Characterization of the Burkholderia cenocepacia J2315 Surface-Exposed Immunoproteome

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
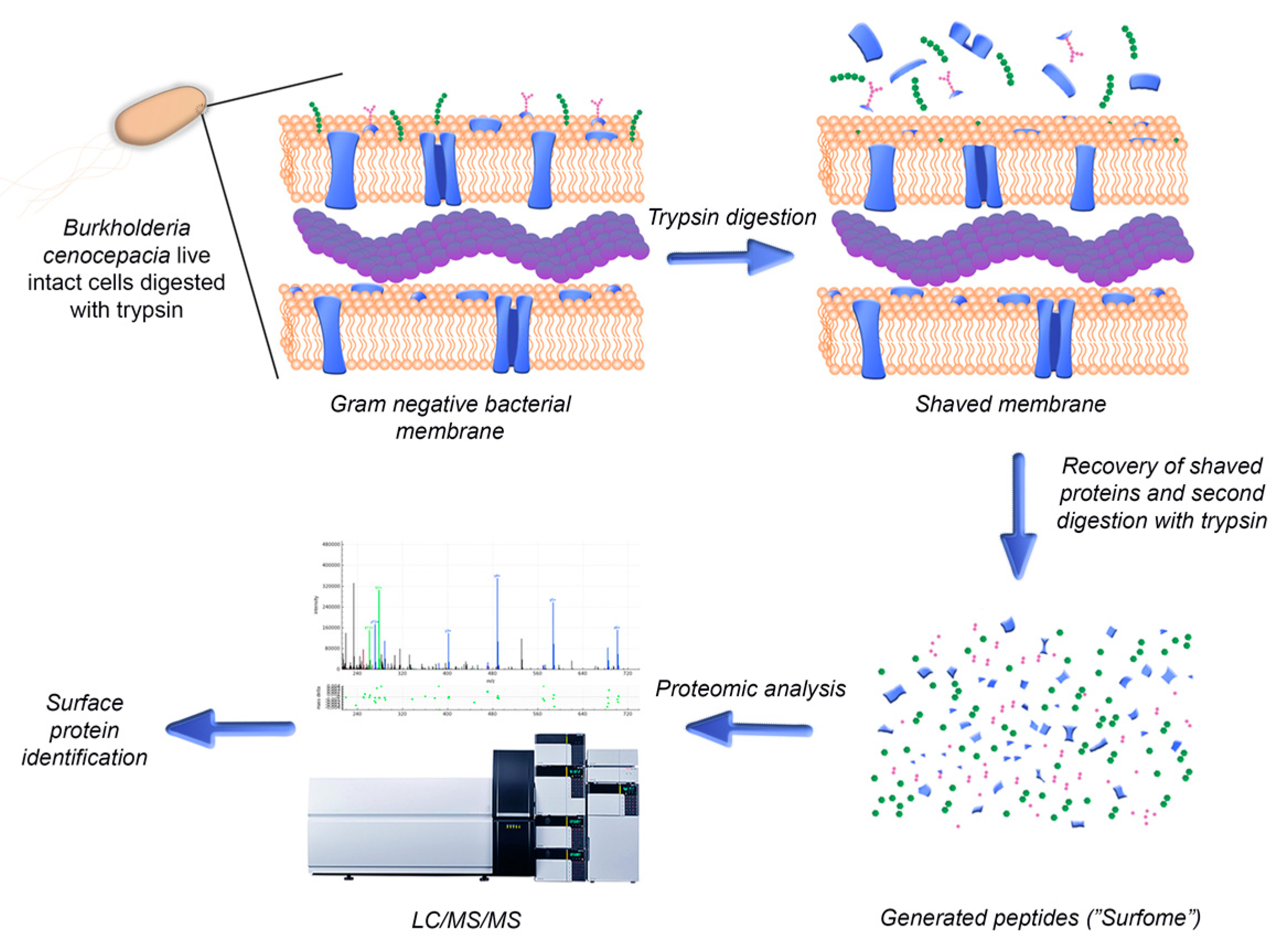
2.2. Digestion of Live B. cenocepacia Cells’ Surface with Trypsin
2.3. Bacterial Cell Viability Assay
2.4. Cleaning of Peptides with Solid-Phase Extraction Cartridges
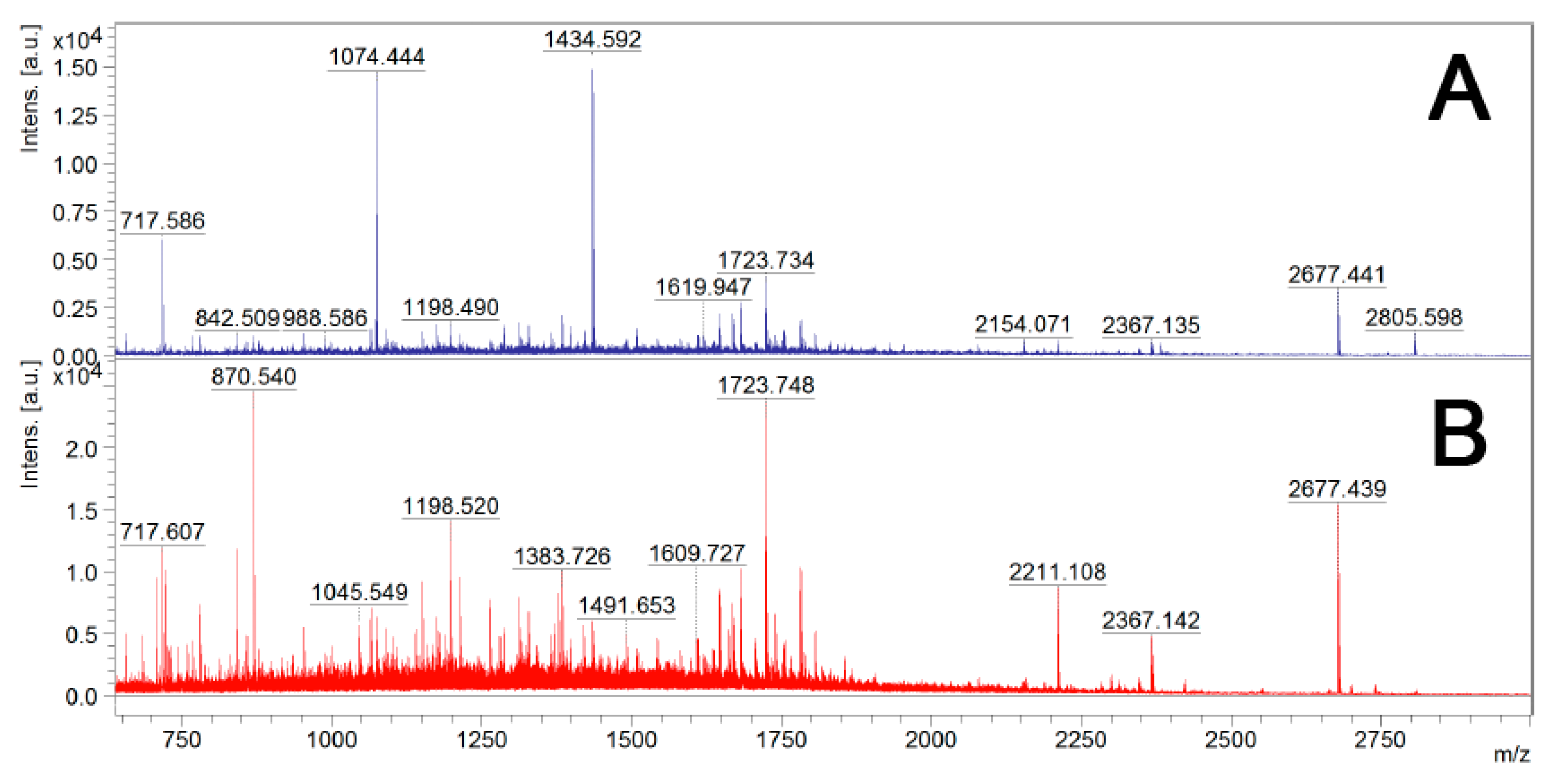
2.5. MALDI-TOF MS Analysis
2.6. LC-MS/MS Analysis
2.7. Protein Identification by Database Searching
2.8. Molecular Biology Techniques
2.9. Cloning and Overexpression of B. cenocepacia J2315 bcal2022, bcal2645 and bcal2958 Proteins
2.10. Purification of B. cenocepacia J2315 His-Tagged Proteins BCAL2022, BCAL2645 and BCAL2958
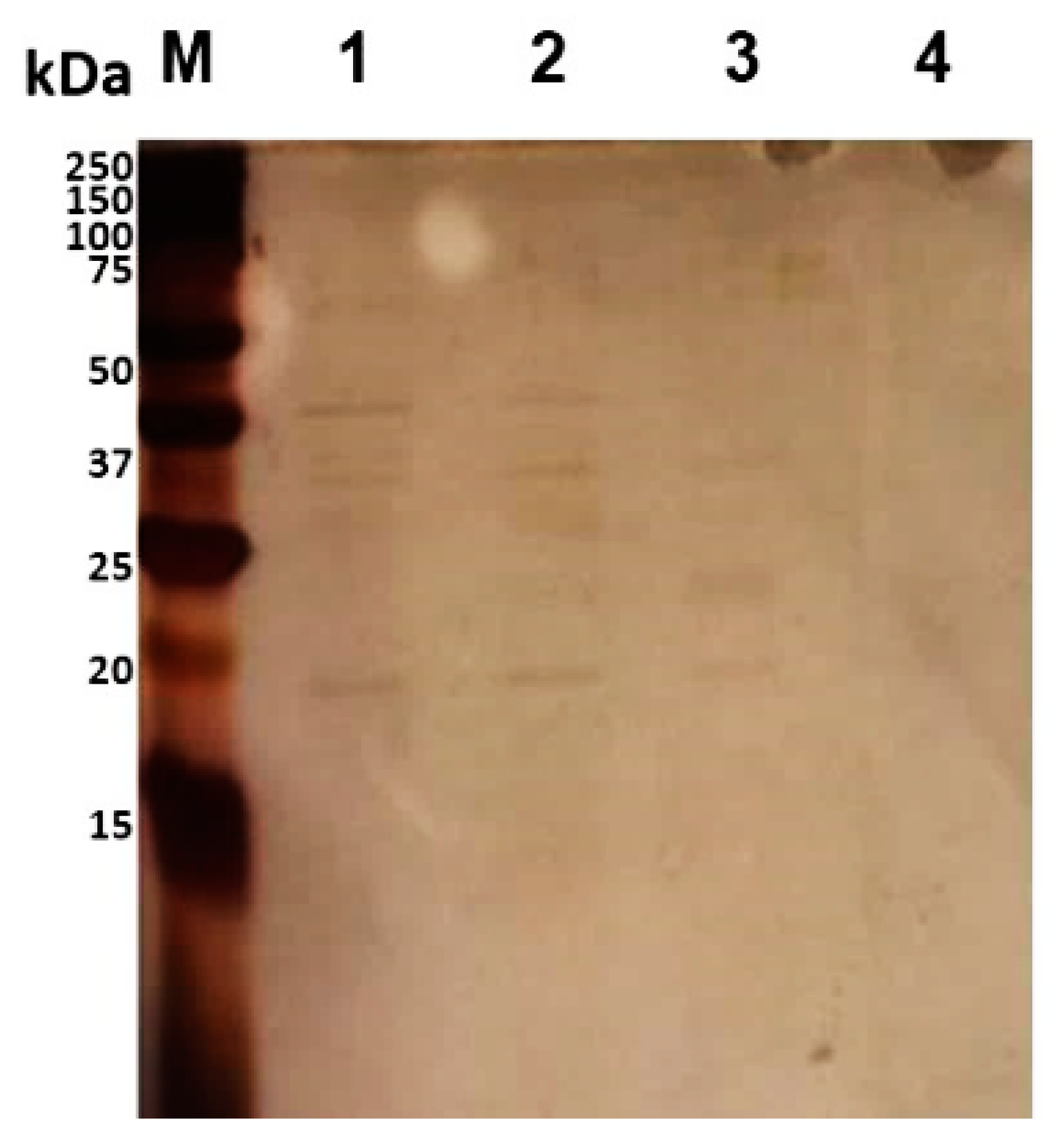
2.11. CF Patients’ Blood Sera’s Immunoreactivity against the Proteins BCAL2022, BCAL2645 and BCAL2958
2.12. Enzyme-Linked Immunosorbent Assay (ELISA)
2.13. Bioinformatics Analysis
3. Results
3.1. Identification and Analysis of Surface-Exposed Proteins of B. cenocepacia J2315 with Immunogenic Epitopes Using Bioinformatics Tools
3.2. Characterization of Surface-Exposed Proteins Using Trypsin Digestion of Live B. cenocepacia J2315 Cells
3.2.1. Optimization of “Shaving” Protocol
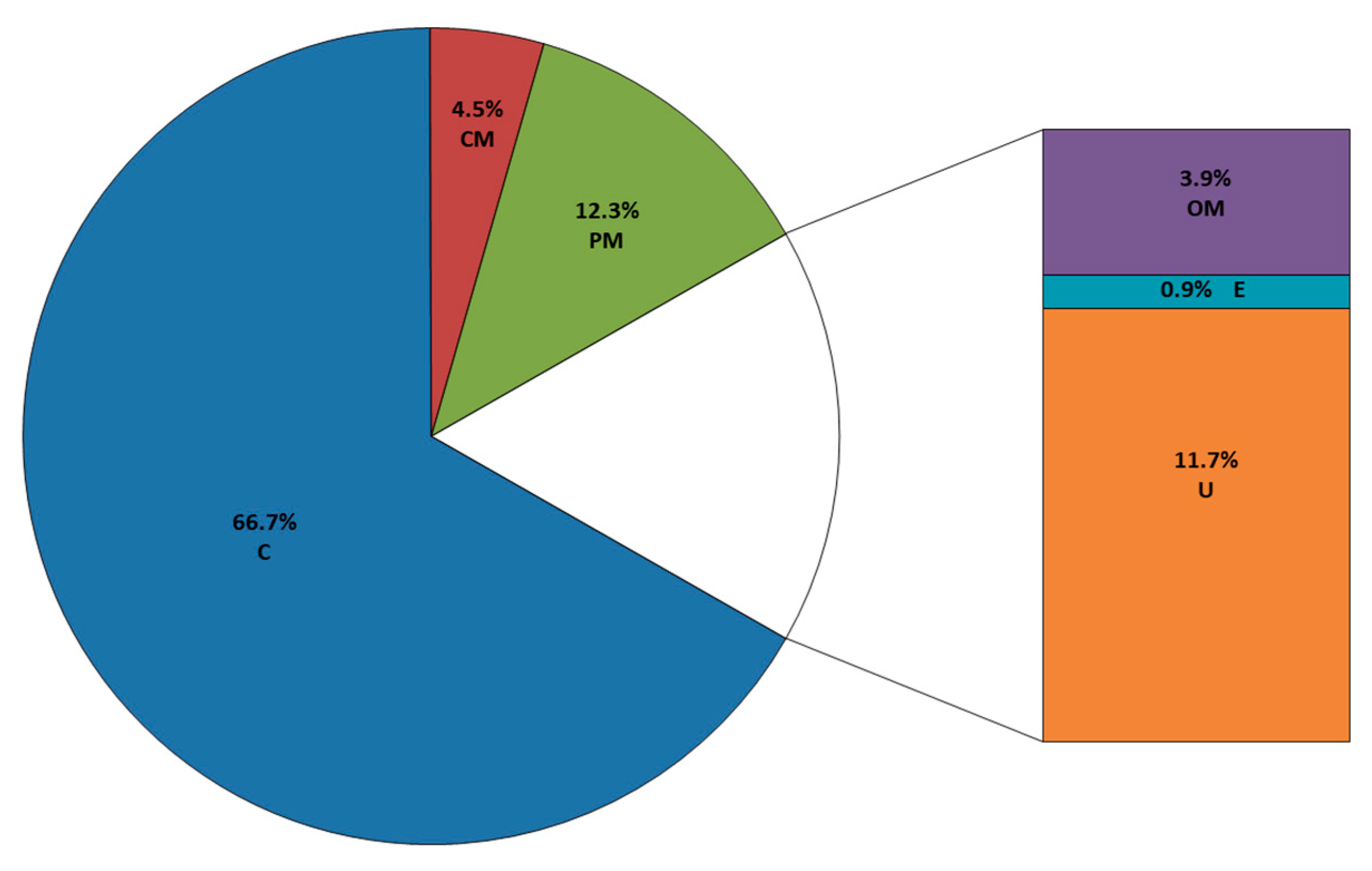
3.2.2. Identification of Surface-Exposed Proteins Using Trypsin Digestion of Live B. cenocepacia J2315 Cells
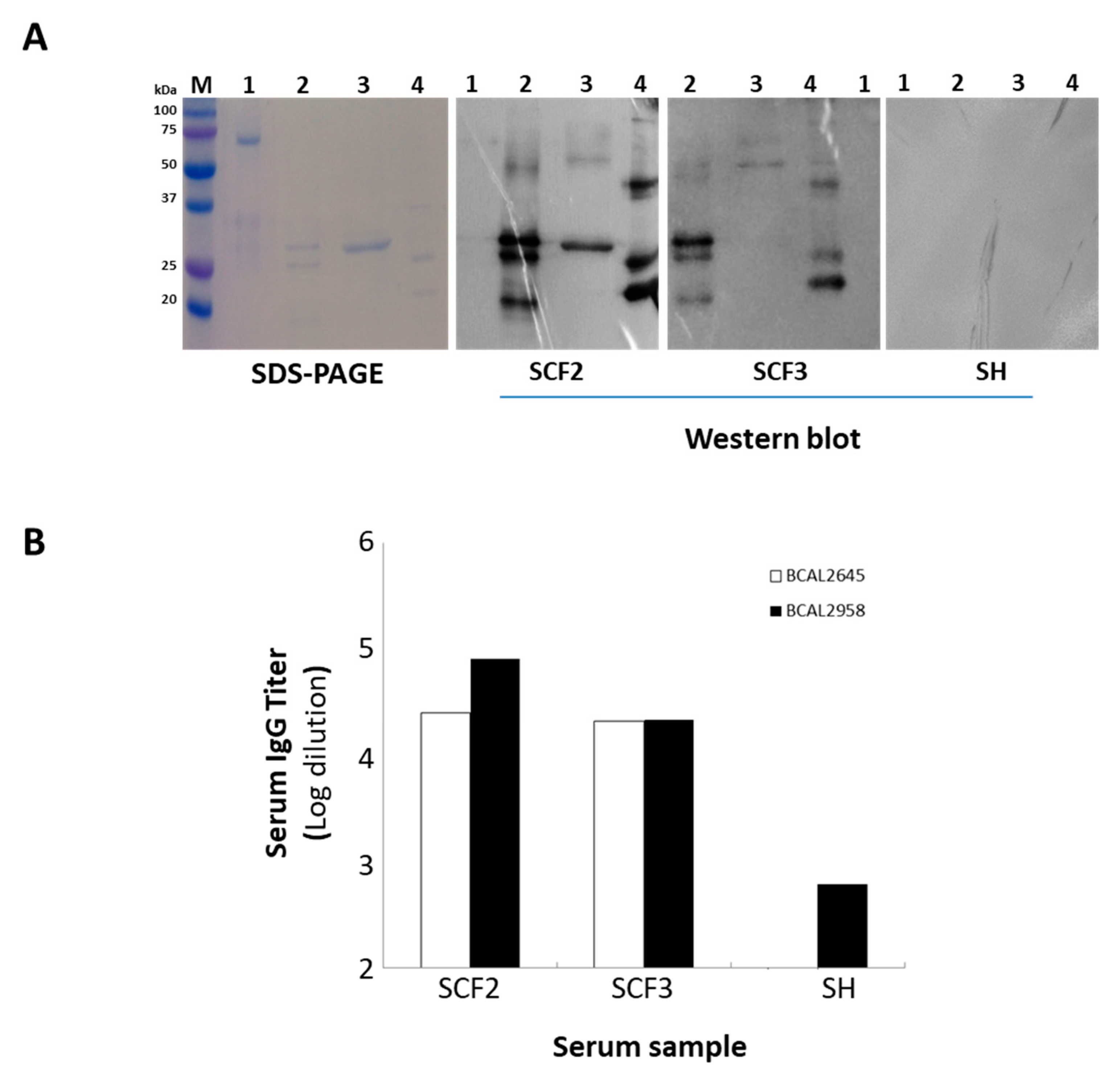
3.3. Characterization of B. cenocepacia bcal2022 and bcal2645 Proteins’ Immunoreactivity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Smet, B.; Mayo, M.; Peeters, C.; Zlosnik, J.E.A.; Spilker, T.; Hird, T.J.; Li Puma, J.J.; Kidd, T.J.; Kaestli, M.; Ginther, J.L.; et al. Burkholderia stagnalis sp. nov. and Burkholderia territorii sp. nov., two novel Burkholderia cepacia complex species from environmental and human sources. Int. J. Syst. Evol. Microbiol. 2015, 65, 2265–2271. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.F.; King, G.M. Volcanic soils as sources of novel CO-oxidizing Paraburkholderia and Burkholderia: Paraburkholderia hiiakae sp. nov., Paraburkholderia metrosideri sp. nov., Paraburkholderia paradisi sp. nov., Paraburkholderia peleae. Front. Microbiol. 