Host Receptors of Influenza Viruses and Coronaviruses—Molecular Mechanisms of Recognition

Abstract
:1. Introduction
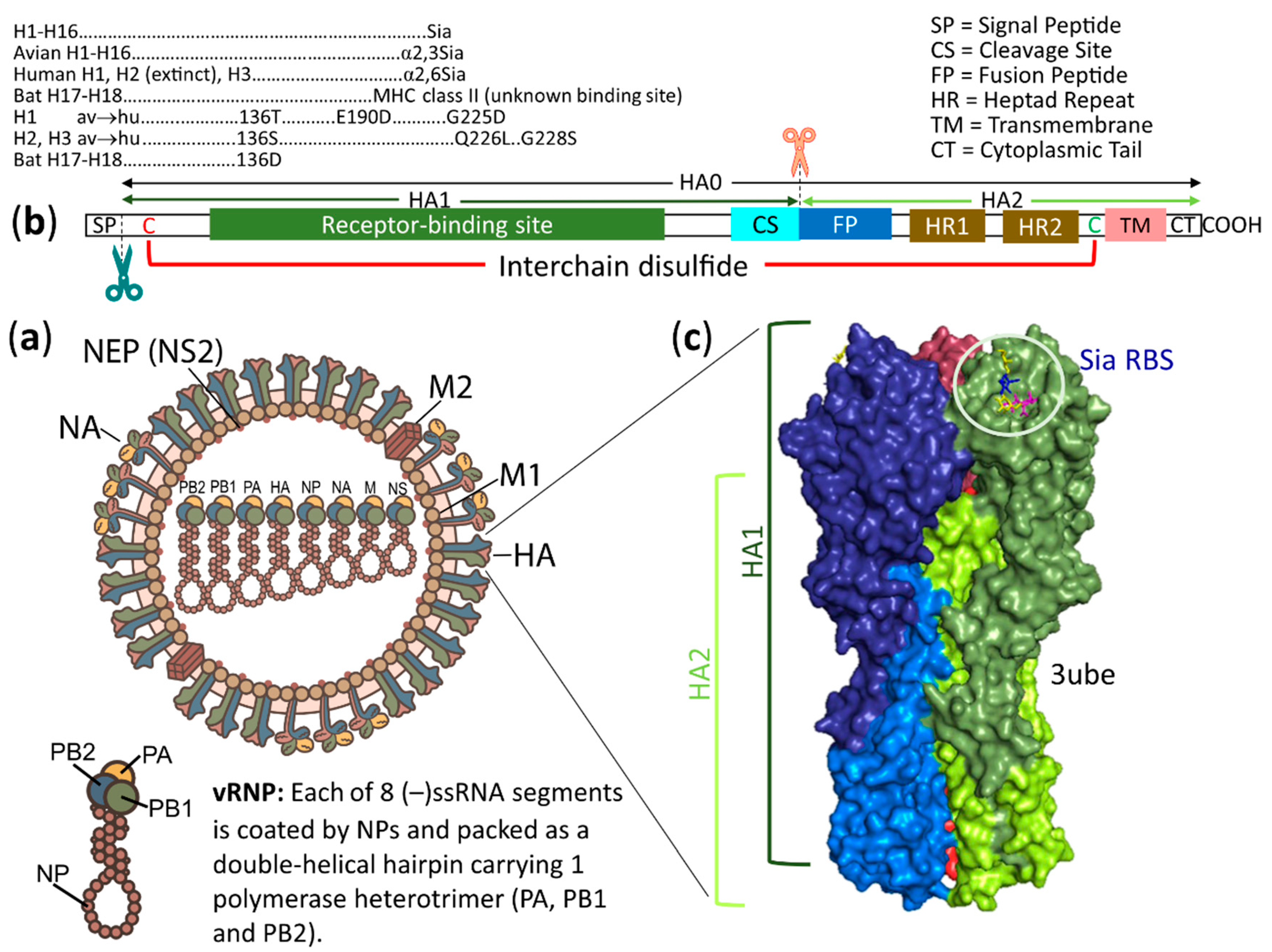
2. Overview of Structures, Host Ranges and Receptors of Influenza Viruses and Coronaviruses
3. Sialyl Glycan Receptor-Dependent Recognition of Influenza A (H1–H16) Viruses, Egyptian Fruit Bat-Isolated Influenza A Virus, Influenza B, C and D Viruses and Lineage A βCoVs (β1CoVs)
3.1. Influenza A (H1–H16) Viruses Use Siaα2,3/2,6Gal Receptors
3.2. Egyptian Fruit Bat-Isolated Influenza A Virus Uses Siaα2,3Gal Receptors
3.3. Influenza B Viruses Use Siaα2,3/2,6Gal Receptors
3.4. Influenza C Viruses Use Neu5,9Ac2
3.5. Influenza D Viruses Use Neu5,9Ac2 and Neu5Gc9Ac Receptors
3.6. β1 HCoV-OC43 and β1 HCoV-HKU1 Use Neu5,9Ac2 Receptors
4. Protein Receptor-Dependent Recognition of Influenza A (H17–H18) Viruses and αCoVs, Lineage B βCoVs (β2CoVs)
4.1. Frugivorous Bat-Identified Influenza A (H17–H18) Viruses Use MHC Class II as a Receptor
4.2. α HCoV-229E Uses hAPN Receptors
4.3. α HCoV-NL63, β2 SARS-CoV and β2 SARS-CoV-2 Use hACE2 Receptors
4.3.1. α HCoV-NL63
4.3.2. β2 SARS-CoV
4.3.3. β2 SARS-CoV-2
4.3.4. Receptor Binding Specificity of α HCoV-NL63, β2 SARS-CoV and β2 SARS-CoV-2
5. Sialyl Glycan and Protein Receptor-Dependent Recognition of Lineage C β (β3) MERS-CoV
6. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Fischer, M.; Staples, J.E.; Arboviral Diseases Branch; National Center for Emerging and Zoonotic Infectious Diseases; CDC. Notes from the field: Chikungunya virus spreads in the Americas-Caribbean and South America, 2013–2014. Morb. Mortal. Wkly. Rep. 2014, 63, 500–501. [Google Scholar] [PubMed]
- Hennessey, M.; Fischer, M.; Staples, J.E. Zika virus spreads to new areas-Region of the Americas, May 2015–January 2016. Morb. Mortal. Wkly. Rep. 2016, 65, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Kamorudeen, R.T.; Adedokun, K.A.; Olarinmoye, A.O. Ebola outbreak in West Africa, 2014–2016: Epidemic timeline, differential diagnoses, determining factors, and lessons for future response. J. Infect. Public Health 2020, 13, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Sriwilaijaroen, N.; Suzuki, Y. Molecular basis of a pandemic of avian-type influenza virus. Methods Mol. Biol. 2014, 1200, 447–480. [Google Scholar] [PubMed]
- WHO. Cumulative Number of Confirmed Human Cases for Avian Influenza A(H5N1) Reported to WHO, 2003–2020. Available online: https://www.who.int/influenza/human_animal_interface/2020_10_07_tableH5N1.pdf?ua=1 (accessed on 1 September 2020).
