivalent genotype VLP vaccine (E1, E2) | Ab response, T-cell response | BALB/c mice | [185] |
| ODN39M | Chimeric NS3EnvCo + rE2 | Ab response, Th1 type immunity | BALB/c mice | [186] |
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Adams, R.L.; Pirakitikulr, N.; Pyle, A.M. Functional RNA structures throughout the Hepatitis C Virus genome. Curr. Opin. Virol. 2017, 24, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Bukh, J.; Kuiken, C.; Muerhoff, A.S.; Rice, C.M.; Stapleton, J.T.; Simmonds, P. Expanded classification of hepatitis C virus into 7 genotypes and 67 subtypes: Updated criteria and genotype assignment web resource. Hepatology 2014, 59, 318–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forns, X.; Purcell, R.H.; Bukh, J. Quasispecies in viral persistence and pathogenesis of hepatitis C virus. Trends Microbiol. 1999, 7, 402–410. [Google Scholar] [CrossRef]
- Farci, P.; Shimoda, A.; Coiana, A.; Diaz, G.; Peddis, G.; Melpolder, J.C.; Strazzera, A.; Chien, D.Y.; Munoz, S.J.; Balestrieri, A.; et al. The outcome of acute hepatitis C predicted by the evolution of the viral quasispecies. Science 2000, 288, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Lapa, D.; Garbuglia, A.R.; Capobianchi, M.R.; Del Porto, P. Hepatitis C Virus Genetic Variability, Human Immune Response, and Genome Polymorphisms: Which Is the Interplay? Cells 2019, 8, 305. [Google Scholar] [CrossRef] [Green Version]
- Spearman, C.W.; Dusheiko, G.M.; Hellard, M.; Sonderup, M. Hepatitis C. Lancet 2019, 394, 1451–1466. [Google Scholar] [CrossRef]
- Westbrook, R.H.; Dusheiko, G. Natural history of hepatitis C. J. Hepatol. 2014, 61, S58–S68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lingala, S.; Ghany, M.G. Natural History of Hepatitis C. Gastroenterol. Clin. N. Am. 2015, 44, 717–734. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. WHO Fact Sheets Hepatitis C. Available online: https://www.who.int/en/news-room/fact-sheets/detail/hepatitis-c (accessed on 8 July 2019).
- Polaris Observatory, H.C.V.C. Global prevalence and genotype distribution of hepatitis C virus infection in 2015: A modelling study. Lancet Gastroenterol Hepatol. 2017, 2, 161–176. [Google Scholar] [CrossRef] [Green Version]
- Ponziani, F.R.; Mangiola, F.; Binda, C.; Zocco, M.A.; Siciliano, M.; Grieco, A.; Rapaccini, G.L.; Pompili, M.; Gasbarrini, A. Future of liver disease in the era of direct acting antivirals for the treatment of hepatitis C. World J. Hepatol. 2017, 9, 352–367. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver. EASL Recommendations on Treatment of Hepatitis C 2016. J. Hepatol. 2017, 66, 153–194. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Draft Global Health Sector Strategies. Viral Hepatitis, 2016–2021; WHO: Geneva, Switzerland, 2016. [Google Scholar]
- Calvaruso, V.; Petta, S.; Craxi, A. Is global elimination of HCV realistic? Liver. Int. 2018, 38 (Suppl. 1), 40–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, A.L. MEDICINE. Global control of hepatitis C virus. Science 2015, 349, 790–791. [Google Scholar] [CrossRef] [PubMed]
- Roingeard, P.; Beaumont, E. Hepatitis C vaccine: 10 good reasons for continuing. Hepatology 2020. [Google Scholar] [CrossRef]
- Fine, P.; Eames, K.; Heymann, D.L. “Herd immunity”: A rough guide. Clin. Infect. Dis. 2011, 52, 911–916. [Google Scholar] [CrossRef]
- Major, M.; Gutfraind, A.; Shekhtman, L.; Cui, Q.; Kachko, A.; Cotler, S.J.; Hajarizadeh, B.; Sacks-Davis, R.; Page, K.; Boodram, B.; et al. Modeling of patient virus titers suggests that availability of a vaccine could reduce hepatitis C virus transmission among injecting drug users. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Hahn, J.A.; Wylie, D.; Dill, J.; Sanchez, M.S.; Lloyd-Smith, J.O.; Page-Shafer, K.; Getz, W.M. Potential impact of vaccination on the hepatitis C virus epidemic in injection drug users. Epidemics 2009, 1, 47–57. [Google Scholar] [CrossRef]
- Stone, J.; Martin, N.K.; Hickman, M.; Hellard, M.; Scott, N.; McBryde, E.; Drummer, H.; Vickerman, P. The Potential Impact of a Hepatitis C Vaccine for People Who Inject Drugs: Is a Vaccine Needed in the Age of Direct-Acting Antivirals? PLoS ONE 2016, 11, e0156213. [Google Scholar] [CrossRef] [Green Version]
- Read, A.F.; Baigent, S.J.; Powers, C.; Kgosana, L.B.; Blackwell, L.; Smith, L.P.; Kennedy, D.A.; Walkden-Brown, S.W.; Nair, V.K. Imperfect Vaccination Can Enhance the Transmission of Highly Virulent Pathogens. PLoS Biol. 2015, 13, e1002198. [Google Scholar] [CrossRef]
- Fauvelle, C.; Colpitts, C.C.; Keck, Z.Y.; Pierce, B.G.; Foung, S.K.; Baumert, T.F. Hepatitis C virus vaccine candidates inducing protective neutralizing antibodies. Expert. Rev. Vaccines 2016, 15, 1535–1544. [Google Scholar] [CrossRef]
- Saito, T.; Owen, D.M.; Jiang, F.; Marcotrigiano, J.; Gale, M., Jr. Innate immunity induced by composition-dependent RIG-I recognition of hepatitis C virus RNA. Nature 2008, 454, 523–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, X.; Ding, Q.; Lu, J.; Tao, W.; Huang, B.; Zhao, Y.; Niu, J.; Liu, Y.J.; Zhong, J. MDA5 plays a critical role in interferon response during hepatitis C virus infection. J. Hepatol. 2015, 62, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.; Gonzalez, V.D.; Li, Q.; Modi, A.A.; Chen, W.; Noureddin, M.; Rotman, Y.; Liang, T.J. HCV infection induces a unique hepatic innate immune response associated with robust production of type III interferons. Gastroenterology 2012, 142, 978–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Israelow, B.; Narbus, C.M.; Sourisseau, M.; Evans, M.J. HepG2 cells mount an effective antiviral interferon-lambda based innate immune response to hepatitis C virus infection. Hepatology 2014, 60, 1170–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickensheets, H.; Sheikh, F.; Park, O.; Gao, B.; Donnelly, R.P. Interferon-lambda (IFN-lambda) induces signal transduction and gene expression in human hepatocytes, but not in lymphocytes or monocytes. J. Leukoc. Biol. 2013, 93, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, N.F.; Humphreys, E.; Jennings, E.; Osburn, W.; Lissauer, S.; Wilson, G.K.; van, I.S.C.D.; Baumert, T.F.; Balfe, P.; Afford, S.; et al. Hepatitis C virus infection of cholangiocarcinoma cell lines. J. Gen. Virol. 2015, 96, 1380–1388. [Google Scholar] [CrossRef] [Green Version]