2017, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martina, P.; Leguizamon, M.; Prieto, C.I.; Sousa, S.A.; Montanaro, P.; Draghi, W.O.; Stämmler, M.; Bettiol, M.; de Carvalho, C.C.C.R.; Palau, J.; et al. Burkholderia puraquae sp. nov., a novel species of the Burkholderia cepacia complex isolated from hospital settings and agricultural soils. Int. J. Syst. Evol. Microbiol. 2018, 68, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Tavares, M.; Kozak, M.; Balola, A.; Sá-Correia, I. Burkholderia cepacia complex bacteria: A feared contamination risk in water-based pharmaceutical products. Clin. Microbiol. Rev. 2020, 33, e00139-19. [Google Scholar] [CrossRef] [PubMed]
- Leitão, J.H.; Sousa, S.A.; Ferreira, A.S.; Ramos, C.G.; Silva, I.N.; Moreira, L.M. Pathogenicity, virulence factors, and strategies to fight against Burkholderia cepacia complex pathogens and related species. Appl. Microbiol. Biotechnol. 2010, 87, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Isles, A.; Maclusky, I.; Corey, M.; Gold, R.; Prober, C.; Fleming, P.; Levison, H. Pseudomonas cepacia infection in cystic fibrosis: An emerging problem. J. Pediatr. 1984, 104, 206–210. [Google Scholar] [CrossRef]
- Leitão, J.H.; Sousa, S.A.; Cunha, M.V.; Salgado, M.J.; Melo-Cristino, J.; Barreto, M.C.; Sá-Correia, I. Variation of the antimicrobial susceptibility profiles of Burkholderia cepacia complex clonal isolates obtained from chronically infected cystic fibrosis patients: A five-year survey in the major Portuguese treatment center. Eur. J. Clin. Microbiol. Infect. Dis. 2008, 27, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- Scoffone, V.C.; Chiarelli, L.R.; Trespidi, G.; Mentasti, M.; Riccardi, G.; Buroni, S. Burkholderia cenocepacia infections in cystic fibrosis patients: Drug resistance and therapeutic approaches. Front. Microbiol. 2017, 8, 1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, S.A.; Feliciano, J.R.; Pita, T.; Guerreiro, S.I.; Leitão, J.H. Burkholderia cepacia Complex Regulation of Virulence Gene Expression: A Review. Genes 2017, 8, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B. Lung Infections Associated with Cystic Fibrosis Lung Infections Associated with Cystic Fibrosis. Clin. Microbiol. Rev. 2002, 15, 194–222. [Google Scholar] [CrossRef] [Green Version]
