Search of Potential Vaccine Candidates against Trueperella pyogenes Infections through Proteomic and Bioinformatic Analysis

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. “Shaving” of Bacterial Live Cells and Peptide Extraction
2.3. Liquid Chromatography-Mass Spectrometry (LC-MS/MS) Analysis
2.4. Database Searching and Protein Identification
2.5. Bioinformatic Analysis of Protein Sequences
2.6. Data Analysis and Statistics
3. Results
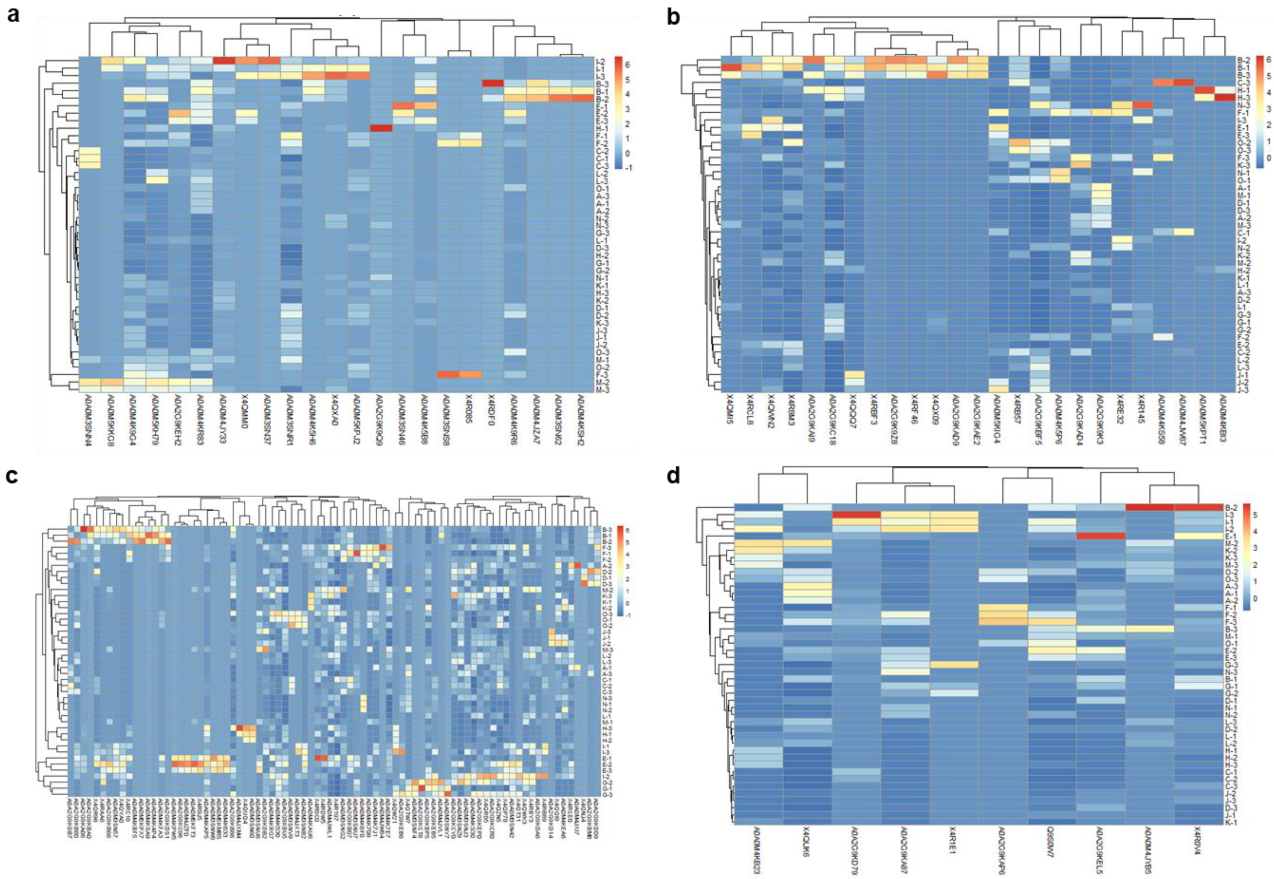
3.1. Describing the “Pan-Surfome” of T. pyogenes
3.2. Analysis of Differences in Surface Protein Abundances of T. pyogenes Clinical Isolates
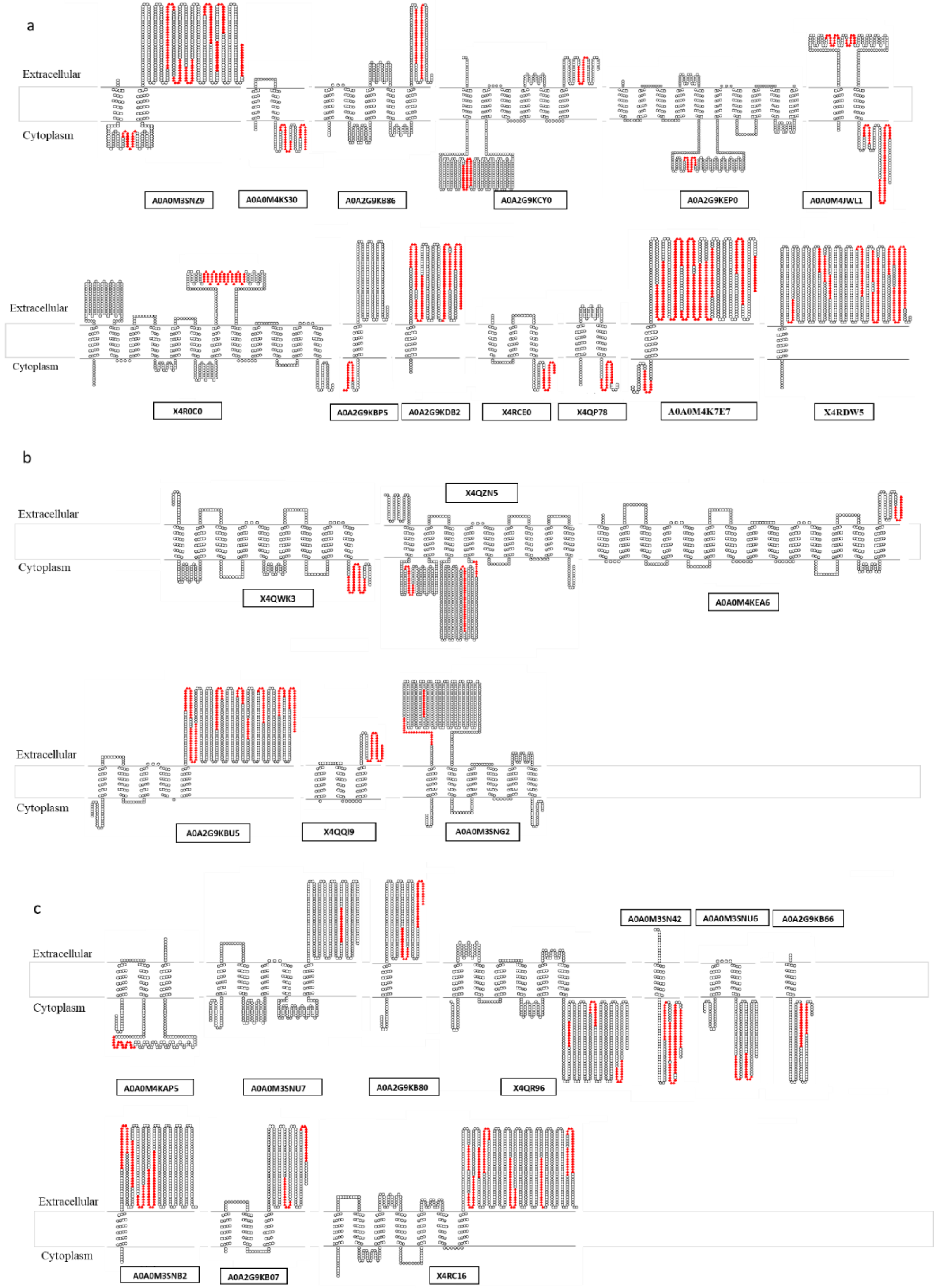
3.3. Ranking of Proteins from the “Pan-Surfome” of T. pyogenes Based on Their Potential as Putative Vaccine Candidates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Wareth, G.; El-Diasty, M.; Melzer, F.; Murugaiyan, J.; Abdulmawjood, A.; Sprague, L.D.; Neubauer, H. Trueperella pyogenes and brucella abortus coinfection in a dog and a cat on a dairy farm in Egypt with recurrent cases of mastitis and abortion. Vet. Med. Int. 2018, 2018, 2056436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rzewuska, M.; Kwiecień, E.; Chrobak-Chmiel, D.; Kizerwetter-Świda, M.; Stefańska, I.; Gieryńska, M. Pathogenicity and Virulence of Trueperella pyogenes: A Review. Int. J. Mol. Sci. 2019, 20, 2737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, M.G.; Risseti, R.M.; Bolaños, C.A.D.; Caffaro, K.A.; de Morais, A.C.B.; Lara, G.H.B.; Zamprogna, T.O.; Paes, A.C.; Listoni, F.J.P.; Franco, M.M.J. Trueperella pyogenes multispecies infections in domestic animals: A retrospective study of 144 cases (2002 to 2012). Vet. Q. 2015, 35, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Galán-Relaño, Á.; Gómez-Gascón, L.; Luque, I.; Barrero-Domínguez, B.; Casamayor, A.; Cardoso-Toset, F.; Vela, A.I.; Fernández-Garayzábal, J.F.; Tarradas, C. Antimicrobial susceptibility and genetic characterization of Trueperella pyogenes isolates from pigs reared under intensive and extensive farming practices. Vet. Microbiol. 2019, 232, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Cameron, C.M.; Botha, W.F.; Smit, B.H.J. Antibody response to and immunity induced by Corynebacterium pyogenes vaccine. Onderstepoort J. Vet. Res. 1976, 43, 97–103. [Google Scholar] [PubMed]
- Derbyshire, J.B.; Matthews, P.R.J. Immunological Studies with Corynebacterium pyogenes in Mice. Res. Vet. Sci. 1963, 4, 537–542. [Google Scholar] [CrossRef]