- Navas, M.C.; Glaser, S.; Dhruv, H.; Celinski, S.; Alpini, G.; Meng, F. Hepatitis C Virus Infection and Cholangiocarcinoma: An Insight into Epidemiologic Evidences and Hypothetical Mechanisms of Oncogenesis. Am. J. Pathol. 2019, 189, 1122–1132. [Google Scholar] [CrossRef] [Green Version]
- Syal, G.; Fausther, M.; Dranoff, J.A. Advances in cholangiocyte immunobiology. Am. J. Physiol. Gastrointest Liver. Physiol. 2012, 303, G1077–G1086. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, D.E. Immunopathogenesis of Hepatitis C Virus Infection. Gastroenterol. Clin. N. Am. 2015, 44, 735–760. [Google Scholar] [CrossRef] [Green Version]
- Seki, E.; Brenner, D.A. Toll-like receptors and adaptor molecules in liver disease: Update. Hepatology 2008, 48, 322–335. [Google Scholar] [CrossRef]
- Florimond, A.; Chouteau, P.; Bruscella, P.; Le Seyec, J.; Merour, E.; Ahnou, N.; Mallat, A.; Lotersztajn, S.; Pawlotsky, J.M. Human hepatic stellate cells are not permissive for hepatitis C virus entry and replication. Gut 2015, 64, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Trippler, M.; Pei, R.; Lu, M.; Broering, R.; Gerken, G.; Schlaak, J.F. Toll-like receptor activated human and murine hepatic stellate cells are potent regulators of hepatitis C virus replication. J. Hepatol. 2009, 51, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Lau, A.H.; Thomson, A.W. Dendritic cells and immune regulation in the liver. Gut 2003, 52, 307–314. [Google Scholar] [CrossRef] [Green Version]
- Giugliano, S.; Kriss, M.; Golden-Mason, L.; Dobrinskikh, E.; Stone, A.E.; Soto-Gutierrez, A.; Mitchell, A.; Khetani, S.R.; Yamane, D.; Stoddard, M.; et al. Hepatitis C virus infection induces autocrine interferon signaling by human liver endothelial cells and release of exosomes, which inhibits viral replication. Gastroenterology 2015, 148, 392–402.e313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Meng, Z.; Jiang, M.; Zhang, E.; Trippler, M.; Broering, R.; Bucchi, A.; Krux, F.; Dittmer, U.; Yang, D.; et al. Toll-like receptor-induced innate immune responses in non-parenchymal liver cells are cell type-specific. Immunology 2010, 129, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Kern, M.; Popov, A.; Scholz, K.; Schumak, B.; Djandji, D.; Limmer, A.; Eggle, D.; Sacher, T.; Zawatzky, R.; Holtappels, R.; et al. Virally infected mouse liver endothelial cells trigger CD8+ T-cell immunity. Gastroenterology 2010, 138, 336–346. [Google Scholar] [CrossRef]
- Gastaminza, P.; Dryden, K.A.; Boyd, B.; Wood, M.R.; Law, M.; Yeager, M.; Chisari, F.V. Ultrastructural and biophysical characterization of hepatitis C virus particles produced in cell culture. J. Virol. 2010, 84, 10999–11009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cormier, E.G.; Durso, R.J.; Tsamis, F.; Boussemart, L.; Manix, C.; Olson, W.C.; Gardner, J.P.; Dragic, T. L-SIGN (CD209L) and DC-SIGN (CD209) mediate transinfection of liver cells by hepatitis C virus. Proc. Natl. Acad. Sci. USA 2004, 101, 14067–14072. [Google Scholar] [CrossRef] [Green Version]
- Rowe, I.A.; Galsinh, S.K.; Wilson, G.K.; Parker, R.; Durant, S.; Lazar, C.; Branza-Nichita, N.; Bicknell, R.; Adams, D.H.; Balfe, P.; et al. Paracrine signals from liver sinusoidal endothelium regulate hepatitis C virus replication. Hepatology 2014, 59, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Crispe, I.N. Liver antigen-presenting cells. J Hepatol. 2011, 54, 357–365. [Google Scholar] [CrossRef]
- Schorey, J.S.; Cheng, Y.; Singh, P.P.; Smith, V.L. Exosomes and other extracellular vesicles in host-pathogen interactions. EMBO. Rep. 2015, 16, 24–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavin, Y.; Winter, D.; Blecher-Gonen, R.; David, E.; Keren-Shaul, H.; Merad, M.; Jung, S.; Amit, I. Tissue-resident macrophage enhancer landscapes are shaped by the local microenvironment. Cell 2014, 159, 1312–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, C.; Tacke, F. Hepatic macrophages in homeostasis and liver diseases: From pathogenesis to novel therapeutic strategies. Cell Mol. Immunol. 2016, 13, 316–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boltjes, A.; Movita, D.; Boonstra, A.; Woltman, A.M. The role of Kupffer cells in hepatitis B and hepatitis C virus infections. J. Hepatol. 2014, 61, 660–671. [Google Scholar] [CrossRef] [Green Version]
- Negash, A.A.; Ramos, H.J.; Crochet, N.; Lau, D.T.; Doehle, B.; Papic, N.; Delker, D.A.; Jo, J.; Bertoletti, A.; Hagedorn, C.H.; et al. IL-1beta production through the NLRP3 inflammasome by hepatic macrophages links hepatitis C virus infection with liver inflammation and disease. PLoS Pathog. 2013, 9, e1003330. [Google Scholar] [CrossRef]
- Shrivastava, S.; Mukherjee, A.; Ray, R.; Ray, R.B. Hepatitis C virus induces interleukin-1beta (IL-1beta)/IL-18 in circulatory and resident liver macrophages. J. Virol. 2013, 87, 12284–12290. [Google Scholar] [CrossRef] [Green Version]
- Tu, Z.; Pierce, R.H.; Kurtis, J.; Kuroki, Y.; Crispe, I.N.; Orloff, M.S. Hepatitis C virus core protein subverts the antiviral activities of human Kupffer cells. Gastroenterology 2010, 138, 305–314. [Google Scholar] [CrossRef]
- Hosomura, N.; Kono, H.; Tsuchiya, M.; Ishii, K.; Ogiku, M.; Matsuda, M.; Fujii, H. HCV-related proteins activate Kupffer cells isolated from human liver tissues. Dig. Dis. Sci. 2011, 56, 1057–1064. [Google Scholar] [CrossRef]
- Zhu, H.; Liu, C. Interleukin-1 inhibits hepatitis C virus subgenomic RNA replication by activation of extracellular regulated kinase pathway. J. Virol. 2003, 77, 5493–5498. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Shang, X.; Terada, N.; Liu, C. STAT3 induces anti-hepatitis C viral activity in liver cells. Biochem. Biophys. Res. Commun. 2004, 324, 518–528. [Google Scholar] [CrossRef]