- Regan, K.; Jayesh, B. Eradication therapy for Burkholderia cepacia complex in people with cystic fibrosis. Cochrane Database Syst. Rev. 2016, 11, CD009876. [Google Scholar] [CrossRef] [PubMed]
- Sousa, S.; Seixas, A.; Leitão, J. Postgenomic Approaches and Bioinformatics Tools to Advance the Development of Vaccines against Bacteria of the Burkholderia cepacia Complex. Vaccines 2018, 6, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scoffone, V.C.; Barbieri, G.; Buroni, S.; Scarselli, M.; Pizza, M.; Rappuoli, R.; Riccardi, G. Vaccines to Overcome Antibiotic Resistance: The Challenge of Burkholderia cenocepacia. Trends Microbiol. 2020, 28, 315–326. [Google Scholar] [CrossRef] [PubMed]
- McClean, S.; Healy, M.E.; Collins, C.; Carberry, S.; O’Shaughnessy, L.; Dennehy, R.; Adams, Á.; Kennelly, H.; Corbett, J.M.; Carty, F.; et al. Linocin and OmpW Are Involved in Attachment of the Cystic Fibrosis-Associated Pathogen Burkholderia cepacia Complex to Lung Epithelial Cells and Protect Mice against Infection. Infect. Immun. 2016, 84, 1424–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradenas, G.; Ross, B.; Torres, A. Burkholderia cepacia Complex Vaccines: Where Do We Go from here? Vaccines 2016, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Grandi, G. Antibacterial vaccine design using genomics and proteomics. Trends Biotechnol. 2001, 19, 181–188. [Google Scholar] [CrossRef]
- Olaya-Abril, A.; Jiménez-Munguía, I.; Gómez-Gascón, L.; Rodríguez-Ortega, M.J. Surfomics: Shaving live organisms for a fast proteomic identification of surface proteins. J. Proteomics 2014, 97, 164–176. [Google Scholar] [CrossRef]
- Rabilloud, T. Membrane proteins and proteomics: Love is possible, but so difficult. Electrophoresis 2009, 30, 174–180. [Google Scholar] [CrossRef] [Green Version]
- Sousa, S.A.; Soares-Castro, P.; Seixas, A.M.M.; Feliciano, J.R.; Balugas, B.; Barreto, C.; Pereira, L.; Santos, P.M.; Leitão, J.H. New insights into the immunoproteome of B. cenocepacia J2315 using serum samples from cystic fibrosis patients. N. Biotechnol. 2020, 54, 62–70. [Google Scholar] [CrossRef]