- Hunter, P.; Van der Lugt, J.J.; Gouws, J.J. Failure of an Actinomyces pyogenes vaccine to protect sheep against an intravenous challenge. Onderstepoort J. Vet. Res. 1990, 57, 239–241. [Google Scholar]
- Desvaux, M.; Candela, T.; Serror, P. Surfaceome and Proteosurfaceome in Parietal Monoderm Bacteria: Focus on Protein Cell-Surface Display. Front. Microbiol. 2018, 9, 100. [Google Scholar] [CrossRef] [Green Version]
- Baums, C.G.; Valentin-Weigand, P. Surface-associated and secreted factors of Streptococcus suis in epidemiology, pathogenesis and vaccine development. Anim. Health Res. Rev. 2009, 10, 65–83. [Google Scholar] [CrossRef]
- Cordwell, S.J. Technologies for bacterial surface proteomics. Curr. Opin. Microbiol. 2006, 9, 320–329. [Google Scholar] [CrossRef]
- Navarre, W.W.; Schneewind, O. Surface proteins of gram-positive bacteria and mechanisms of their targeting to the cell wall envelope. Microbiol. Mol. Biol. Rev. 1999, 63, 174–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olaya-Abril, A.; Gómez-Gascón, L.; Jiménez-Munguía, I.; Obando, I.; Rodríguez-Ortega, M.J. Another turn of the screw in shaving Gram-positive bacteria: Optimization of proteomics surface protein identification in Streptococcus pneumoniae. J. Proteom. 2012, 75, 3733–3746. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gascón, L.; Luque, I.; Tarradas, C.; Olaya-Abril, A.; Astorga, R.J.; Huerta, B.; Rodríguez-Ortega, M.J. Comparative immunosecretome analysis of prevalent Streptococcus suis serotypes. Comp. Immunol. Microbiol. Infect. Dis. 2018, 57, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Merrill, C.; Ensermu, D.B.; Abdi, R.D.; Gillespie, B.E.; Vaughn, J.; Headrick, S.I.; Hash, K.; Walker, T.B.; Stone, E.; Kerro Dego, O. Immunological responses and evaluation of the protection in dairy cows vaccinated with staphylococcal surface proteins. Vet. Immunol. Immunopathol. 2019, 214, 109890. [Google Scholar] [CrossRef]
- Żakowska, D.; Graniak, G.; Rutyna, P.; Naylor, K.; Głowacka, P.; Niemcewicz, M. Protective antigen domain 4 of Bacillus anthracis as a candidate for use as vaccine for anthrax. Ann. Agric. Environ. Med. 2019, 26, 392–395. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Song, X.; Jing, J.; Zhao, K.; Shen, Y.; Zhang, X.; Yue, B. Chitosan-DNA nanoparticles enhanced the immunogenicity of multivalent DNA vaccination on mice against Trueperella pyogenes infection. J. Nanobiotechnol. 2018, 16, 8. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Zhang, W.; Bao, J.; Wu, Y.; Yan, M.; Xiao, Y.; Yang, L.; Zhang, Y.; Wang, J. A chimeric protein composed of the binding domains of Clostridium perfringens phospholipase C and Trueperella pyogenes pyolysin induces partial immunoprotection in a mouse model. Res. Vet. Sci. 2016, 107, 106–115. [Google Scholar] [CrossRef]