- WHO. Cumulative Number of Reported Probable Cases of SARS. Available online: https://www.who.int/csr/sars/country/2003_07_11/en/ (accessed on 1 September 2020).
- WHO. China Reports Additional SARS Cases-update. Available online: https://www.who.int/csr/don/2004_04_23/en/ (accessed on 1 September 2020).
- WHO. Middle East Respiratory Syndrome Coronavirus (MERS-CoV)–Saudi Arabia. Available online: https://www.who.int/csr/don/02-jul-2020-mers-saudi-arabia/en/ (accessed on 1 September 2020).
- Heymann, D.L.; Shindo, N. COVID-19: What is next for public health? Lancet 2020, 395, 542–545. [Google Scholar] [CrossRef] [Green Version]
- Sriwilaijaroen, N.; Suzuki, Y. Molecular basis of the structure and function of H1 hemagglutinin of influenza virus. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2012, 88, 226–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Timeline of WHO’s Response to COVID-19. Available online: https://www.who.int/news-room/detail/29-06-2020-covidtimeline (accessed on 1 September 2020).
- WHO. Influenza (Seasonal). Available online: https://www.who.int/news-room/fact-sheets/detail/influenza-(seasonal) (accessed on 1 September 2020).
- Calvo, C.; Garcia-Garcia, M.L.; Centeno, M.; Perez-Brena, P.; Casas, I. Influenza C virus infection in children, Spain. Emerg. Infect. Dis. 2006, 12, 1621–1622. [Google Scholar] [CrossRef]
- Flynn, O.; Gallagher, C.; Mooney, J.; Irvine, C.; Ducatez, M.; Hause, B.; McGrath, G.; Ryan, E. Influenza D virus in cattle, Ireland. Emerg. Infect. Dis. 2018, 24, 389–391. [Google Scholar] [CrossRef] [Green Version]
- White, S.K.; Ma, W.; McDaniel, C.J.; Gray, G.C.; Lednicky, J.A. Serologic evidence of exposure to influenza D virus among persons with occupational contact with cattle. J. Clin. Virol. 2016, 81, 31–33. [Google Scholar] [CrossRef]
- Horimoto, T.; Kawaoka, Y. Influenza: Lessons from past pandemics, warnings from current incidents. Nat. Rev. Microbiol. 2005, 3, 591–600. [Google Scholar] [CrossRef]
- Wertheim, J.O. The re-emergence of H1N1 influenza virus in 1977: A cautionary tale for estimating divergence times using biologically unrealistic sampling dates. PLoS ONE 2010, 5, e11184. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.; Oldstone, M.B.; Palese, P. Protective immunity and susceptibility to infectious diseases: Lessons from the 1918 influenza pandemic. Nat. Commun. 2007, 8, 1188–1193. [Google Scholar] [CrossRef] [PubMed]
- Sriwilaijaroen, N.; Nakakita, S.I.; Kondo, S.; Yagi, H.; Kato, K.; Murata, T.; Hiramatsu, H.; Kawahara, T.; Watanabe, Y.; Kanai, Y.; et al. N-glycan structures of human alveoli provide insight into influenza A virus infection and pathogenesis. FEBS J. 2018, 285, 1611–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, genetic recombination, and pathogenesis of coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.X.; Liang, J.Q.; Fung, T.S. Human Coronavirus-229E,-OC43,-NL63, and-HKU1. Ref. Modul. Life Sci. 2020. [Google Scholar] [CrossRef]
- CDC. SARS Update—19 May 2004. Available online: https://www.cdc.gov/sars/media/2004-05-19.html (accessed on 1 September 2020).
- Gambaryan, A.S.; Matrosovich, T.Y.; Boravleva, E.Y.; Lomakina, N.F.; Yamnikova, S.S.; Tuzikov, A.B.; Pazynina, G.V.; Bovin, N.V.; Fouchier, R.A.M.; Klenk, H.D.; et al. Receptor-binding properties of influenza viruses isolated from gulls. Virology 2018, 522, 37–45. [Google Scholar] [CrossRef]
- OIE. Update on Avian Influenza in Animals (types H5 and H7). Available online: https://www.oie.int/animal-health-in-the-world/update-on-avian-influenza%20/ (accessed on 1 September 2020).