- Walters, M.S.; Mobley, H.L.T. Identification of uropathogenic Escherichia coli surface proteins by shotgun proteomics. J. Microbiol. Methods 2009, 78, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Braconi, D.; Amato, L.; Bernardini, G.; Arena, S.; Orlandini, M.; Scaloni, A.; Santucci, A. Surfome analysis of a wild-type wine Saccharomyces cerevisiae strain. Food Microbiol. 2011, 28, 1220–1230. [Google Scholar] [CrossRef] [PubMed]
- Castro-Borges, W.; Dowle, A.; Curwen, R.S.; Thomas-Oates, J.; Wilson, R.A. Enzymatic shaving of the tegument surface of live schistosomes for proteomic analysis: A rational approach to select vaccine candidates. PLoS Negl. Trop. Dis. 2011, 5, e993. [Google Scholar] [CrossRef] [PubMed]
- Worlitzsch, D.; Tarran, R.; Ulrich, M.; Schwab, U.; Cekici, A.; Meyer, K.C.; Birrer, P.; Bellon, G.; Berger, J.; Weiss, T.; et al. Effects of reduced mucus oxygen concentration in airway Pseudomonas infections of cystic fibrosis patients. J. Clin. Investig. 2002, 109, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Sass, A.M.; Schmerk, C.; Agnoli, K.; Norville, P.J.; Eberl, L.; Valvano, M.A.; Mahenthiralingam, E. The unexpected discovery of a novel low-oxygen-activated locus for the anoxic persistence of Burkholderia cenocepacia. ISME J. 2013, 14, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govan, J.R.; Deretic, V. Microbial pathogenesis in cystic fibrosis: Mucoid Pseudomonas aeruginosa and Burkholderia cepacia. Microbiol. Rev. 1996, 60, 539–574. [Google Scholar] [CrossRef] [PubMed]
- Sousa, S.A.; Morad, M.; Feliciano, J.R.; Pita, T.; Nady, S.; El-Hennamy, R.E.; Abdel-Rahman, M.; Cavaco, J.; Pereira, L.; Barreto, C.; et al. The Burkholderia cenocepacia OmpA-like protein BCAL2958: Identification, characterization, and detection of anti-BCAL2958 antibodies in serum from B. cepacia complex-infected Cystic Fibrosis patients. AMB Express 2016, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Ortega, M.J. ‘Shaving’ Live Bacterial Cells with Proteases for Proteomic Analysis of Surface Proteins. In Methods in Molecular Biology (Clifton, N.J.); Humana Press: Totowa, NJ, USA, 2018; Volume 1722, pp. 21–29. [Google Scholar]
- Sriramulu, D.D.; Lünsdorf, H.; Lam, J.S.; Römling, U. Microcolony formation: A novel biofilm model of Pseudomonas aeruginosa for the cystic fibrosis lung. J. Med. Microbiol. 2005, 54, 667–676. [Google Scholar] [CrossRef]