- Yang, L.; Liang, H.; Wang, B.; Ma, B.; Wang, J.; Zhang, W. Evaluation of the potency of two pyolysin-derived recombinant proteins as vaccine candidates of trueperella pyogenes in a mouse model: Pyolysin oligomerization and structural change affect the efficacy of pyolysin-based vaccines. Vaccines 2020, 8, 79. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Gascón, L.; Luque, I.; Olaya-Abril, A.; Jiménez-Munguía, I.; Orbegozo-Medina, R.A.; Peralbo, E.; Tarradas, C.; Rodríguez-Ortega, M.J. Exploring the pan-surfome of Streptococcus suis: Looking for common protein antigens. J. Proteom. 2012, 75, 5654–5666. [Google Scholar] [CrossRef]
- Rodríguez-Ortega, M.J. “Shaving” live bacterial cells with proteases for proteomic analysis of surface proteins. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2018; Volume 1722, pp. 21–29. [Google Scholar]
- Santos, T.; Viala, D.; Chambon, C.; Esbelin, J.; Hébraud, M. Listeria monocytogenes Biofilm Adaptation to Different Temperatures Seen through Shotgun Proteomics. Front. Nutr. 2019, 6, 89. [Google Scholar] [CrossRef]
- Espino, E.; Koskenniemi, K.; Mato-Rodriguez, L.; Nyman, T.A.; Reunanen, J.; Koponen, J.; Öhman, T.; Siljamäki, P.; Alatossava, T.; Varmanen, P.; et al. Uncovering surface-exposed antigens of Lactobacillus rhamnosus by cell shaving proteomics and two-dimensional immunoblotting. J. Proteome Res. 2015, 14, 1010–1024. [Google Scholar] [CrossRef] [PubMed]
- Feßler, A.T.; Schwarz, S. Antimicrobial Resistance in Corynebacterium spp., Arcanobacterium spp., and Trueperella pyogenes. Microbiol. Spectr. 2017, 5, 395–408. [Google Scholar] [CrossRef]
- van de Rijn, I.; Kessler, R.E. Growth characteristics of group A streptococci in a new chemically defined medium. Infect. Immun. 1980, 27, 444–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Angulo, V.A.; Kalita, A.; Kalita, M.; Lozano, L.; Torres, A.G. Comparative genomics and immunoinformatics approach for the identification of vaccine candidates for enterohemorrhagic Escherichia coli O157:H7. Infect. Immun. 2014, 82, 2016–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doytchinova, I.A.; Flower, D.R. VaxiJen: A server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinform. 2007, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Solis, N.; Cordwell, S.J. Current methodologies for proteomics of bacterial surface-exposed and cell envelope proteins. Proteomics 2011, 11, 3169–3189. [Google Scholar] [CrossRef]