- Sasaki, R.; Devhare, P.B.; Steele, R.; Ray, R.; Ray, R.B. Hepatitis C virus-induced CCL5 secretion from macrophages activates hepatic stellate cells. Hepatology 2017, 66, 746–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broering, R.; Wu, J.; Meng, Z.; Hilgard, P.; Lu, M.; Trippler, M.; Szczeponek, A.; Gerken, G.; Schlaak, J.F. Toll-like receptor-stimulated non-parenchymal liver cells can regulate hepatitis C virus replication. J. Hepatol. 2008, 48, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, N.F.; Sutaria, R.; Jo, J.; Barnes, A.; Blahova, M.; Meredith, L.W.; Cosset, F.L.; Curbishley, S.M.; Adams, D.H.; Bertoletti, A.; et al. Activated macrophages promote hepatitis C virus entry in a tumor necrosis factor-dependent manner. Hepatology 2014, 59, 1320–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simeonova, P.P.; Gallucci, R.M.; Hulderman, T.; Wilson, R.; Kommineni, C.; Rao, M.; Luster, M.I. The role of tumor necrosis factor-alpha in liver toxicity, inflammation, and fibrosis induced by carbon tetrachloride. Toxicol. Appl. Pharmacol. 2001, 177, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.P.; Zhou, L.; Wang, J.; Xiong, S.; Garner, W.L.; French, S.W.; Tsukamoto, H. Essential role of matrix metalloproteinases in interleukin-1-induced myofibroblastic activation of hepatic stellate cell in collagen. J. Biol. Chem. 2004, 279, 4820–4828. [Google Scholar] [CrossRef] [Green Version]
- Sandler, N.G.; Koh, C.; Roque, A.; Eccleston, J.L.; Siegel, R.B.; Demino, M.; Kleiner, D.E.; Deeks, S.G.; Liang, T.J.; Heller, T.; et al. Host response to translocated microbial products predicts outcomes of patients with HBV or HCV infection. Gastroenterology 2011, 141, 1220–1230. [Google Scholar] [CrossRef] [Green Version]
- Mengshol, J.A.; Golden-Mason, L.; Arikawa, T.; Smith, M.; Niki, T.; McWilliams, R.; Randall, J.A.; McMahan, R.; Zimmerman, M.A.; Rangachari, M.; et al. A crucial role for Kupffer cell-derived galectin-9 in regulation of T cell immunity in hepatitis C infection. PLoS ONE 2010, 5, e9504. [Google Scholar] [CrossRef]
- Harwood, N.M.; Golden-Mason, L.; Cheng, L.; Rosen, H.R.; Mengshol, J.A. HCV-infected cells and differentiation increase monocyte immunoregulatory galectin-9 production. J. Leukoc. Biol. 2016, 99, 495–503. [Google Scholar] [CrossRef] [Green Version]
- Nishio, A.; Tatsumi, T.; Nawa, T.; Suda, T.; Yoshioka, T.; Onishi, Y.; Aono, S.; Shigekawa, M.; Hikita, H.; Sakamori, R.; et al. CD14(+) monocyte-derived galectin-9 induces natural killer cell cytotoxicity in chronic hepatitis C. Hepatology 2017, 65, 18–31. [Google Scholar] [CrossRef] [Green Version]
- Nellore, A.; Fishman, J.A. NK cells, innate immunity and hepatitis C infection after liver transplantation. Clin. Infect. Dis. 2011, 52, 369–377. [Google Scholar] [CrossRef]
- Vivier, E.; Raulet, D.H.; Moretta, A.; Caligiuri, M.A.; Zitvogel, L.; Lanier, L.L.; Yokoyama, W.M.; Ugolini, S. Innate or adaptive immunity? The example of natural killer cells. Science 2011, 331, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Cooper, M.A.; Fehniger, T.A.; Turner, S.C.; Chen, K.S.; Ghaheri, B.A.; Ghayur, T.; Carson, W.E.; Caligiuri, M.A. Human natural killer cells: A unique innate immunoregulatory role for the CD56(bright) subset. Blood 2001, 97, 3146–3151. [Google Scholar] [CrossRef] [Green Version]
- Caligiuri, M.A. Human natural killer cells. Blood 2008, 112, 461–469. [Google Scholar] [CrossRef]
- Guidotti, L.G.; Chisari, F.V. Noncytolytic control of viral infections by the innate and adaptive immune response. Annu. Rev. Immunol. 2001, 19, 65–91. [Google Scholar] [CrossRef] [PubMed]
- Golden-Mason, L.; Rosen, H.R. Natural killer cells: Primary target for hepatitis C virus immune evasion strategies? Liver. Transpl. 2006, 12, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Irshad, M.; Khushboo, I.; Singh, S.; Singh, S. Hepatitis C virus (HCV): A review of immunological aspects. Int. Rev. Immunol. 2008, 27, 497–517. [Google Scholar] [CrossRef] [PubMed]
- Cheent, K.; Khakoo, S.I. Natural killer cells and hepatitis C: Action and reaction. Gut 2011, 60, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Golden-Mason, L.; Rosen, H.R. Natural killer cells: Multifaceted players with key roles in hepatitis C immunity. Immunol. Rev. 2013, 255, 68–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, E.O.; Kim, H.S.; Liu, D.; Peterson, M.E.; Rajagopalan, S. Controlling natural killer cell responses: Integration of signals for activation and inhibition. Annu. Rev. Immunol. 2013, 31, 227–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maini, M.K.; Peppa, D. NK cells: A double-edged sword in chronic hepatitis B virus infection. Front. Immunol. 2013, 4, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehermann, B. Pathogenesis of chronic viral hepatitis: Differential roles of T cells and NK cells. Nat. Med. 2013, 19, 859–868. [Google Scholar] [CrossRef] [Green Version]
- Swain, M.G. Hepatic NKT cells: Friend or foe? Clin. Sci. (Lond) 2008, 114, 457–466. [Google Scholar] [CrossRef] [Green Version]
- Merad, M.; Sathe, P.; Helft, J.; Miller, J.; Mortha, A. The dendritic cell lineage: Ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu. Rev. Immunol. 2013, 31, 563–604. [Google Scholar] [CrossRef] [Green Version]
- Steinman, R.M. Decisions about dendritic cells: Past, present, and future. Annu. Rev. Immunol. 2012, 30, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Swiecki, M.; Colonna, M. The multifaceted biology of plasmacytoid dendritic cells. Nat. Rev. Immunol. 2015, 15, 471–485. [Google Scholar] [CrossRef]
- Takahashi, K.; Asabe, S.; Wieland, S.; Garaigorta, U.; Gastaminza, P.; Isogawa, M.; Chisari, F.V. Plasmacytoid dendritic cells sense hepatitis C virus-infected cells, produce interferon, and inhibit infection. Proc. Natl. Acad. Sci. USA 2010, 107, 7431–7436. [Google Scholar] [CrossRef] [Green Version]
- Yoshio, S.; Kanto, T.; Kuroda, S.; Matsubara, T.; Higashitani, K.; Kakita, N.; Ishida, H.; Hiramatsu, N.; Nagano, H.; Sugiyama, M.; et al. Human blood dendritic cell antigen 3 (BDCA3)(+) dendritic cells are a potent producer of interferon-lambda in response to hepatitis C virus. Hepatology 2013, 57, 1705–1715. [Google Scholar] [CrossRef]
- Marukian, S.; Jones, C.T.; Andrus, L.; Evans, M.J.; Ritola, K.D.; Charles, E.D.; Rice, C.M.; Dustin, L.B. Cell culture-produced hepatitis C virus does not infect peripheral blood mononuclear cells. Hepatology 2008, 48, 1843–1850. [Google Scholar] [CrossRef] [Green Version]