- Chevallet, M.; Luche, S.; Rabilloud, T. Silver staining of proteins in polyacrylamide gels. Nat. Protoc. 2006, 1, 1852–1858. [Google Scholar] [CrossRef] [Green Version]
- Winsor, G.L.; Khaira, B.; Van Rossum, T.; Lo, R.; Whiteside, M.D.; Brinkman, F.S.L. The Burkholderia Genome Database: Facilitating flexible queries and comparative analyses. Bioinformatics 2008, 24, 2803–2804. [Google Scholar] [CrossRef] [Green Version]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Yu, N.Y.; Wagner, J.R.; Laird, M.R.; Melli, G.; Rey, S.; Lo, R.; Dao, P.; Sahinalp, S.C.; Ester, M.; Foster, L.J.; et al. PSORTb 3.0: Improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 2010, 26, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2012. [Google Scholar]
- Burkholderia cenocepacia—Wellcome Sanger Institute. Available online: https://www.sanger.ac.uk/resources/downloads/bacteria/burkholderia-cenocepacia.html (accessed on 30 July 2020).
- Jespersen, M.C.; Peters, B.; Nielsen, M.; Marcatili, P. BepiPred-2.0: Improving sequence-based B-cell epitope prediction using conformational epitopes. Nucleic Acids Res. 2017, 45, W24–W29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharon, J.; Rynkiewicz, M.J.; Lu, Z.; Yang, C.-Y. Discovery of protective B-cell epitopes for development of antimicrobial vaccines and antibody therapeutics. Immunology 2014, 142, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Olaya-Abril, A.; Gómez-Gascón, L.; Jiménez-Munguía, I.; Obando, I.; Rodríguez-Ortega, M.J. Another turn of the screw in shaving Gram-positive bacteria: Optimization of proteomics surface protein identification in Streptococcus pneumoniae. J. Proteomics 2012, 75, 3733–3746. [Google Scholar] [CrossRef] [PubMed]
- Higdon, R.; Kolker, E. A predictive model for identifying proteins by a single peptide match. Bioinformatics 2007, 23, 277–280. [Google Scholar] [CrossRef]
- Mani, M.; Chen, C.; Amblee, V.; Liu, H.; Mathur, T.; Zwicke, G.; Zabad, S.; Patel, B.; Thakkar, J.; Jeffery, C.J. MoonProt: A database for proteins that are known to moonlight. Nucleic Acids Res. 2014, 43, 277–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Burkholderia Genome Database. Available online: https://www.burkholderia.com/ (accessed on 30 July 2020).