- Deng, G.; Tan, D.; Shi, J.; Cui, P.; Jiang, Y.; Liu, L.; Tian, G.; Kawaoka, Y.; Li, C.; Chen, H. Complex reassortment of multiple subtypes of avian influenza viruses in domestic ducks at the Dongting Lake Region of China. J. Virol. 2013, 87, 9452–9462. [Google Scholar] [CrossRef] [Green Version]
- Laudert, E.; Sivanandan, V.; Halvorson, D.; Shaw, D.; Webster, R.G. Biological and molecular characterization of H13N2 influenza type A viruses isolated from turkeys and surface water. Avian Dis. 1993, 37, 793–799. [Google Scholar] [CrossRef]
- Zhao, G.; Gu, X.; Lu, X.; Pan, J.; Duan, Z.; Zhao, K.; Gu, M.; Liu, Q.; He, L.; Chen, J.; et al. Novel reassortant highly pathogenic H5N2 avian influenza viruses in poultry in China. PLoS ONE 2012, 7, e46183. [Google Scholar] [CrossRef]
- Gu, M.; Liu, W.; Cao, Y.; Peng, D.; Wang, X.; Wan, H.; Zhao, G.; Xu, Q.; Zhang, W.; Song, Q.; et al. Novel reassortant highly pathogenic avian influenza (H5N5) viruses in domestic ducks, China. Emerg. Infect. Dis. 2011, 17, 1060. [Google Scholar] [CrossRef]
- Wong, F.Y.; Phommachanh, P.; Kalpravidh, W.; Chanthavisouk, C.; Gilbert, J.; Bingham, J.; Davies, K.R.; Cooke, J.; Eagles, D.; Phiphakhavong, S.; et al. Reassortant highly pathogenic influenza A(H5N6) virus in Laos. Emerg. Infect. Dis. 2015, 21, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Monne, I.; Fusaro, A.; Nelson, M.I.; Bonfanti, L.; Mulatti, P.; Hughes, J.; Murcia, P.R.; Schivo, A.; Valastro, V.; Moreno, A.; et al. Emergence of a highly pathogenic avian influenza virus from a low-pathogenic progenitor. J. Virol. 2014, 88, 4375–4388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Deng, G.; Ma, S.; Zeng, X.; Yin, X.; Li, M.; Zhang, B.; Cui, P.; Chen, Y.; Yang, H.; et al. Rapid evolution of H7N9 highly pathogenic viruses that emerged in China in 2017. Cell Host Microbe 2018, 24, 558–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berhane, Y.; Hisanaga, T.; Kehler, H.; Neufeld, J.; Manning, L.; Argue, C.; Handel, K.; Hooper-McGrevy, K.; Jonas, M.; Robinson, J.; et al. Highly pathogenic avian influenza virus A (H7N3) in domestic poultry, Saskatchewan, Canada, 2007. Emerg. Infect. Dis. 2009, 15, 1492. [Google Scholar] [CrossRef] [PubMed]
- Jonges, M.; Bataille, A.; Enserink, R.; Meijer, A.; Fouchier, R.A.; Stegeman, A.; Koch, G.; Koopmans, M. Comparative analysis of avian influenza virus diversity in poultry and humans during a highly pathogenic avian influenza A (H7N7) virus outbreak. J. Virol. 2011, 85, 10598–10604. [Google Scholar] [CrossRef] [Green Version]
- Selleck, P.W.; Arzey, G.; Kirkland, P.D.; Reece, R.L.; Gould, A.R.; Daniels, P.W.; Westbury, H.A. An outbreak of highly pathogenic avian influenza in Australia in 1997 caused by an H7N4 virus. Avian Dis. 2003, 47, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Chen, L.H.; Chen, Y.P.; Liu, Y.P.; Li, W.C.; Lin, Y.L.; Lee, F. Highly pathogenic avian influenza viruses H5N2, H5N3, and H5N8 in Taiwan in 2015. Vet. Microbiol. 2016, 187, 50–57. [Google Scholar] [CrossRef]
- Heider, A.; Mochalova, L.; Harder, T.; Tuzikov, A.; Bovin, N.; Wolff, T.; Matrosovich, M.; Schweiger, B. Alterations in hemagglutinin receptor-binding specificity accompany the emergence of highly pathogenic avian influenza viruses. J. Virol. 2015, 89, 5395–5405. [Google Scholar] [CrossRef] [Green Version]
- Ping, J.; Selman, M.; Tyler, S.; Forbes, N.; Keleta, L.; Brown, E.G. Low-pathogenic avian influenza virus A/turkey/Ontario/6213/1966 (H5N1) is the progenitor of highly pathogenic A/turkey/Ontario/7732/1966 (H5N9). J. Gen. Virol. 2012, 93, 1649–1657. [Google Scholar] [CrossRef]
- CDC. Highly Pathogenic Avian Influenza A(H5N1) in Birds and other Animals. Available online: https://www.cdc.gov/flu/avianflu/h5n1-animals.htm (accessed on 1 September 2020).
- Sriwilaijaroen, N.; Kondo, S.; Yagi, H.; Takemae, N.; Saito, T.; Hiramatsu, H.; Kato, K.; Suzuki, Y. N-glycans from porcine trachea and lung: Predominant NeuAcα2-6Gal could be a selective pressure for influenza variants in favor of human-type receptor. PLoS ONE 2011, 6, e16302. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Horiike, G.; Yamazaki, Y.; Kawabe, K.; Masuda, H.; Miyamoto, D.; Matsuda, M.; Nishimura, S.I.; Yamagata, T.; Ito, T.; et al. Swine influenza virus strains recognize sialylsugar chains containing the molecular species of sialic acid predominantly present in the swine tracheal epithelium. FEBS Lett. 1997, 404, 192–196. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Couceiro, J.N.; Kelm, S.; Baum, L.G.; Krauss, S.; Castrucci, M.R.; Donatelli, I.; Kida, H.; Paulson, J.C.; Webster, R.G.; et al. Molecular basis for the generation in pigs of influenza A viruses with pandemic potential. J. Virol. 1998, 72, 7367–7373. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Ito, T.; Suzuki, T.; Holland, R.E., Jr.; Chambers, T.M.; Kiso, M.; Ishida, H.; Kawaoka, Y. Sialic acid species as a determinant of the host range of influenza A viruses. J. Virol. 2000, 74, 11825–11831. [Google Scholar] [CrossRef] [Green Version]