- Dustin, L.B. Innate and Adaptive Immune Responses in Chronic HCV Infection. Curr. Drug Targets 2017, 18, 826–843. [Google Scholar] [CrossRef] [Green Version]
- Dolganiuc, A.; Kodys, K.; Kopasz, A.; Marshall, C.; Mandrekar, P.; Szabo, G. Additive inhibition of dendritic cell allostimulatory capacity by alcohol and hepatitis C is not restored by DC maturation and involves abnormal IL-10 and IL-2 induction. Alcohol. Clin. Exp. Res. 2003, 27, 1023–1031. [Google Scholar] [CrossRef]
- Sarobe, P.; Lasarte, J.J.; Zabaleta, A.; Arribillaga, L.; Arina, A.; Melero, I.; Borras-Cuesta, F.; Prieto, J. Hepatitis C virus structural proteins impair dendritic cell maturation and inhibit in vivo induction of cellular immune responses. J. Virol. 2003, 77, 10862–10871. [Google Scholar] [CrossRef] [Green Version]
- Szabo, G.; Dolganiuc, A. Subversion of plasmacytoid and myeloid dendritic cell functions in chronic HCV infection. Immunobiology 2005, 210, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Marrack, P.; McKee, A.S.; Munks, M.W. Towards an understanding of the adjuvant action of aluminium. Nat. Rev. Immunol. 2009, 9, 287–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georg, P.; Sander, L.E. Innate sensors that regulate vaccine responses. Curr. Opin. Immunol. 2019, 59, 31–41. [Google Scholar] [CrossRef]
- Gutjahr, A.; Tiraby, G.; Perouzel, E.; Verrier, B.; Paul, S. Triggering Intracellular Receptors for Vaccine Adjuvantation. Trends Immunol. 2016, 37, 573–587. [Google Scholar] [CrossRef]
- Iwasaki, A.; Medzhitov, R. Control of adaptive immunity by the innate immune system. Nat. Immunol. 2015, 16, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B. The varieties of immunological experience: Of pathogens, stress, and dendritic cells. Annu. Rev. Immunol. 2015, 33, 563–606. [Google Scholar] [CrossRef] [PubMed]
- Vasou, A.; Sultanoglu, N.; Goodbourn, S.; Randall, R.E.; Kostrikis, L.G. Targeting Pattern Recognition Receptors (PRR) for Vaccine Adjuvantation: From Synthetic PRR Agonists to the Potential of Defective Interfering Particles of Viruses. Viruses 2017, 9, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowling, J.K.; Mansell, A. Toll-like receptors: The swiss army knife of immunity and vaccine development. Clin. Transl. Immunol. 2016, 5, e85. [Google Scholar] [CrossRef] [PubMed]
- Lester, S.N.; Li, K. Toll-like receptors in antiviral innate immunity. J. Mol. Biol. 2014, 426, 1246–1264. [Google Scholar] [CrossRef]
- Kumar, H.; Kawai, T.; Akira, S. Pathogen recognition by the innate immune system. Int. Rev. Immunol. 2011, 30, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.; Garcia-Martinez, E.; Pitter, M.R.; Fucikova, J.; Spisek, R.; Zitvogel, L.; Kroemer, G.; Galluzzi, L. Trial Watch: Toll-like receptor agonists in cancer immunotherapy. Oncoimmunology 2018, 7, e1526250. [Google Scholar] [CrossRef]
- Griffiths, E.A.; Srivastava, P.; Matsuzaki, J.; Brumberger, Z.; Wang, E.S.; Kocent, J.; Miller, A.; Roloff, G.W.; Wong, H.Y.; Paluch, B.E.; et al. NY-ESO-1 Vaccination in Combination with Decitabine Induces Antigen-Specific T-lymphocyte Responses in Patients with Myelodysplastic Syndrome. Clin. Cancer Res. 2018, 24, 1019–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabbatini, P.; Tsuji, T.; Ferran, L.; Ritter, E.; Sedrak, C.; Tuballes, K.; Jungbluth, A.A.; Ritter, G.; Aghajanian, C.; Bell-McGuinn, K.; et al. Phase I trial of overlapping long peptides from a tumor self-antigen and poly-ICLC shows rapid induction of integrated immune response in ovarian cancer patients. Clin. Cancer Res. 2012, 18, 6497–6508. [Google Scholar] [CrossRef] [Green Version]
- Harper, D.M.; Franco, E.L.; Wheeler, C.; Ferris, D.G.; Jenkins, D.; Schuind, A.; Zahaf, T.; Innis, B.; Naud, P.; De Carvalho, N.S.; et al. Efficacy of a bivalent L1 virus-like particle vaccine in prevention of infection with human papillomavirus types 16 and 18 in young women: A randomised controlled trial. Lancet 2004, 364, 1757–1765. [Google Scholar] [CrossRef]
- Paavonen, J.; Jenkins, D.; Bosch, F.X.; Naud, P.; Salmeron, J.; Wheeler, C.M.; Chow, S.N.; Apter, D.L.; Kitchener, H.C.; Castellsague, X.; et al. Efficacy of a prophylactic adjuvanted bivalent L1 virus-like-particle vaccine against infection with human papillomavirus types 16 and 18 in young women: An interim analysis of a phase III double-blind, randomised controlled trial. Lancet 2007, 369, 2161–2170. [Google Scholar] [CrossRef]
- Casares, S.; Brumeanu, T.D.; Richie, T.L. The RTS,S malaria vaccine. Vaccine 2010, 28, 4880–4894. [Google Scholar] [CrossRef] [Green Version]
- Behzad, H.; Huckriede, A.L.; Haynes, L.; Gentleman, B.; Coyle, K.; Wilschut, J.C.; Kollmann, T.R.; Reed, S.G.; McElhaney, J.E. GLA-SE, a synthetic toll-like receptor 4 agonist, enhances T-cell responses to influenza vaccine in older adults. J. Infect. Dis. 2012, 205, 466–473. [Google Scholar] [CrossRef] [Green Version]
- Penn-Nicholson, A.; Tameris, M.; Smit, E.; Day, T.A.; Musvosvi, M.; Jayashankar, L.; Vergara, J.; Mabwe, S.; Bilek, N.; Geldenhuys, H.; et al. Safety and immunogenicity of the novel tuberculosis vaccine ID93 + GLA-SE in BCG-vaccinated healthy adults in South Africa: A randomised, double-blind, placebo-controlled phase 1 trial. Lancet Respir. Med. 2018, 6, 287–298. [Google Scholar] [CrossRef]
- Dupont, J.; Altclas, J.; Lepetic, A.; Lombardo, M.; Vazquez, V.; Salgueira, C.; Seigelchifer, M.; Arndtz, N.; Antunez, E.; von Eschen, K.; et al. A controlled clinical trial comparing the safety and immunogenicity of a new adjuvanted hepatitis B vaccine with a standard hepatitis B vaccine. Vaccine 2006, 24, 7167–7174. [Google Scholar] [CrossRef]
- Del Giudice, G.; Rappuoli, R.; Didierlaurent, A.M. Correlates of adjuvanticity: A review on adjuvants in licensed vaccines. Semin. Immunol. 2018, 39, 14–21. [Google Scholar] [CrossRef]
- Turley, C.B.; Rupp, R.E.; Johnson, C.; Taylor, D.N.; Wolfson, J.; Tussey, L.; Kavita, U.; Stanberry, L.; Shaw, A. Safety and immunogenicity of a recombinant M2e-flagellin influenza vaccine (STF2.4xM2e) in healthy adults. Vaccine 2011, 29, 5145–5152. [Google Scholar] [CrossRef]