- Pfam Database. Available online: http://pfam.xfam.org/ (accessed on 30 July 2020).
- BepiPred 2.0 Server. Available online: http://tools.iedb.org/bcell/ (accessed on 30 July 2020).
- Holden, M.T.G.; Seth-Smith, H.M.B.; Crossman, L.C.; Sebaihia, M.; Bentley, S.D.; Cerdeno-Tarraga, A.M.; Thomson, N.R.; Bason, N.; Quail, M.A.; Sharp, S.; et al. The genome of Burkholderia cenocepacia J2315, an epidemic pathogen of Cystic Fibrosis patients. J. Bacteriol. 2009, 191, 261–277. [Google Scholar] [CrossRef] [Green Version]
- Spencer, H.K.; Spitznogle, S.L.; Borjan, J.; Aitken, S.L. An Overview of the Treatment of Less Common Non-Lactose Fermenting Gram Negative Bacteria. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2020. accepted. [Google Scholar] [CrossRef]
- Pradenas, G.; Myers, J.; Torres, A. Characterization of the Burkholderia cenocepacia TonB Mutant as a Potential Live Attenuated Vaccine. Vaccines 2017, 5, 33. [Google Scholar] [CrossRef]
- Musson, J.A.; Reynolds, C.J.; Rinchai, D.; Nithichanon, A.; Khaenam, P.; Favry, E.; Spink, N.; Chu, K.K.Y.; De Soyza, A.; Bancroft, G.J.; et al. CD4 + T Cell Epitopes of FliC Conserved between Strains of Burkholderia: Implications for Vaccines against Melioidosis and Cepacia Complex in Cystic Fibrosis. J. Immunol. 2014, 193, 6041–6049. [Google Scholar] [CrossRef] [Green Version]
- Sousa, S.A.; Feliciano, J.R.; Grilo, A.M.; Leitão, J.H. Bioinformatics: A Molecular Microbiologist’s Perspective. Curr. Bioinform. 2014, 9, 8–17. [Google Scholar] [CrossRef]
- Liu, H.; Ibrahim, M.; Qiu, H.; Kausar, S.; Ilyas, M.; Cui, Z.; Hussain, A.; Li, B.; Waheed, A.; Zhu, B.; et al. Protein Profiling Analyses of the Outer Membrane of Burkholderia cenocepacia Reveal a Niche-Specific Proteome. Microb. Ecol. 2015, 69, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Wolden, R.; Pain, M.; Karlsson, R.; Karlsson, A.; Aarag Fredheim, E.G.; Cavanagh, J.P. Identification of surface proteins in a clinical Staphylococcus haemolyticus isolate by bacterial surface shaving. BMC Microbiol. 2020, 20, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solis, N.; Cordwell, S.J. Cell shaving and False-Positive control strategies coupled to novel statistical tools to profile Gram-Positive bacterial surface proteomes. In Methods in Molecular Biology; Humana Press Inc: New York, NY, USA, 2016; Volume 1440, pp. 47–55. [Google Scholar]
- Henderson, B.; Martin, A. Bacterial virulence in the moonlight: Multitasking bacterial moonlighting proteins are virulence determinants in infectious disease. Infect. Immun. 2011, 79, 3476–3491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pore, D.; Chakrabarti, M.K. Outer membrane protein A (OmpA) from Shigella flexneri 2a: A promising subunit vaccine candidate. Vaccine 2013, 31, 3644–3650. [Google Scholar] [CrossRef] [PubMed]
- Confer, A.W.; Ayalew, S. The OmpA family of proteins: Roles in bacterial pathogenesis and immunity. Vet. Microbiol. 2013, 163, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Darwin, A.J. Stress relief during host infection: The phage shock protein response supports bacterial virulence in various ways. PLoS Pathog. 2013, 9, e1003388. [Google Scholar] [CrossRef]
- Southern, S.J.; Male, A.; Milne, T.; Sarkar-Tyson, M.; Tavassoli, A.; Oyston, P.C.F. Evaluating the role of phage-shock protein A in Burkholderia pseudomallei. Microbiology 2015, 161, 2192–2203. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Genotype or Description | Source or Reference |
|---|---|---|
| B. cenocepacia J2315 | Cystic Fibrosis isolate; ET12 lineage reference strain, genome sequence available. | [25] |
| E. coli DH5α | F− Φ80lacZΔM15 Δ(lacZYA-argF) U169 recA1 endA1 hsdR17(rk−, mk+) phoA supE44 thi-1 gyrA96 relA1 λ− | Invitrogen (Carlsbad, CA, USA) |
| E. coli BL21 (DE3) | F− ompT hsdSB (rB−mB−) dcm gal λ(DE3) | Stratagene (San Diego, CA, USA) |
| Plasmids | ||
| pET23a+ | Cloning/expression vector, T7 promoter, C-terminal 6x His-Tag, Apr | Novagen (Madison, WI, USA) |
| pET29a+ | Cloning/expression vector, T7 promoter, thrombin recognition site, C-terminal 6’ His-Tag, Kanr | Novagen |
| pSAS38 | pET23a+ containing the pET29a+ thrombin recognition site, cloned downstream of the T7 promoter and upstream of the C-terminal 6× His-Tag, Apr | This study |
| pMM1 | pSAS38 containing the BCAL2022 gene, cloned downstream of the T7 promoter, Apr | This study |
| pSAS36 | pET23a+ containing the BCAL2645 gene, cloned downstream of the T7 promoter and upstream of the C-terminal 6× His-Tag, Apr | This study |
| pSAS6 | pET23a+ containing the BCAL2958 gene, cloned downstream of the T7 promoter and upstream of the C-terminal 6× His-Tag, Apr | [26] |
| ORF 1 | Description 1 | Peptides Identified (PSMs) 2 | Subcellular Localization 3 | Domains 4 | B-Cell Epitope Average (Predicted Peptides) 5 |