- Feng, K.H.; Gonzalez, G.; Deng, L.; Yu, H.; Tse, V.L.; Huang, L.; Huang, K.; Wasik, B.R.; Zhou, B.; Wentworth, D.E.; et al. Equine and canine influenza H3N8 viruses show minimal biological differences despite phylogenetic divergence. J. Virol. 2015, 89, 6860–6873. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.; Sun, S.; Du, L.; Ma, J.; Fan, L.; Pu, J.; Sun, Y.; Zhao, J.; Sun, H.; Liu, J. Natural and experimental infection of dogs with pandemic H1N1/2009 influenza virus. J. Gen. Virol. 2012, 93, 119–123. [Google Scholar] [CrossRef]
- Pulit-Penaloza, J.A.; Simpson, N.; Yang, H.; Creager, H.M.; Jones, J.; Carney, P.; Belser, J.A.; Yang, G.; Chang, J.; Zeng, H.; et al. Assessment of molecular, antigenic, and pathological features of canine influenza A(H3N2) viruses that emerged in the United States. J. Infect. 2017, 216, S499–S507. [Google Scholar] [CrossRef] [Green Version]
- Song, Q.Q.; Zhang, F.X.; Liu, J.J.; Ling, Z.S.; Zhu, Y.L.; Jiang, S.J.; Xie, Z.J. Dog to dog transmission of a novel influenza virus (H5N2) isolated from a canine. Vet. Microbiol. 2013, 161, 331–333. [Google Scholar] [CrossRef]
- Song, D.S.; An, D.J.; Moon, H.J.; Yeom, M.J.; Jeong, H.Y.; Jeong, W.S.; Park, S.J.; Kim, H.K.; Han, S.Y.; Oh, J.S.; et al. Interspecies transmission of the canine influenza H3N2 virus to domestic cats in South Korea, 2010. J. Gen. Virol. 2011, 92, 2350–2355. [Google Scholar] [CrossRef]
- Hatta, M.; Zhong, G.; Gao, Y.; Nakajima, N.; Fan, S.; Chiba, S.; Deering, K.; Ito, M.; Imai, M.; Kiso, M.; et al. Characterization of a feline Influenza A(H7N2) virus. Emerg. Infect. Dis. 2018, 24, 75. [Google Scholar] [CrossRef]
- Patterson, A.R.; Cooper, V.L.; Yoon, K.J.; Janke, B.H.; Gauger, P.C. Naturally occurring influenza infection in a ferret (Mustela putorius furo) colony. J. Vet. Diagn. Investig. 2009, 21, 527–530. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.T.; Wang, C.H.; Wu, W.L.; Chi, C.H.; Wang, L.C. Natural A (H1N1) pdm09 influenza virus infection case in a pet ferret in Taiwan. Jpn. J. Vet. Res. 2014, 62, 181–185. [Google Scholar]
- Hinshaw, V.S.; Bean, W.J.; Geraci, J.; Fiorelli, P.; Early, G.; Webster, R.G. Characterization of two influenza A viruses from a pilot whale. J. Virol. 1986, 58, 655–656. [Google Scholar] [CrossRef] [Green Version]
- Hussein, I.T.M.; Krammer, F.; Ma, E.; Estrin, M.; Viswanathan, K.; Stebbins, N.W.; Quinlan, D.S.; Sasisekharan, R.; Runstadler, J. New England harbor seal H3N8 influenza virus retains avian-like receptor specificity. Sci. Rep. 2016, 6, 21428. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Kawaoka, Y.; Nomura, A.; Otsuki, K. Receptor specificity of influenza A viruses from sea mammals correlates with lung sialyloligosaccharides in these animals. J. Vet. Med. Sci. 1999, 61, 955–958. [Google Scholar] [CrossRef] [Green Version]
- Naguib, M.M.; Kinne, J.; Chen, H.; Chan, K.-H.; Joseph, S.; Wong, P.-C.; Woo, P.C.Y.; Wernery, R.; Beer, M.; Wernery, U.; et al. Outbreaks of highly pathogenic avian influenza H5N1 clade 2.3.2.1c in hunting falcons and kept wild birds in Dubai implicate intercontinental virus spread. J. Gen. Virol. 2015, 96, 3212–3222. [Google Scholar] [CrossRef]
- Yong-Feng, Z.; Fei-Fei, D.; Jia-Yu, Y.; Feng-Xia, Z.; Chang-Qing, J.; Jian-Li, W.; Shou-Yu, G.; Kai, C.; Chuan-Yi, L.; Xue-Hua, W.; et al. Intraspecies and interspecies transmission of mink H9N2 influenza virus. Sci. Rep. 2017, 7, 7429. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, E.; Fujii, K.; Ogawa, H.; Imai, K. First detection of influenza A virus genes from wild raccoons in Japan. Virus Genes 2018, 54, 591–595. [Google Scholar] [CrossRef]
- WHO. Influenza (Avian and other Zoonotic). Available online: https://www.who.int/news-room/fact-sheets/detail/influenza-(avian-and-other-zoonotic) (accessed on 1 September 2020).
- Zhang, T.; Bi, Y.; Tian, H.; Li, X.; Liu, D.; Wu, Y.; Jin, T.; Wang, Y.; Chen, Q.; Chen, Z.; et al. Human infection with influenza virus A(H10N8) from live poultry markets, China, 2014. Emerg. Infect. Dis. 2014, 20, 2076–2079. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Zhang, L.; Kan, X.; Jiang, L.; Yang, J.; Guo, Z.; Ren, Q. Origin and molecular characteristics of a novel 2013 avian influenza A(H6N1) virus causing human infection in Taiwan. Clin. Infect. Dis. 2013, 57, 1367–1368. [Google Scholar] [CrossRef] [Green Version]
- Gulati, S.; Smith, D.F.; Cummings, R.D.; Couch, R.B.; Griesemer, S.B.; St George, K.; Webster, R.G.; Air, G.M. Human H3N2 influenza viruses isolated from 1968 to 2012 show varying preference for receptor substructures with no apparent consequences for disease or spread. PLoS ONE 2013, 8, e66325. [Google Scholar] [CrossRef] [Green Version]