- Grandi, G. Genomics and proteomics in reverse vaccines. Methods Biochem. Anal. 2006, 49, 379–393. [Google Scholar]
- Rodríguez-Ortega, M.J.; Luque, I.; Tarradas, C.; Bárcena, J.A. Overcoming function annotation errors in the Gram-positive pathogen Streptococcus suis by a proteomics-driven approach. BMC Genom. 2008, 9, 588. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Ortega, M.J.; Norais, N.; Bensi, G.; Liberatori, S.; Capo, S.; Mora, M.; Scarselli, M.; Doro, F.; Ferrari, G.; Garaguso, I.; et al. Characterization and identification of vaccine candidate proteins through analysis of the group A Streptococcus surface proteome. Nat. Biotechnol. 2006, 24, 191–197. [Google Scholar] [CrossRef]
- Bøhle, L.A.; Riaz, T.; Egge-Jacobsen, W.; Skaugen, M.; Busk, Ø.L.; Eijsink, V.G.H.; Mathiesen, G. Identification of surface proteins in Enterococcus faecalis V583. BMC Genom. 2011, 12, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olaya-Abril, A.; Jiménez-Munguía, I.; Gómez-Gascón, L.; Rodríguez-Ortega, M.J. Surfomics: Shaving live organisms for a fast proteomic identification of surface proteins. J Proteom. 2014, 97, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, W.J.; Davies, A.H.; Chambers, C.J.; Roberts, A.K.; Shone, C.C.; Acharya, K.R. Molecular features of the sortase enzyme family. FEBS J. 2015, 282, 2097–2114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spirig, T.; Weiner, E.M.; Clubb, R.T. Sortase enzymes in Gram-positive bacteria. Mol. Microbiol. 2011, 82, 1044–1059. [Google Scholar] [CrossRef] [PubMed]
- Susmitha, A.; Nampoothiri, K.M.; Bajaj, H. Insights into the biochemical and functional characterization of sortase E transpeptidase of Corynebacterium glutamicum. Biochem. J. 2019, 476, 3835–3847. [Google Scholar] [CrossRef]
- Das, S.; Pawale, V.S.; Dadireddy, V.; Singh, A.K.; Ramakumar, S.; Roy, R.P. Structure and specificity of a new class of Ca2+-independent housekeeping sortase from Streptomyces avermitilis provide insights into its non-canonical substrate preference. J. Biol. Chem. 2017, 292, 7244–7257. [Google Scholar] [CrossRef] [Green Version]