|---|---|---|---|---|---|
| BCAL1416 (AlaS) | Alanyl-tRNA synthetase | 23 (28) | U (Class 3) | tRNA synthetases class II (A) (PF01411) Threonyl and Alanyl tRNA synthetase second additional domain (PF07973) DHHA1 domain (PF02272) | 0.464 (23) |
| BCAM0965 (mdh) | Malate dehydrogenase | 17 (29) | U (Class 3) | Lactate/malate dehydrogenase, NAD binding domain (PF00056) Lactate/malate dehydrogenase, alpha/beta C-terminal domain (PF02866) | 0.430 (6) |
| BCAL2736 | Isocitrate dehydrogenase | 17 (25) | U (Class 3) | Isocitrate/isopropylmalate dehydrogenase (PF00180) | 0.460 (8) |
| BCAL3203 (TolB) | Translocation protein TolB | 17 (22) | U (Class 3) | TolB amino-terminal domain (PF04052) WD40-like Beta Propeller Repeat (PF07676) | 0.454 (9) |
| BCAL2777 | Putative N-acetylmuramoyl-L-alanine amidase | 14 (15) | U (Class 3) | AMIN domain (PF11741) N-acetylmuramoyl-L-alanine amidase (PF01520) | 0.511 (12) |
| BCAL2956 | Hypothetical protein | 13 (32) | U (Class 3) | DUF2059 (PF09832) | 0.485 (6) |
| BCAL2993 (PepN) | Aminopeptidase N | 13 (14) | U (Class 3) | Peptidase M1 N-terminal domain (PF17900) Peptidase family M1 domain (PF01433) DUF3458, Ig-like fold (PF11940) DUF3458_C, ARM repeats (PF17432) | 0.456 (24) |
| BCAL3311 (BcnA) | Secreted bacterial lipocalin | 12 (22) | E (Class 1) | YceI-like domain (PF04264) | 0.455 (3) |
| BCAL1262 (CarB) | Carbamoyl phosphate synthase large subunit | 12 (13) | U (Class 3) | Carbamoyl-phosphate synthase L chain: ATP binding domain (PF02786); Oligomerization domain (PF02787) MGS-like domain (PF02142) | 0.457 (25) |
| BCAM1931 | Putative porin | 11 (15) | OM (Class 3) | Porin_4 (PF13609) | 0.503 (10) |
| BCAL0765 | Hypothetical protein | 11 (12) | U (Class 3) | ABC transporter substrate-binding protein (PF04392) | 0.454 (5) |
| BCAL1961 | Hypothetical protein | 10 (20) | U (Class 3) | Ankyrin repeats (PF12796) | 0.472 (6) |
| BCAM0906 | Putative dienelactone hydrolase family protein | 10 (14) | U (Class 3) | Dienelactone hydrolase family (PF01738) | 0.471 (7) |
| BCAL2206 (PhaP) | Phasin-like protein | 10 (13) | U (Class 3) | Phasin_2 protein (PF09361) | 0.482 (5) |
| BCAM0043 | Hypothetical protein | 10 (12) | U (Class 3) | Phage late control gene D protein (PF05954) T6SS_Vgr (PF13296) DUF2345 (PF10106) | 0.494 (12) |
| BCAM1576 | Phosphoesterase family protein | 10 (10) | E (Class 3) | Phosphoesterase family (PF04185) | 0.489 (10) |
| BCAL0340 (BscM) | Putative lipoprotein. Part of the T6SS gene cluster | 9 (13) | U (Class 3) | Tetratricopeptide repeat (PF14559) | 0.506 (6) |
| BCAL1493 | Putative exported protein | 9 (12) | U (Class 3) | NlpB/DapX lipoprotein (PF06804) | 0.518 (6) |
| BCAL2166 | Putative lipoprotein | 9 (10) | U (Class 3) | NlpB/DapX lipoprotein (PF06804) | 0.500 (10) |
| BCAL0305 | Hypothetical protein | 8 (21) | U (Class 3) | MlaC protein (PF05494) | 0.480 (7) |
| BCAL1893 | Family M23 peptidase | 8 (13) | OM (Class 3) | LysM domain (PF014769) Peptidase family M23 (PF01551) | 0.523 (4) |
| BCAL0151 | Putative lipoprotein | 8 (12) | U (Class 3) | Periplasmic binding protein (PF13458) | 0.439 (10) |
| BCAL2934 (EtfA) | Electron transfer flavoprotein alpha-subunit | 8 (10) | U (Class 3) | Electron transfer flavoprotein domain (PF01012) Electron transfer flavoprotein FAD-binding domain (PF00766) | 0.443 (3) |
| BCAM1012 | Putative histone-like protein | 8 (10) | U (Class 3) | Bacterial DNA-binding protein (PF00216) | 0.520 (2) |
| BCAL2958 | Putative OmpA family protein | 7 (25) | OM (Class 3) | OmpA family (PF00691) | 0.512 (3) |
| BCAM2827 | Hypothetical protein | 7 (13) | U (Class 3) | MlaC protein (PF05494) | 0.462 (4) |
| BCAS0667 | Uncharacterized protein | 7 (9) | U (Class 3) | Phage late control gene D protein (PF05954) | 0.500 (20) |
| BCAL0020 | Putative branched-chain amino acid ABC transporter periplasmic substrate binding protein | 7 (9) | U (Class 3) | Periplasmic binding protein (PF13458) | 0.442 (8) |
| BCAS0219 | Hypothetical protein | 7 (8) | U (Class 3) | Outer membrane lipoprotein-sorting protein (PF17131) | 0.461 (7) |
| BCAM2736 | Hypothetical protein | 7 (8) | U (Class 3) | none | 0.534 (0) |
| BCAM1712 (PaaH) | 3-hydroxy-acyl-CoA dehydrogenase | 7 (8) | U (Class 3) | 3-hydroxyacyl-CoA dehydrogenase: NAD binding domain (PF02737); C-terminal domain (PF00725) 3-hydroxybutyryl-CoA dehydrogenase reduced Rossmann-fold domain (PF18321) | 0.443 (7) |
| BCAL2687 | Putative lipoprotein | 7 (7) | U (Class 3) | Periplasmic binding protein (PF13458) | 0.438 (7) |
| BCAL2946 (udg) | UDP-glucose dehydrogenase | 7 (7) | U (Class 3) | UDP-glucose/GDP-mannose dehydrogenase family: NAD binding domain (PF03721); Central domain (PF00984); UDP binding domain (PF03720) | 0.447 (11) |