- Childs, R.A.; Palma, A.S.; Wharton, S.; Matrosovich, T.; Liu, Y.; Chai, W.; Campanero-Rhodes, M.A.; Zhang, Y.; Eickmann, M.; Kiso, M.; et al. Receptor-binding specificity of pandemic influenza A (H1N1) 2009 virus determined by carbohydrate microarray. Nat. Biotechnol. 2009, 27, 797–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karakus, U.; Thamamongood, T.; Ciminski, K.; Ran, W.; Gunther, S.C.; Pohl, M.O.; Eletto, D.; Jeney, C.; Hoffmann, D.; Reiche, S.; et al. MHC class II proteins mediate cross-species entry of bat influenza viruses. Nature 2019, 567, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Giotis, E.S.; Carnell, G.; Young, E.F.; Ghanny, S.; Soteropoulos, P.; Wang, L.F.; Barclay, W.S.; Skinner, M.A.; Temperton, N. Entry of the bat influenza H17N10 virus into mammalian cells is enabled by the MHC class II HLA-DR receptor. Nat. Microbiol. 2019, 4, 2035–2038. [Google Scholar] [CrossRef] [PubMed]
- Kandeil, A.; Gomaa, M.R.; Shehata, M.M.; El Taweel, A.N.; Mahmoud, S.H.; Bagato, O.; Moatasim, Y.; Kutkat, O.; Kayed, A.S.; Dawson, P.; et al. Isolation and characterization of a distinct influenza A virus from Egyptian bats. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.P.; Tsai, H.J. Influenza B viruses in pigs, Taiwan. Influenza Other Respir. Viruses 2019, 13, 91–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.F.; Chang, C.F.; Chi, C.Y.; Wang, H.C.; Wang, J.R.; Su, I.J. Characterization of glycan binding specificities of influenza B viruses with correlation with hemagglutinin genotypes and clinical features. J. Med. Virol. 2012, 84, 679–685. [Google Scholar] [CrossRef]
- Lugovtsev, V.Y.; Smith, D.F.; Weir, J.P. Changes of the receptor-binding properties of influenza B virus B/Victoria/504/2000 during adaptation in chicken eggs. Virology 2009, 394, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Ni, F.; Nnadi Mbawuike, I.; Kondrashkina, E.; Wang, Q. The roles of hemagglutinin Phe-95 in receptor binding and pathogenicity of influenza B virus. Virology 2014, 450–451, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.J.; Jin, F.G.; Wang, P.; Wang, M.; Zhu, J.M. Isolation of influenza C virus from pigs and experimental infection of pigs with influenza C virus. J. Gen. Virol. 1983, 64 Pt 1, 177–182. [Google Scholar] [PubMed]
- Zhang, H.; Porter, E.; Lohman, M.; Lu, N.; Peddireddi, L.; Hanzlicek, G.; Marthaler, D.; Liu, X.; Bai, J. Influenza C virus in cattle with respiratory disease, United States, 2016–2018. Emerg. Infect. Dis. 2018, 24, 1926. [Google Scholar] [CrossRef]
- Matsuzaki, Y.; Sugawara, K.; Furuse, Y.; Shimotai, Y.; Hongo, S.; Oshitani, H.; Mizuta, K.; Nishimura, H. Genetic lineage and reassortment of influenza C viruses circulating between 1947 and 2014. J. Virol. 2016, 90, 8251–8265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Sreenivasan, C.; Yu, H.; Sheng, Z.; Newkirk, S.J.; An, W.; Smith, D.F.; Chen, X.; Wang, D.; Li, F. Influenza D virus diverges from its related influenza C virus in the recognition of 9-O-acetylated N-acetyl- or N-glycolyl-neuraminic acid-containing glycan receptors. Virology 2020, 545, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Sato, R.; Ishida, H.; Katayama, M.; Takenaka-Uema, A.; Horimoto, T. Influenza D virus of new phylogenetic lineage, Japan. Emerg. Infect. Dis. 2020, 26, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, P.C.; Lau, S.K.; Wernery, U.; Wong, E.Y.; Tsang, A.K.; Johnson, B.; Yip, C.C.; Lau, C.C.; Sivakumar, S.; Cai, J.P.; et al. Novel betacoronavirus in dromedaries of the Middle East, 2013. Emerg. Infect. Dis. 2014, 20, 560–572. [Google Scholar] [CrossRef] [Green Version]
- Jaimes, J.A.; Whittaker, G.R. Feline coronavirus: Insights into viral pathogenesis based on the spike protein structure and function. Virology 2018, 517, 108–121. [Google Scholar] [CrossRef]
- Schultze, B.; Krempl, C.; Ballesteros, M.L.; Shaw, L.; Schauer, R.; Enjuanes, L.; Herrler, G. Transmissible gastroenteritis coronavirus, but not the related porcine respiratory coronavirus, has a sialic acid (N-glycolylneuraminic acid) binding activity. J. Virol. 1996, 70, 5634–5637. [Google Scholar] [CrossRef] [Green Version]
- Stoian, A.; Rowland, R.R.R.; Petrovan, V.; Sheahan, M.; Samuel, M.S.; Whitworth, K.M.; Wells, K.D.; Zhang, J.; Beaton, B.; Cigan, M.; et al. The use of cells from ANPEP knockout pigs to evaluate the role of aminopeptidase N (APN) as a receptor for porcine deltacoronavirus (PDCoV). Virology 2020, 541, 136–140. [Google Scholar] [CrossRef]
- Li, W.; van Kuppeveld, F.J.M.; He, Q.; Rottier, P.J.M.; Bosch, B.J. Cellular entry of the porcine epidemic diarrhea virus. Virus Res. 2016, 226, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Shirato, K.; Maejima, M.; Islam, M.T.; Miyazaki, A.; Kawase, M.; Matsuyama, S.; Taguchi, F. Porcine aminopeptidase N is not a cellular receptor of porcine epidemic diarrhea virus, but promotes its infectivity via aminopeptidase activity. J. Gen. Virol. 2016, 97, 2528–2539. [Google Scholar] [CrossRef]
- Hansen, G.H.; Delmas, B.; Besnardeau, L.; Vogel, L.K.; Laude, H.; Sjostrom, H.; Noren, O. The coronavirus transmissible gastroenteritis virus causes infection after receptor-mediated endocytosis and acid-dependent fusion with an intracellular compartment. J. Virol. 1998, 72, 527–534. [Google Scholar] [CrossRef] [Green Version]