- Goudarzi, M.; Kobayashi, N.; Hashemi, A.; Fazeli, M.; Navidinia, M. Genetic variability of methicillin resistant staphylococcus aureus strains isolated from burns patients. Osong Public Heal. Res. Perspect. 2019, 10, 170–176. [Google Scholar] [CrossRef]
- Prados de la Torre, E.; Rodríguez-Franco, A.; Rodríguez-Ortega, M.J. Proteomic and bioinformatic analysis of streptococcus suis human isolates: Combined prediction of potential vaccine candidates. Vaccines 2020, 8, 188. [Google Scholar] [CrossRef] [Green Version]
- Olaya-Abril, A.; Jiménez-Munguía, I.; Gómez-Gascón, L.; Obando, I.; Rodríguez-Ortega, M.J. Identification of potential new protein vaccine candidates through pan-surfomic analysis of pneumococcal clinical isolates from adults. PLoS ONE 2013, 8, e70365. [Google Scholar] [CrossRef] [Green Version]
- Jost, B.H.; Trinh, H.T.; Songer, J.G.; Billington, S.J. Immunization with genetic toxoids of the Arcanobacterium pyogenes cholesterol-dependent cytolysin, pyolysin, protects mice against infection. Infect. Immun. 2003, 71, 2966–2969. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Wang, P.; Wang, B.; Ma, B.; Wang, J. A combined Clostridium perfringens/Trueperella pyogenes inactivated vaccine induces complete immunoprotection in a mouse model. Biologicals 2017, 47, 1–10. [Google Scholar] [CrossRef]
- Gómez-Gascón, L.; Cardoso-Toset, F.; Tarradas, C.; Gómez-Laguna, J.; Maldonado, A.; Nielsen, J.; Olaya-Abril, A.; Rodríguez-Ortega, M.J.; Luque, I. Characterization of the immune response and evaluation of the protective capacity of rSsnA against Streptococcus suis infection in pigs. Comp. Immunol. Microbiol. Infect. Dis. 2016, 47, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Zhao, K.; Zhang, Z.; Tang, C.; Zhang, X.; Yue, B. DNA vaccination based on pyolysin co-immunized with IL-1β enhances host antibacterial immunity against Trueperella pyogenes infection. Vaccine 2016, 34, 3469–3477. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Proteomics Reference | Origin a | Rearing System b | PFGE Pattern c | Cluster d |
|---|---|---|---|---|
| A | Liver | Extensive | 43 | B |
| B | Lung | Intensive | 28 | A |
| C | Lymph nodes | Intensive | 43 | B |
| D | Brain | Intensive | 53 | C |
| E | Lung | Intensive | 37 | B |
| F | Joint | Intensive | 37 | B |
| G | Lymph nodes | Extensive | 12 | A |
| H | Lymph nodes | Extensive | 1 | A |
| I | Liver | Extensive | 12 | A |
| J | Spleen | Extensive | 44 | B |
| K | Lymph nodes | Extensive | 43 | B |
| L | Heart | Intensive | 43 | B |
| M | Joint | Intensive | 13 | A |
| N | Heart | Intensive | 44 | B |
| O | Abscess | Extensive | 64 | C |