| BCAL2828 | Hypothetical protein | 6 (11) | U (Class 3) | Carboxypeptidase regulatory-like domain (PF13620) | 0.468 (3) |
| BCAL3204 (Pal) | Peptidoglycan-associated lipoprotein | 6 (9) | OM (Class 3) | OmpA family (PF00691) | 0.507 (3) |
| BCAS0147 | Hypothetical protein | 6 (7) | U (Class 3) | Lactonase, 7-bladed beta-propeller (PF10282) | 0.463 (12) |
| BCAL3228 | Hypothetical protein | 6 (7) | U (Class 3) | none | 0.516 (3) |
| BCAL2022 | PspA/IM30 family protein | 6 (6) | OM (Class 3) | PspA/IM30 family (PF04012) | 0.503 (3) |
| BCAL1985 | Putative peptidylprolyl isomerase | 6 (6) | OM (Class 3) | PPIC-type PPIASE domain (PF13616) | 0.494 (5) |
| BCAL2083 (YaeT) | Outer membrane protein assembly factor YaeT | 6 (6) | OM (Class 3) | Surface antigen variable number repeat POTRA (PF07244) Omp85 (PF01103) | 0.488 (19) |
| BCAM0900 | Hypothetical protein | 6 (6) | U (Class 3) | Peptidase propeptide and YPEB domain (PF13670) | 0.537 (0) |
| BCAL0377 | Subfamily M24B metalopeptidase | 6 (6) | U (Class 3) | Creatinase/prolidase N-terminal domain (PF01321) (PF16189) Metallopeptidase family M24 (PF00557) C-terminal region of peptidase_M24 (PF16188) | 0.455 (12) |
| BCAM1833 (AcnB) | Bifunctional aconitate hydratase 2/2-methylisocitrate dehydratase | 6 (6) | U (Class 3) | Aconitate B, N-terminal domain (PF11791) Aconitate hydratase 2, N-terminus (PF06434) Aconitase family (PF00330) | 0.469 (16) |
| BCAL0804 | Hypothetical protein | 6 (6) | U (Class 3) | Tetratricopeptide repeat (PF14559) Tetratricopeptide repeat (PF13432) | 0.455 (20) |
| BCAM2761 (CblA) | Giant cable pilus | 5 (12) | E (Class 3) | CS1 type fimbrial major subunit (PF04449) | 0.501 (2) |
| BCAL2645 | Putative OmpA family protein | 5 (11) | OM (Class 3) | YMGG-like Gly-zipper (PF13441) OmpA family (PF00691) | 0.502 (4) |
| BCAL1288 | Family M23 peptidase | 5 (9) | OM (Class 3) | LysM domain (PF01476) Peptidase family M23 (PF01551) | 0.513 (5) |
| BCAL2738 | Hypothetical protein | 5 (7) | U (Class 3) | DUF192 (PF02643) | 0.461 (4) |
| BCAL0389 (DsbC) | Thiol:disulfide interchange protein | 5 (7) | U (Class 3) | Disulfide bond isomerase protein N-terminus (PF10411) Thioredoxin-like domain (PF13098) | 0.481 (3) |
| BCAL3035 (TrxB) | Thioredoxin reductase | 5 (6) | U (Class 3) | Pyridine nucleotide-disulphide oxidoreductase (PF07992) | 0.449 (6) |
| BCAL1881 (BamB) | Outer membrane protein assembly factor BamB | 5 (5) | OM (Class 3) | PQQ-like domain (PF13360) | 0.444 (8) |
| BCAM2549 (OpcM) | Multidrug efflux system outer membrane protein | 5 (5) | OM (Class 1) | Outer membrane efflux protein (PF02321) | 0.521 (11) |
| BCAL0304 | VacJ-like lipoprotein | 5 (5) | OM (Class 3) | MlaA lipoprotein (PF04333) | 0.469 (4) |
| BCAL0342 (BcsK) | Putative type VI secretion system protein TssC | 5 (5) | OM (Class 3) | EvpB/VC_A0108, tail sheath N-terminal domain (PF05943) | 0.472 (15) |
| BCAL0273 (CyaY) | Frataxin-like protein | 5 (5) | U (Class 3) | Frataxin-like domain (PF01491) | 0.465 (2) |
| ORF | Description | Peptides Identified 1 | Predicted B-Cell Epitopes 2 |
|---|---|---|---|
| BCAL2958 | OmpA family protein | 93ITYQADALFDFDK105 93ITYQADALFDFDKATLKPLGK113 114QKLDELASK122 116LDELASK122 123IEGMNTEVVVATGYTDR139 203RVEVEVVGTQQVQK216 204VEVEVVGTQQVQK216 | 101FDFDKATLKPLGKQ114 139RIGSDKYNDRL149 211TQQVQKTTV219 |
| BCAL2645 | OmpA family protein | 79LAPSAAQTGTQVTEQPDGSLK99 161AQSVVNALVQR171 178LSAQGMGASNPIADNATEAGR198 203RVEIYLR209 204VEIYLR209 | 88TQVTEQ93 109ATNQYAITPA118 145DSTGSAQLNQTL156 183MGASNPIADNATEAGRAQN201 |
| BCAL2022 | PspA/IM30 family protein | 13GLLNDAADSVQDPSR27 35ELDDSIGR42 43AENSLIEIEAQVATQR58 78ALQGGDEALAR88 89EALAAQSNAEAER101 150DVAASALGGIGGK162 | 20DSVQDPSRD28 156LGGIGGKNLSEDFQKLEDK174 215AALKKQLD222 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, S.A.; Seixas, A.M.M.; Mandal, M.; Rodríguez-Ortega, M.J.; Leitão, J.H. Characterization of the Burkholderia cenocepacia J2315 Surface-Exposed Immunoproteome. Vaccines 2020, 8, 509. https://doi.org/10.3390/vaccines8030509
Sousa SA, Seixas AMM, Mandal M, Rodríguez-Ortega MJ, Leitão JH. Characterization of the Burkholderia cenocepacia J2315 Surface-Exposed Immunoproteome. Vaccines. 2020; 8(3):509. https://doi.org/10.3390/vaccines8030509
Chicago/Turabian StyleSousa, Sílvia A., António M.M. Seixas, Manoj Mandal, Manuel J. Rodríguez-Ortega, and Jorge H. Leitão. 2020. "Characterization of the Burkholderia cenocepacia J2315 Surface-Exposed Immunoproteome" Vaccines 8, no. 3: 509. https://doi.org/10.3390/vaccines8030509
APA StyleSousa, S. A., Seixas, A. M. M., Mandal, M., Rodríguez-Ortega, M. J., & Leitão, J. H. (2020). Characterization of the Burkholderia cenocepacia J2315 Surface-Exposed Immunoproteome. Vaccines, 8(3), 509. https://doi.org/10.3390/vaccines8030509