- Hohdatsu, T.; Izumiya, Y.; Yokoyama, Y.; Kida, K.; Koyama, H. Differences in virus receptor for type I and type II feline infectious peritonitis virus. Arch. Virol. 1998, 143, 839–850. [Google Scholar] [CrossRef]
- Regan, A.D.; Ousterout, D.G.; Whittaker, G.R. Feline lectin activity is critical for the cellular entry of feline infectious peritonitis virus. J. Virol. 2010, 84, 7917–7921. [Google Scholar] [CrossRef] [Green Version]
- Wong, A.H.M.; Tomlinson, A.C.A.; Zhou, D.; Satkunarajah, M.; Chen, K.; Sharon, C.; Desforges, M.; Talbot, P.J.; Rini, J.M. Receptor-binding loops in alphacoronavirus adaptation and evolution. Nat. Commun. 2017, 8, 1735. [Google Scholar] [CrossRef]
- Hofmann, H.; Pyrc, K.; van der Hoek, L.; Geier, M.; Berkhout, B.; Pohlmann, S. Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry. Proc. Natl. Acad. Sci. USA 2005, 102, 7988–7993. [Google Scholar] [CrossRef] [Green Version]
- Lau, S.K.; Woo, P.C.; Li, K.S.; Tsang, A.K.; Fan, R.Y.; Luk, H.K.; Cai, J.P.; Chan, K.H.; Zheng, B.J.; Wang, M.; et al. Discovery of a novel coronavirus, China Rattus coronavirus HKU24, from Norway rats supports the murine origin of Betacoronavirus 1 and has implications for the ancestor of Betacoronavirus lineage A. J. Virol. 2015, 89, 3076–3092. [Google Scholar] [CrossRef] [Green Version]
- Sriwilaijaroen, N.; Suzuki, Y. Sialoglycovirology of lectins: Sialyl glycan binding of enveloped and non-enveloped Viruses. Methods Mol. Biol. 2020, 2132, 483–545. [Google Scholar] [PubMed] [Green Version]
- Bakkers, M.J.; Zeng, Q.; Feitsma, L.J.; Hulswit, R.J.; Li, Z.; Westerbeke, A.; van Kuppeveld, F.J.; Boons, G.J.; Langereis, M.A.; Huizinga, E.G.; et al. Coronavirus receptor switch explained from the stereochemistry of protein-carbohydrate interactions and a single mutation. Proc. Natl. Acad. Sci. USA 2016, 113, E3111–E3119. [Google Scholar] [CrossRef] [Green Version]
- Peng, G.; Sun, D.; Rajashankar, K.R.; Qian, Z.; Holmes, K.V.; Li, F. Crystal structure of mouse coronavirus receptor-binding domain complexed with its murine receptor. Proc. Natl. Acad. Sci. USA 2011, 108, 10696–10701. [Google Scholar] [CrossRef] [Green Version]
- Langereis, M.A.; Zeng, Q.; Heesters, B.A.; Huizinga, E.G.; de Groot, R.J. The murine coronavirus hemagglutinin-esterase receptor-binding site: A major shift in ligand specificity through modest changes in architecture. PLoS Pathog. 2012, 8, e1002492. [Google Scholar] [CrossRef]
- Langereis, M.A.; Bakkers, M.J.; Deng, L.; Padler-Karavani, V.; Vervoort, S.J.; Hulswit, R.J.; van Vliet, A.L.; Gerwig, G.J.; de Poot, S.A.; Boot, W.; et al. Complexity and diversity of the mammalian sialome revealed by nidovirus virolectins. Cell Rep. 2015, 11, 1966–1978. [Google Scholar] [CrossRef] [Green Version]
- Dong, B.; Gao, W.; Lu, H.; Zhao, K.; Ding, N.; Liu, W.; Zhao, J.; Lan, Y.; Tang, B.; Jin, Z.; et al. A small region of porcine hemagglutinating encephalomyelitis virus spike protein interacts with the neural cell adhesion molecule. Intervirology 2015, 58, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Hulswit, R.J.G.; Lang, Y.; Bakkers, M.J.G.; Li, W.; Li, Z.; Schouten, A.; Ophorst, B.; van Kuppeveld, F.J.M.; Boons, G.J.; Bosch, B.J.; et al. Human coronaviruses OC43 and HKU1 bind to 9-O-acetylated sialic acids via a conserved receptor-binding site in spike protein domain A. Proc. Natl. Acad. Sci. USA 2019, 116, 2681–2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krempl, C.; Schultze, B.; Herrler, G. Analysis of cellular receptors for human coronavirus OC43. Adv. Exp. Med. Biol. 1995, 380, 371–374. [Google Scholar] [PubMed] [Green Version]
- Bakkers, M.J.; Lang, Y.; Feitsma, L.J.; Hulswit, R.J.; de Poot, S.A.; van Vliet, A.L.; Margine, I.; de Groot-Mijnes, J.D.; van Kuppeveld, F.J.; Langereis, M.A.; et al. Betacoronavirus adaptation to humans involved progressive loss of hemagglutinin-esterase lectin activity. Cell Host Microbe 2017, 21, 356–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, S.K.; Woo, P.C.; Li, K.S.; Huang, Y.; Tsoi, H.W.; Wong, B.H.; Wong, S.S.; Leung, S.Y.; Chan, K.H.; Yuen, K.Y. Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats. Proc. Natl. Acad. Sci. USA 2005, 102, 14040–14045. [Google Scholar] [CrossRef] [Green Version]
- Bolles, M.; Donaldson, E.; Baric, R. SARS-CoV and emergent coronaviruses: Viral determinants of interspecies transmission. Curr. Opin. Virol. 2011, 1, 624–634. [Google Scholar] [CrossRef]
- Jaimes, J.A.; André, N.M.; Chappie, J.S.; Millet, J.K.; Whittaker, G.R. Phylogenetic analysis and structural modeling of SARS-CoV-2 spike protein reveals an evolutionary distinct and proteolytically sensitive activation loop. J. Mol. Biol. 2020, 432, 3309–3325. [Google Scholar] [CrossRef]
- Li, F. Structure, function, and evolution of coronavirus spike proteins. Annu. Rev. Virol. 2016, 3, 237–261. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Hu, Z. A review of studies on animal reservoirs of the SARS coronavirus. Virus Res. 2008, 133, 74–87. [Google Scholar] [CrossRef]