- Taylor, D.N.; Treanor, J.J.; Sheldon, E.A.; Johnson, C.; Umlauf, S.; Song, L.; Kavita, U.; Liu, G.; Tussey, L.; Ozer, K.; et al. Development of VAX128, a recombinant hemagglutinin (HA) influenza-flagellin fusion vaccine with improved safety and immune response. Vaccine 2012, 30, 5761–5769. [Google Scholar] [CrossRef]
- Hornung, V.; Rothenfusser, S.; Britsch, S.; Krug, A.; Jahrsdorfer, B.; Giese, T.; Endres, S.; Hartmann, G. Quantitative expression of toll-like receptor 1-10 mRNA in cellular subsets of human peripheral blood mononuclear cells and sensitivity to CpG oligodeoxynucleotides. J. Immunol. 2002, 168, 4531–4537. [Google Scholar] [CrossRef] [Green Version]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-specific recognition of single-stranded RNA via toll-like receptor 7 and 8. Science 2004, 303, 1526–1529. [Google Scholar] [CrossRef] [Green Version]
- Diebold, S.S.; Kaisho, T.; Hemmi, H.; Akira, S.; Reis e Sousa, C. Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science 2004, 303, 1529–1531. [Google Scholar] [CrossRef]
- Tanji, H.; Ohto, U.; Shibata, T.; Taoka, M.; Yamauchi, Y.; Isobe, T.; Miyake, K.; Shimizu, T. Toll-like receptor 8 senses degradation products of single-stranded RNA. Nat. Struct. Mol. Biol. 2015, 22, 109–115. [Google Scholar] [CrossRef]
- Zhang, Z.; Ohto, U.; Shibata, T.; Krayukhina, E.; Taoka, M.; Yamauchi, Y.; Tanji, H.; Isobe, T.; Uchiyama, S.; Miyake, K.; et al. Structural Analysis Reveals that Toll-like Receptor 7 Is a Dual Receptor for Guanosine and Single-Stranded RNA. Immunity 2016, 45, 737–748. [Google Scholar] [CrossRef] [Green Version]
- Hemmi, H.; Kaisho, T.; Takeuchi, O.; Sato, S.; Sanjo, H.; Hoshino, K.; Horiuchi, T.; Tomizawa, H.; Takeda, K.; Akira, S. Small anti-viral compounds activate immune cells via the TLR7 MyD88-dependent signaling pathway. Nat. Immunol. 2002, 3, 196–200. [Google Scholar] [CrossRef]
- Sospedra, M.; Schippling, S.; Yousef, S.; Jelcic, I.; Bofill-Mas, S.; Planas, R.; Stellmann, J.P.; Demina, V.; Cinque, P.; Garcea, R.; et al. Treating progressive multifocal leukoencephalopathy with interleukin 7 and vaccination with JC virus capsid protein VP1. Clin. Infect. Dis. 2014, 59, 1588–1592. [Google Scholar] [CrossRef] [PubMed]
- Ray, U.; Cinque, P.; Gerevini, S.; Longo, V.; Lazzarin, A.; Schippling, S.; Martin, R.; Buck, C.B.; Pastrana, D.V. JC polyomavirus mutants escape antibody-mediated neutralization. Sci. Transl. Med. 2015, 7, 306ra151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, I.F.; Zhang, A.J.; To, K.K.; Chan, J.F.; Li, C.; Zhu, H.S.; Li, P.; Li, C.; Chan, T.C.; Cheng, V.C.; et al. Immunogenicity of intradermal trivalent influenza vaccine with topical imiquimod: A double blind randomized controlled trial. Clin. Infect. Dis. 2014, 59, 1246–1255. [Google Scholar] [CrossRef] [PubMed]
- Wille-Reece, U.; Flynn, B.J.; Lore, K.; Koup, R.A.; Kedl, R.M.; Mattapallil, J.J.; Weiss, W.R.; Roederer, M.; Seder, R.A. HIV Gag protein conjugated to a Toll-like receptor 7/8 agonist improves the magnitude and quality of Th1 and CD8+ T cell responses in nonhuman primates. Proc. Natl. Acad. Sci. USA 2005, 102, 15190–15194. [Google Scholar] [CrossRef] [Green Version]
- Lissina, A.; Briceno, O.; Afonso, G.; Larsen, M.; Gostick, E.; Price, D.A.; Mallone, R.; Appay, V. Priming of Qualitatively Superior Human Effector CD8+ T Cells Using TLR8 Ligand Combined with FLT3 Ligand. J. Immunol. 2016, 196, 256–263. [Google Scholar] [CrossRef] [Green Version]
- Dowling, D.J. Recent Advances in the Discovery and Delivery of TLR7/8 Agonists as Vaccine Adjuvants. Immunohorizons 2018, 2, 185–197. [Google Scholar] [CrossRef] [Green Version]
- Phillips, B.; Van Rompay, K.K.A.; Rodriguez-Nieves, J.; Lorin, C.; Koutsoukos, M.; Tomai, M.; Fox, C.B.; Eudailey, J.; Dennis, M.; Alam, S.M.; et al. Adjuvant-Dependent Enhancement of HIV Env-Specific Antibody Responses in Infant Rhesus Macaques. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Campbell, J.D.; Cho, Y.; Foster, M.L.; Kanzler, H.; Kachura, M.A.; Lum, J.A.; Ratcliffe, M.J.; Sathe, A.; Leishman, A.J.; Bahl, A.; et al. CpG-containing immunostimulatory DNA sequences elicit TNF-alpha-dependent toxicity in rodents but not in humans. J. Clin. Investig. 2009, 119, 2564–2576. [Google Scholar] [CrossRef]
- Kobayashi, H.; Horner, A.A.; Takabayashi, K.; Nguyen, M.D.; Huang, E.; Cinman, N.; Raz, E. Immunostimulatory DNA pre-priming: A novel approach for prolonged Th1-biased immunity. Cell Immunol. 1999, 198, 69–75. [Google Scholar] [CrossRef]
- Tighe, H.; Takabayashi, K.; Schwartz, D.; Van Nest, G.; Tuck, S.; Eiden, J.J.; Kagey-Sobotka, A.; Creticos, P.S.; Lichtenstein, L.M.; Spiegelberg, H.L.; et al. Conjugation of immunostimulatory DNA to the short ragweed allergen amb a 1 enhances its immunogenicity and reduces its allergenicity. J. Allergy Clin. Immunol. 2000, 106, 124–134. [Google Scholar] [CrossRef]
- Krug, A.; Towarowski, A.; Britsch, S.; Rothenfusser, S.; Hornung, V.; Bals, R.; Giese, T.; Engelmann, H.; Endres, S.; Krieg, A.M.; et al. Toll-like receptor expression reveals CpG DNA as a unique microbial stimulus for plasmacytoid dendritic cells which synergizes with CD40 ligand to induce high amounts of IL-12. Eur. J. Immunol. 2001, 31, 3026–3037. [Google Scholar] [CrossRef]
- Krieg, A.M.; Efler, S.M.; Wittpoth, M.; Al Adhami, M.J.; Davis, H.L. Induction of systemic TH1-like innate immunity in normal volunteers following subcutaneous but not intravenous administration of CPG 7909, a synthetic B-class CpG oligodeoxynucleotide TLR9 agonist. J. Immunother. 2004, 27, 460–471. [Google Scholar] [CrossRef] [PubMed]
- Speiser, D.E.; Lienard, D.; Rufer, N.; Rubio-Godoy, V.; Rimoldi, D.; Lejeune, F.; Krieg, A.M.; Cerottini, J.C.; Romero, P. Rapid and strong human CD8+ T cell responses to vaccination with peptide, IFA, and CpG oligodeoxynucleotide 7909. J. Clin. Investig. 2005, 115, 739–746. [Google Scholar] [CrossRef]