| Protein Category a | Identified Proteins among All the Strains | Range of Identified Proteins Per Strain |
|---|---|---|
| Surface proteins | 140 | 35–73 |
| Lipoprotein | 23 (16.4%) | 1–13 |
| Cell Wall | 25 (17.9%) | 13–25 |
| Secretory | 10 (7.1%) | 5–10 |
| Membrane | 82 (58.6%) | 22–50 |
| Ranking | Proteins |
|---|---|
| A (16) a | Cell wall: X4QWN2, X4R8M3 Lipoproteins: A0A0M3SNR1, A0A0M4K9G4, A0A0M4JY33 Secreted proteins: X4R0V4, A0A2G9KEL5, Q9S0W7, X4QUK6 Membrane proteins (1 TMD): A0A0M4K7E7, A0A2G9KDB2, X4RDW5 Membrane proteins (>1 TMD): A0A0M3SNZ9, A0A0M4KS30, A0A2G9KB86, A0A2G9KCY0, A0A0M4JWL1 |
| B (9) | Cell wall: A0A2G9KC18, A0A2G9KBF5 Lipoproteins: A0A0M5KH79, A0A0M4KR83 Secreted proteins: A0A2G9KD79, A0A2G9KA87 Membrane proteins (>1 TMD): X4QZN5, A0A2G9KBU5, A0A0M3SNG2 |
| C (15) | Cell wall: X4RE32, X4RB57, A0A0M5KIG4, X4RCL8, X4QMI5, A0A2G9KAD4, A0A0M4K5P6 Lipoproteins: A0A0M5KPJ2, A0A0M4K9R6, X4QXA0, A0A2G9KEH2, A0A0M4K5B8 Secreted proteins: A0A0M4KB23, A0A0M4JYB5 Membrane proteins (1 TMD): A0A2G9KB80, A0A0M3SN42, A0A2G9KB66, A0A0M3SNB2 Membrane proteins (>1 TMD): X4QR96, A0A0M3SNU6, A0A2G9KB07, X4RC16 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galán-Relaño, Á.; Gómez-Gascón, L.; Rodríguez-Franco, A.; Luque, I.; Huerta, B.; Tarradas, C.; Rodríguez-Ortega, M.J. Search of Potential Vaccine Candidates against Trueperella pyogenes Infections through Proteomic and Bioinformatic Analysis. Vaccines 2020, 8, 314. https://doi.org/10.3390/vaccines8020314
Galán-Relaño Á, Gómez-Gascón L, Rodríguez-Franco A, Luque I, Huerta B, Tarradas C, Rodríguez-Ortega MJ. Search of Potential Vaccine Candidates against Trueperella pyogenes Infections through Proteomic and Bioinformatic Analysis. Vaccines. 2020; 8(2):314. https://doi.org/10.3390/vaccines8020314
Chicago/Turabian StyleGalán-Relaño, Ángela, Lidia Gómez-Gascón, Antonio Rodríguez-Franco, Inmaculada Luque, Belén Huerta, Carmen Tarradas, and Manuel J. Rodríguez-Ortega. 2020. "Search of Potential Vaccine Candidates against Trueperella pyogenes Infections through Proteomic and Bioinformatic Analysis" Vaccines 8, no. 2: 314. https://doi.org/10.3390/vaccines8020314
APA StyleGalán-Relaño, Á., Gómez-Gascón, L., Rodríguez-Franco, A., Luque, I., Huerta, B., Tarradas, C., & Rodríguez-Ortega, M. J. (2020). Search of Potential Vaccine Candidates against Trueperella pyogenes Infections through Proteomic and Bioinformatic Analysis. Vaccines, 8(2), 314. https://doi.org/10.3390/vaccines8020314