- Kan, B.; Wang, M.; Jing, H.; Xu, H.; Jiang, X.; Yan, M.; Liang, W.; Zheng, H.; Wan, K.; Liu, Q.; et al. Molecular evolution analysis and geographic investigation of severe acute respiratory syndrome coronavirus-like virus in palm civets at an animal market and on farms. J. Virol. 2005, 79, 11892–11900. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, R.; Dhama, K.; Sharun, K.; Iqbal Yatoo, M.; Malik, Y.S.; Singh, R.; Michalak, I.; Sah, R.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J. COVID-19: Animals, veterinary and zoonotic links. Vet. Q. 2020, 40, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Halfmann, P.J.; Hatta, M.; Chiba, S.; Maemura, T.; Fan, S.; Takeda, M.; Kinoshita, N.; Hattori, S.I.; Sakai-Tagawa, Y.; Iwatsuki-Horimoto, K.; et al. Transmission of SARS-CoV-2 in domestic cats. N. Engl. J. Med. 2020, 383, 592–594. [Google Scholar] [CrossRef] [PubMed]
- Xiao, K.; Zhai, J.; Feng, Y.; Zhou, N.; Zhang, X.; Zou, J.J.; Li, N.; Guo, Y.; Li, X.; Shen, X.; et al. Isolation of SARS-CoV-2-related coronavirus from Malayan pangolins. Nature 2020, 583, 286–289. [Google Scholar] [CrossRef] [PubMed]
- Graham, R.L.; Donaldson, E.F.; Baric, R.S. A decade after SARS: Strategies for controlling emerging coronaviruses. Nat. Rev. Microbiol. 2013, 11, 836–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glowacka, I.; Bertram, S.; Pöhlmann, S. Cellular entry of the SARS coronavirus: Implications for transmission, pathogenicity and antiviral strategies. In Molecular Biology of the SARS-Coronavirus; Lal, S.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 3–22. [Google Scholar] [CrossRef]
- Chan, J.F.; Lau, S.K.; To, K.K.; Cheng, V.C.; Woo, P.C.; Yuen, K.Y. Middle East respiratory syndrome coronavirus: Another zoonotic betacoronavirus causing SARS-like disease. Clin. Microbiol. Rev. 2015, 28, 465–522. [Google Scholar] [CrossRef] [Green Version]
- Lau, S.K.P.; Zhang, L.; Luk, H.K.H.; Xiong, L.; Peng, X.; Li, K.S.M.; He, X.; Zhao, P.S.; Fan, R.Y.Y.; Wong, A.C.P.; et al. Receptor usage of a novel bat lineage C betacoronavirus reveals evolution of Middle East respiratory syndrome-related coronavirus spike proteins for human dipeptidyl peptidase 4 binding. J. Infect. Dis. 2018, 218, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Widagdo, W.; Okba, N.M.A.; Li, W.; de Jong, A.; de Swart, R.L.; Begeman, L.; van den Brand, J.M.A.; Bosch, B.-J.; Haagmans, B.L. Species-specific colocalization of Middle East respiratory syndrome coronavirus attachment and entry receptors. J. Virol. 2019, 93, e00107–e00119. [Google Scholar] [CrossRef] [Green Version]
- Adney, D.; Bielefeldt-Ohmann, H.; Hartwig, A.; Bowen, R. Infection, replication, and transmission of Middle East respiratory syndrome coronavirus in Alpacas. Emerg. Infect. Dis. 2016, 22, 1031. [Google Scholar] [CrossRef] [Green Version]
- Khalafalla, A.; Lu, X.; Al-Mubarak, A.I.A.; Dalab, A.H.; Al-Busadah, K.A.S.; Erdman, D. MERS-CoV in upper respiratory tract and lungs of dromedary camels, Saudi Arabia, 2013–2014. Emerg. Infect. Dis. 2015, 21, 1153. [Google Scholar] [CrossRef] [Green Version]
- Alnaeem, A.; Kasem, S.; Qasim, I.; Al-Doweriej, A.; Refaat, M.; Al-Shabebi, A.; Hemida, M.G. The dipeptidyl peptidase-4 expression in some MERS-CoV naturally infected dromedary camels in Saudi Arabia 2018–2019. VirusDis. 2020, 31, 200–203. [Google Scholar] [CrossRef]
- Li, W.; Hulswit, R.J.G.; Widjaja, I.; Raj, V.S.; McBride, R.; Peng, W.; Widagdo, W.; Tortorici, M.A.; van Dieren, B.; Lang, Y.; et al. Identification of sialic acid-binding function for the Middle East respiratory syndrome coronavirus spike glycoprotein. Proc. Natl. Acad. Sci. USA 2017, 114, E8508–E8517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qing, E.; Hantak, M.; Perlman, S.; Gallagher, T. Distinct roles for sialoside and protein receptors in coronavirus infection. MBio 2020, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambepitiya Wickramasinghe, I.N.; de Vries, R.P.; Weerts, E.A.; van Beurden, S.J.; Peng, W.; McBride, R.; Ducatez, M.; Guy, J.; Brown, P.; Eterradossi, N.; et al. Novel receptor specificity of avian gammacoronaviruses that cause enteritis. J. Virol. 2015, 89, 8783–8792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madu, I.G.; Chu, V.C.; Lee, H.; Regan, A.D.; Bauman, B.E.; Whittaker, G.R. Heparan sulfate is a selective attachment factor for the avian coronavirus infectious bronchitis virus beaudette. Avian Dis. 2007, 51, 45–51, 47. [Google Scholar] [CrossRef] [Green Version]
- Lau, S.K.P.; Wong, E.Y.M.; Tsang, C.C.; Ahmed, S.S.; Au-Yeung, R.K.H.; Yuen, K.Y.; Wernery, U.; Woo, P.C.Y. Discovery and sequence analysis of four deltacoronaviruses from birds in the Middle East reveal interspecies jumping with recombination as a potential mechanism for avian-to-avian and avian-to-mammalian transmission. J. Virol. 2018, 92, e00265-18. [Google Scholar] [CrossRef] [Green Version]
- International Committee on Taxonomy of Viruses (ICTV). Available online: https://talk.ictvonline.org/ (accessed on 27 September 2020).