- Schillie, S.; Harris, A.; Link-Gelles, R.; Romero, J.; Ward, J.; Nelson, N. Recommendations of the Advisory Committee on Immunization Practices for Use of a Hepatitis B Vaccine with a Novel Adjuvant. MMWR Morb. Mortal. Wkly Rep. 2018, 67, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.; Lentino, J.; Kopp, J.; Murray, L.; Ellison, W.; Rhee, M.; Shockey, G.; Akella, L.; Erby, K.; Heyward, W.L.; et al. Immunogenicity of a two-dose investigational hepatitis B vaccine, HBsAg-1018, using a toll-like receptor 9 agonist adjuvant compared with a licensed hepatitis B vaccine in adults. Vaccine 2018, 36, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Schenten, D.; Medzhitov, R. The control of adaptive immune responses by the innate immune system. Adv. Immunol. 2011, 109, 87–124. [Google Scholar] [CrossRef]
- Griffin, M.E.; Hespen, C.W.; Wang, Y.C.; Hang, H.C. Translation of peptidoglycan metabolites into immunotherapeutics. Clin. Transl. Immunol. 2019, 8, e1095. [Google Scholar] [CrossRef]
- Sun, L.; Wu, J.; Du, F.; Chen, X.; Chen, Z.J. Cyclic GMP-AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway. Science 2013, 339, 786–791. [Google Scholar] [CrossRef] [Green Version]
- Ng, K.W.; Marshall, E.A.; Bell, J.C.; Lam, W.L. cGAS-STING and Cancer: Dichotomous Roles in Tumor Immunity and Development. Trends Immunol. 2018, 39, 44–54. [Google Scholar] [CrossRef]
- Cerboni, S.; Jeremiah, N.; Gentili, M.; Gehrmann, U.; Conrad, C.; Stolzenberg, M.C.; Picard, C.; Neven, B.; Fischer, A.; Amigorena, S.; et al. Intrinsic antiproliferative activity of the innate sensor STING in T lymphocytes. J. Exp. Med. 2017, 214, 1769–1785. [Google Scholar] [CrossRef] [Green Version]
- Trinchieri, G.; Sher, A. Cooperation of Toll-like receptor signals in innate immune defence. Nat. Rev. Immunol. 2007, 7, 179–190. [Google Scholar] [CrossRef]
- Napolitani, G.; Rinaldi, A.; Bertoni, F.; Sallusto, F.; Lanzavecchia, A. Selected Toll-like receptor agonist combinations synergistically trigger a T helper type 1-polarizing program in dendritic cells. Nat. Immunol. 2005, 6, 769–776. [Google Scholar] [CrossRef]
- Kester, K.E.; Cummings, J.F.; Ofori-Anyinam, O.; Ockenhouse, C.F.; Krzych, U.; Moris, P.; Schwenk, R.; Nielsen, R.A.; Debebe, Z.; Pinelis, E.; et al. Randomized, double-blind, phase 2a trial of falciparum malaria vaccines RTS,S/AS01B and RTS,S/AS02A in malaria-naive adults: Safety, efficacy, and immunologic associates of protection. J. Infect. Dis. 2009, 200, 337–346. [Google Scholar] [CrossRef]
- Leroux-Roels, G.; Leroux-Roels, I.; Clement, F.; Ofori-Anyinam, O.; Lievens, M.; Jongert, E.; Moris, P.; Ballou, W.R.; Cohen, J. Evaluation of the immune response to RTS,S/AS01 and RTS,S/AS02 adjuvanted vaccines: Randomized, double-blind study in malaria-naive adults. Hum. Vaccin. Immunother. 2014, 10, 2211–2219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Drug Administration. Shingrix. Available online: https://www.fda.gov/vaccines-blood-biologics/vaccines/shingrix (accessed on 31 May 2020).
- Laupeze, B.; Herve, C.; Di Pasquale, A.; Tavares Da Silva, F. Adjuvant Systems for vaccines: 13years of post-licensure experience in diverse populations have progressed the way adjuvanted vaccine safety is investigated and understood. Vaccine 2019, 37, 5670–5680. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.R.; Barnes, E.; Cox, A.L. Approaches, Progress, and Challenges to Hepatitis C Vaccine Development. Gastroenterology 2019, 156, 418–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, S.; Lauer, G.; Isba, R.; Walker, B.; Klenerman, P. Cellular immune responses against hepatitis C virus: The evidence base 2002. Clin. Exp. Immunol. 2002, 128, 195–203. [Google Scholar] [CrossRef]
- Lechner, F.; Wong, D.K.; Dunbar, P.R.; Chapman, R.; Chung, R.T.; Dohrenwend, P.; Robbins, G.; Phillips, R.; Klenerman, P.; Walker, B.D. Analysis of successful immune responses in persons infected with hepatitis C virus. J. Exp. Med. 2000, 191, 1499–1512. [Google Scholar] [CrossRef] [Green Version]
- Day, C.L.; Lauer, G.M.; Robbins, G.K.; McGovern, B.; Wurcel, A.G.; Gandhi, R.T.; Chung, R.T.; Walker, B.D. Broad specificity of virus-specific CD4+ T-helper-cell responses in resolved hepatitis C virus infection. J. Virol. 2002, 76, 12584–12595. [Google Scholar] [CrossRef] [Green Version]
- Grakoui, A.; Shoukry, N.H.; Woollard, D.J.; Han, J.H.; Hanson, H.L.; Ghrayeb, J.; Murthy, K.K.; Rice, C.M.; Walker, C.M. HCV persistence and immune evasion in the absence of memory T cell help. Science 2003, 302, 659–662. [Google Scholar] [CrossRef] [Green Version]
- Shoukry, N.H.; Grakoui, A.; Houghton, M.; Chien, D.Y.; Ghrayeb, J.; Reimann, K.A.; Walker, C.M. Memory CD8+ T cells are required for protection from persistent hepatitis C virus infection. J. Exp. Med. 2003, 197, 1645–1655. [Google Scholar] [CrossRef]
- Bassett, S.E.; Guerra, B.; Brasky, K.; Miskovsky, E.; Houghton, M.; Klimpel, G.R.; Lanford, R.E. Protective immune response to hepatitis C virus in chimpanzees rechallenged following clearance of primary infection. Hepatology 2001, 33, 1479–1487. [Google Scholar] [CrossRef] [PubMed]
- Major, M.E.; Mihalik, K.; Puig, M.; Rehermann, B.; Nascimbeni, M.; Rice, C.M.; Feinstone, S.M. Previously infected and recovered chimpanzees exhibit rapid responses that control hepatitis C virus replication upon rechallenge. J. Virol. 2002, 76, 6586–6595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimbeni, M.; Mizukoshi, E.; Bosmann, M.; Major, M.E.; Mihalik, K.; Rice, C.M.; Feinstone, S.M.; Rehermann, B. Kinetics of CD4+ and CD8+ memory T-cell responses during hepatitis C virus rechallenge of previously recovered chimpanzees. J. Virol. 2003, 77, 4781–4793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swadling, L.; Capone, S.; Antrobus, R.D.; Brown, A.; Richardson, R.; Newell, E.W.; Halliday, J.; Kelly, C.; Bowen, D.; Fergusson, J.; et al. A human vaccine strategy based on chimpanzee adenoviral and MVA vectors that primes, boosts, and sustains functional HCV-specific T cell memory. Sci. Transl. Med. 2014, 6, 261ra153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Institute of Allergy and Infectious Diseaes. Trial Evaluating Experimental Hepatitis C Vaccine Concludes. Available online: https://www.niaid.nih.gov/news-events/trial-evaluating-experimental-hepatitis-c-vaccine-concludes (accessed on 31 May 2020).
- Lee, H.; Jeong, M.; Oh, J.; Cho, Y.; Shen, X.; Stone, J.; Yan, J.; Rothkopf, Z.; Khan, A.S.; Cho, B.M.; et al. Preclinical evaluation of multi antigenic HCV DNA vaccine for the prevention of Hepatitis C virus infection. Sci. Rep. 2017, 7, 43531. [Google Scholar] [CrossRef]
- Han, J.W.; Sung, P.S.; Hong, S.H.; Lee, H.; Koh, J.Y.; Lee, H.; White, S.; Maslow, J.N.; Weiner, D.B.; Park, S.H.; et al. IFNL3-adjuvanted HCV DNA vaccine reduces regulatory T cell frequency and increases virus-specific T cell responses. J. Hepatol. 2020. [Google Scholar] [CrossRef]
- Lu, L.; Barbi, J.; Pan, F. The regulation of immune tolerance by FOXP3. Nat. Rev. Immunol. 2017, 17, 703–717. [Google Scholar] [CrossRef]
- Langhans, B.; Nischalke, H.D.; Kramer, B.; Hausen, A.; Dold, L.; van Heteren, P.; Huneburg, R.; Nattermann, J.; Strassburg, C.P.; Spengler, U. Increased peripheral CD4(+) regulatory T cells persist after successful direct-acting antiviral treatment of chronic hepatitis C. J. Hepatol. 2017, 66, 888–896. [Google Scholar] [CrossRef]
- Dougan, G.; Hormaeche, C. How bacteria and their products provide clues to vaccine and adjuvant development. Vaccine 2006, 24 (Suppl. 2), S13–S19. [Google Scholar] [CrossRef]
- Pouriayevali, M.H.; Bamdad, T.; Sadat, S.M.; Sadeghi, S.A.; Sabahi, F.; Mahdavi, M.; Aghasadeghi, M.R. Listeriolysin O immunogenetic adjuvant enhanced potency of hepatitis C virus NS3 DNA vaccine. IUBMB Life 2019, 71, 1645–1652. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, S.; Irani, S.; Bolhassani, A.; Sadat, S.M. Simultaneous use of natural adjuvants and cell penetrating peptides improves HCV NS3 antigen-specific immune responses. Immunol. Lett. 2019, 212, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Basirnejad, M.; Bolhassani, A. Development of HCV Therapeutic Vaccines Using Hp91 Peptide and Small Heat Shock Protein 20 as an Adjuvant. Protein. Pept. Lett. 2018, 25, 924–932. [Google Scholar] [CrossRef]
- Borhani, K.; Bamdad, T.; Hashempour, T. Lenalidomide acts as an adjuvant for HCV DNA vaccine. Int. Immunopharmacol. 2017, 48, 231–240. [Google Scholar] [CrossRef]
- Levander, S.; Sallberg, M.; Ahlen, G.; Frelin, L. A non-human hepadnaviral adjuvant for hepatitis C virus-based genetic vaccines. Vaccine 2016, 34, 2821–2833. [Google Scholar] [CrossRef] [Green Version]
- Kuprianov, V.V.; Nikolaeva, L.I.; Zykova, A.A.; Dedova, A.V.; Grishechkin, A.E.; Kapustin, I.V.; Kotlyarov, R.Y.; Ravin, N.V. Combination of three adjuvants enhances the immunogenicity of a recombinant protein containing the CTL epitopes of non-structural proteins of hepatitis C virus. Virus Res. 2020, 284, 197984. [Google Scholar] [CrossRef]
- Javadi, F.; Rahimi, P.; Modarresi, M.H.; Bolhassani, A.; Shafiee Ardestani, M.; Sadat, S.M. G2 Dendrimer as a Carrier Can Enhance Immune Responses Against HCV-NS3 Protein in BALB/c Mice. Avicenna J. Med. Biotechnol. 2019, 11, 292–298. [Google Scholar]
- Liu, J.; Feng, X.; Chen, Z.; Yang, X.; Shen, Z.; Guo, M.; Deng, F.; Liu, Y.; Zhang, H.; Chen, C. The adjuvant effect of C60(OH)22 nanoparticles promoting both humoral and cellular immune responses to HCV recombinant proteins. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 97, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Masalova, O.V.; Lesnova, E.I.; Onishchuk, A.A.; Ivanova, A.M.; Gerasimova, E.V.; Ivanov, A.V.; Narovlyansky, A.N.; Sanin, A.V.; Pronin, A.V.; Kushch, A.A. Polyprenyl Phosphates Induce a High Humoral and Cellular Response to Immunization with Recombinant Proteins of the Replicative Complex of the Hepatitis C Virus. Dokl. Biochem. Biophys. 2018, 482, 261–263. [Google Scholar] [CrossRef]
- Filskov, J.; Mikkelsen, M.; Hansen, P.R.; Christensen, J.P.; Thomsen, A.R.; Andersen, P.; Bukh, J.; Agger, E.M. Broadening CD4(+) and CD8(+) T Cell Responses against Hepatitis C Virus by Vaccination with NS3 Overlapping Peptide Panels in Cross-Priming Liposomes. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Filskov, J.; Andersen, P.; Agger, E.M.; Bukh, J. HCV p7 as a novel vaccine-target inducing multifunctional CD4(+) and CD8(+) T-cells targeting liver cells expressing the viral antigen. Sci. Rep. 2019, 9, 14085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hekmat, S.; Sadat, S.M.; Aslani, M.M.; Mahdavi, M.; Bolhassani, A.; Asgar Halvaee, F.; Ghahari, S.M.M.; Aghasadeghi, M.R.; Siadat, S.D. Truncated Core/NS3 Fusion Protein of HCV Adjuvanted with Outer Membrane Vesicles of Neisseria meningitidis Serogroup B: Potent Inducer of the Murine Immune System. Iran Biomed. J. 2019, 23, 235–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikram, A.; Zaheer, T.; Awan, F.M.; Obaid, A.; Naz, A.; Hanif, R.; Paracha, R.Z.; Ali, A.; Naveed, A.K.; Janjua, H.A. Exploring NS3/4A, NS5A and NS5B proteins to design conserved subunit multi-epitope vaccine against HCV utilizing immunoinformatics approaches. Sci. Rep. 2018, 8, 16107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, B.; Xia, T. Nanomaterial-Based Vaccine Adjuvants. J. Mater. Chem. B 2016, 4, 5496–5509. [Google Scholar] [CrossRef] [PubMed]
- Pati, R.; Shevtsov, M.; Sonawane, A. Nanoparticle Vaccines Against Infectious Diseases. Front. Immunol. 2018, 9, 2224. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Liu, Y.; Chen, Z.; Li, W.; Liu, Y.; Wang, L.; Ma, L.; Shao, Y.; Zhao, Y.; Chen, C. Morphologically virus-like fullerenol nanoparticles act as the dual-functional nanoadjuvant for HIV-1 vaccine. Adv. Mater. 2013, 25, 5928–5936. [Google Scholar] [CrossRef]
- Korsholm, K.S.; Hansen, J.; Karlsen, K.; Filskov, J.; Mikkelsen, M.; Lindenstrom, T.; Schmidt, S.T.; Andersen, P.; Christensen, D. Induction of CD8+ T-cell responses against subunit antigens by the novel cationic liposomal CAF09 adjuvant. Vaccine 2014, 32, 3927–3935. [Google Scholar] [CrossRef]
- Hekmat, S.; Siadat, S.D.; Aghasadeghi, M.R.; Sadat, S.M.; Bahramali, G.; Aslani, M.M.; Mahdavi, M.; Shahbazi, S. From in-silico immunogenicity verification to in vitro expression of recombinant Core-NS3 fusion protein of HCV. Bratisl. Lek. Listy 2017, 118, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Aldaco, K.; Torres-Reyes, L.A.; Ojeda-Granados, C.; Jose-Abrego, A.; Fierro, N.A.; Roman, S. Immunometabolic Effect of Cholesterol in Hepatitis C Infection: Implications in Clinical Management and Antiviral Therapy. Ann. Hepatol. 2018, 17, 908–919. [Google Scholar] [CrossRef]
- Frey, S.E.; Houghton, M.; Coates, S.; Abrignani, S.; Chien, D.; Rosa, D.; Pileri, P.; Ray, R.; Di Bisceglie, A.M.; Rinella, P.; et al. Safety and immunogenicity of HCV E1E2 vaccine adjuvanted with MF59 administered to healthy adults. Vaccine 2010, 28, 6367–6373. [Google Scholar] [CrossRef] [Green Version]
- Law, J.L.; Chen, C.; Wong, J.; Hockman, D.; Santer, D.M.; Frey, S.E.; Belshe, R.B.; Wakita, T.; Bukh, J.; Jones, C.T.; et al. A hepatitis C virus (HCV) vaccine comprising envelope glycoproteins gpE1/gpE2 derived from a single isolate elicits broad cross-genotype neutralizing antibodies in humans. PLoS ONE 2013, 8, e59776. [Google Scholar] [CrossRef] [PubMed]
- Ray, R.; Meyer, K.; Banerjee, A.; Basu, A.; Coates, S.; Abrignani, S.; Houghton, M.; Frey, S.E.; Belshe, R.B. Characterization of antibodies induced by vaccination with hepatitis C virus envelope glycoproteins. J. Infect. Dis. 2010, 202, 862–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.A.; Bhat, R.; Hockman, D.; Logan, M.; Chen, C.; Levin, A.; Frey, S.E.; Belshe, R.B.; Tyrrell, D.L.; Law, J.L.; et al. Recombinant hepatitis C virus envelope glycoprotein vaccine elicits antibodies targeting multiple epitopes on the envelope glycoproteins associated with broad cross-neutralization. J. Virol. 2014, 88, 14278–14288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Nagy, K.; Chavez, D.; Willis, S.; McBride, R.; Giang, E.; Honda, A.; Bukh, J.; Ordoukhanian, P.; Zhu, J.; et al. Antibody Responses to Immunization With HCV Envelope Glycoproteins as a Baseline for B-Cell-Based Vaccine Development. Gastroenterology 2020, 158, 1058–1071.e6. [Google Scholar] [CrossRef]
- Landi, A.; Law, J.; Hockman, D.; Logan, M.; Crawford, K.; Chen, C.; Kundu, J.; Ebensen, T.; Guzman, C.A.; Deschatelets, L.; et al. Superior immunogenicity of HCV envelope glycoproteins when adjuvanted with cyclic-di-AMP, a STING activator or archaeosomes. Vaccine 2017, 35, 6949–6956. [Google Scholar] [CrossRef]
- Akache, B.; Deschatelets, L.; Harrison, B.A.; Dudani, R.; Stark, F.C.; Jia, Y.; Landi, A.; Law, J.L.M.; Logan, M.; Hockman, D.; et al. Effect of Different Adjuvants on the Longevity and Strength of Humoral and Cellular Immune Responses to the HCV Envelope Glycoproteins. Vaccines 2019, 7, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, L.; Dicaire, C.J.; Patel, G.B.; Sprott, G.D. Archaeosome vaccine adjuvants induce strong humoral, cell-mediated, and memory responses: Comparison to conventional liposomes and alum. Infect. Immun. 2000, 68, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Akache, B.; Stark, F.C.; Iqbal, U.; Chen, W.; Jia, Y.; Krishnan, L.; McCluskie, M.J. Safety and biodistribution of sulfated archaeal glycolipid archaeosomes as vaccine adjuvants. Hum. Vaccin. Immunother. 2018, 14, 1746–1759. [Google Scholar] [CrossRef]
- Akache, B.; Stark, F.C.; Jia, Y.; Deschatelets, L.; Dudani, R.; Harrison, B.A.; Agbayani, G.; Williams, D.; Jamshidi, M.P.; Krishnan, L.; et al. Sulfated archaeol glycolipids: Comparison with other immunological adjuvants in mice. PLoS ONE 2018, 13, e0208067. [Google Scholar] [CrossRef]
- Masavuli, M.G.; Wijesundara, D.K.; Underwood, A.; Christiansen, D.; Earnest-Silveira, L.; Bull, R.; Torresi, J.; Gowans, E.J.; Grubor-Bauk, B. A Hepatitis C Virus DNA Vaccine Encoding a Secreted, Oligomerized Form of Envelope Proteins Is Highly Immunogenic and Elicits Neutralizing Antibodies in Vaccinated Mice. Front. Immunol. 2019, 10, 1145. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, D.; Earnest-Silveira, L.; Chua, B.; Boo, I.; Drummer, H.E.; Grubor-Bauk, B.; Gowans, E.J.; Jackson, D.C.; Torresi, J. Antibody Responses to a Quadrivalent Hepatitis C Viral-Like Particle Vaccine Adjuvanted with Toll-Like Receptor 2 Agonists. Viral. Immunol. 2018, 31, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Olivera, S.; Perez, A.; Falcon, V.; Urquiza, D.; Pichardo, D.; Martinez-Donato, G. Protective cellular immune response against hepatitis C virus elicited by chimeric protein formulations in BALB/c mice. Arch. Virol. 2020, 165, 593–607. [Google Scholar] [CrossRef] [PubMed]

| PRR Type | Class of Activated Innate Receptor/Pathway | Adjuvant Name | Composition | Main Stimulated Immune Responses |
|---|---|---|---|---|
| TLRs | TLR3 | Poly(I:C) or Poly(I:C) stabilized with poly-L-lysine | dsRNA analogs | Ab response, CD8+ T-cell response, Th1 type immunity |
| TLR4 | MPLA, GLA-SE, RC-529 | MPL GLA AGP | Ab response CD8+ T-cell response Th1 type immunity | |
| TLR5 | Flagellin fused to antigen | Bacterial flagellin | Ab response, Th1/Th2 response | |
| TLR7, 8 or both | Imiquimod/R837 (TLR7), Resiquimod/R848 (TLR7/8), 3M-052 (TLR7/8) | Imidazoquinoline analogs | Ab response, CD4+/CD8+ T-cell response, Th1 type immunity | |
| TLR9 | CpG-ODN | Synthetic ODN with optimized CpG motifs | Ab response, CD8+ T-cell response, Th1 type immunity | |
| NLRs | RIG-I MDA-5 | M8, Defective interfering RNA | dsRNA analogs | Ab response, CD4+/CD8+ T-cell response |
| RLRs | Nod1 Nod2 | iE-DAP, MDP | Bacterial peptidoglycan analogs | Ab response |
| CDs | STING | c-di-GAMP | Bacterial cyclic dinucleotides | Ab response, CD8+ T-cell response, Th1 type immunity |
| Adjuvant Name | Composition | Vaccine Type | Main Stimulated Immune Responses |
|---|---|---|---|
| AS01 | MPLA + QS-21 in liposomes | Mosquirix: Malaria, Shingrix: Herpes zoster | Ab response, CD8+ T-cell response, Th1 type immunity |
| AS02 | MPLA + QS-21 in oil-in-water emulsions | HIV, tuberculosis, HBV | Ab response, Th1 type immunity |
| AS03 | Squalene + α-tocopherol + polysorbate 80 | Pandemrix, Arepanrix: Influenza H1N1, Prepandrix: Influenza pre-H5N1 | Ab response, Th1/Th2 response |
| AS04 | MPLA + aluminum hydroxide | Fendrix: HBV Cervarix: HPV 16/18 | Ab response, Th1 type immunity |
| MF59 | Squalene + polysorbate 80 + sorbitan trioleate | Influenza | Ab response, Th1/Th2 response |
| AF03 | Squalene + polyoxyethylene cetostearyl ether + mannitol + sorbitan oleate | Influenza | Ab response |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sepulveda-Crespo, D.; Resino, S.; Martinez, I. Innate Immune Response against Hepatitis C Virus: Targets for Vaccine Adjuvants. Vaccines 2020, 8, 313. https://doi.org/10.3390/vaccines8020313
Sepulveda-Crespo D, Resino S, Martinez I. Innate Immune Response against Hepatitis C Virus: Targets for Vaccine Adjuvants. Vaccines. 2020; 8(2):313. https://doi.org/10.3390/vaccines8020313
Chicago/Turabian StyleSepulveda-Crespo, Daniel, Salvador Resino, and Isidoro Martinez. 2020. "Innate Immune Response against Hepatitis C Virus: Targets for Vaccine Adjuvants" Vaccines 8, no. 2: 313. https://doi.org/10.3390/vaccines8020313
APA StyleSepulveda-Crespo, D., Resino, S., & Martinez, I. (2020). Innate Immune Response against Hepatitis C Virus: Targets for Vaccine Adjuvants. Vaccines, 8(2), 313. https://doi.org/10.3390/vaccines8020313