- Coronaviridae. Fenner’s Veterinary Virology (Fifth Edition); MacLachlan, N.J., Dubovi, E.J., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 435–461. [Google Scholar]
- Tong, S.; Li, Y.; Rivailler, P.; Conrardy, C.; Castillo, D.A.; Chen, L.M.; Recuenco, S.; Ellison, J.A.; Davis, C.T.; York, I.A.; et al. A distinct lineage of influenza A virus from bats. Proc. Natl. Acad. Sci. USA 2012, 109, 4269–4274. [Google Scholar] [CrossRef] [Green Version]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco, S.; Gomez, J.; et al. New world bats harbor diverse influenza A viruses. PLoS Pathog. 2013, 9, e1003657. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Suzuki, Y.; Suzuki, T.; Takada, A.; Horimoto, T.; Wells, K.; Kida, H.; Otsuki, K.; Kiso, M.; Ishida, H.; et al. Recognition of N-glycolylneuraminic acid linked to galactose by the α2,3 linkage is associated with intestinal replication of influenza A virus in ducks. J. Virol. 2000, 74, 9300–9305. [Google Scholar] [CrossRef] [Green Version]
- Hiono, T.; Okamatsu, M.; Nishihara, S.; Takase-Yoden, S.; Sakoda, Y.; Kida, H. A chicken influenza virus recognizes fucosylated a2,3 sialoglycan receptors on the epithelial cells lining upper respiratory tracts of chickens. Virology 2014, 456–457, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Venkatesh, D.; Poen, M.J.; Bestebroer, T.M.; Scheuer, R.D.; Vuong, O.; Chkhaidze, M.; Machablishvili, A.; Mamuchadze, J.; Ninua, L.; Fedorova, N.B.; et al. Avian influenza viruses in wild birds: Virus evolution in a multihost ecosystem. J. Virol. 2018, 92, e00433-18. [Google Scholar] [CrossRef] [Green Version]
- Smits, S.L.; Gerwig, G.J.; van Vliet, A.L.; Lissenberg, A.; Briza, P.; Kamerling, J.P.; Vlasak, R.; de Groot, R.J. Nidovirus sialate-O-acetylesterases: Evolution and substrate specificity of coronaviral and toroviral receptor-destroying enzymes. J. Biol. Chem. 2005, 280, 6933–6941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfefferle, S.; Oppong, S.; Drexler, J.F.; Gloza-Rausch, F.; Ipsen, A.; Seebens, A.; Muller, M.A.; Annan, A.; Vallo, P.; Adu-Sarkodie, Y.; et al. Distant relatives of severe acute respiratory syndrome coronavirus and close relatives of human coronavirus 229E in bats, Ghana. Emerg. Infect. Dis. 2009, 15, 1377–1384. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Baldwin, H.J.; Tateno, A.F.; Zerbinati, R.M.; Annan, A.; Owusu, M.; Nkrumah, E.E.; Maganga, G.D.; Oppong, S.; Adu-Sarkodie, Y.; et al. Evidence for an ancestral association of human coronavirus 229E with bats. J. Virol. 2015, 89, 11858–11870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.; Shi, M.; Chommanard, C.; Queen, K.; Zhang, J.; Markotter, W.; Kuzmin, I.V.; Holmes, E.C.; Tong, S. Surveillance of bat coronaviruses in Kenya identifies relatives of human coronaviruses NL63 and 229E and their recombination history. J. Virol. 2017, 91, e01953-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Zeng, L.P.; Yang, X.L.; Ge, X.Y.; Zhang, W.; Li, B.; Xie, J.Z.; Shen, X.R.; Zhang, Y.Z.; Wang, N.; et al. Discovery of a rich gene pool of bat SARS-related coronaviruses provides new insights into the origin of SARS coronavirus. PLoS Pathog. 2017, 13, e1006698. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.P.; Feng, Y.; Chen, H.; Luk, H.K.H.; Yang, W.H.; Li, K.S.M.; Zhang, Y.Z.; Huang, Y.; Song, Z.Z.; Chow, W.-N.; et al. Severe acute respiratory syndrome (SARS) coronavirus ORF8 protein Is acquired from SARS-related coronavirus from greater horseshoe bats through recombination. J. Virol. 2015, 89, 10532–10547. [Google Scholar] [CrossRef] [Green Version]
- Hon, C.C.; Lam, T.Y.; Shi, Z.L.; Drummond, A.J.; Yip, C.W.; Zeng, F.; Lam, P.Y.; Leung, F.C. Evidence of the recombinant origin of a bat severe acute respiratory syndrome (SARS)-like coronavirus and its implications on the direct ancestor of SARS coronavirus. J. Virol. 2008, 82, 1819–1826. [Google Scholar] [CrossRef] [Green Version]
- Lau, S.K.P.; Li, K.S.M.; Tsang, A.K.L.; Lam, C.S.F.; Ahmed, S.; Chen, H.; Chan, K.-H.; Woo, P.C.Y.; Yuen, K.-Y. Genetic characterization of betacoronavirus lineage C viruses in bats reveals marked sequence divergence in the spike protein of Pipistrellus bat coronavirus HKU5 in Japanese pipistrelle: Implications for the origin of the novel Middle East respiratory syndrome coronavirus. J. Virol. 2013, 87, 8638–8650. [Google Scholar] [PubMed] [Green Version]
- Zhou, P.; Yang, X.L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Chen, X.; Hu, T.; Li, J.; Song, H.; Liu, Y.; Wang, P.; Liu, D.; Yang, J.; Holmes, E.C.; et al. A novel bat coronavirus closely related to SARS-CoV-2 contains natural insertions at the S1/S2 cleavage site of the spike protein. Curr. Biol. 2020, 30, 2196–2203. [Google Scholar] [CrossRef]
- Braakman, I.; Hoover-Litty, H.; Wagner, K.R.; Helenius, A. Folding of influenza hemagglutinin in the endoplasmic reticulum. J. Cell. Biol. 1991, 114, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Steinhauer, D.A. Role of hemagglutinin cleavage for the pathogenicity of influenza virus. Virology 1999, 258, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Kido, H.; Okumura, Y.; Takahashi, E.; Pan, H.Y.; Wang, S.; Chida, J.; Le, T.Q.; Yano, M. Host envelope glycoprotein processing proteases are indispensable for entry into human cells by seasonal and highly pathogenic avian influenza viruses. J. Mol. Genet. Med. 2008, 3, 167–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, R.T.; Kushnir, A.S.; White, J.M. The transmembrane domain of influenza hemagglutinin exhibits a stringent length requirement to support the hemifusion to fusion transition. J. Cell Biol. 2000, 151, 425–437. [Google Scholar] [CrossRef] [Green Version]
- Ohuchi, M.; Fischer, C.; Ohuchi, R.; Herwig, A.; Klenk, H.D. Elongation of the cytoplasmic tail interferes with the fusion activity of influenza virus hemagglutinin. J. Virol. 1998, 72, 3554–3559. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; McBride, R.; Nycholat, C.M.; Paulson, J.C.; Wilson, I.A. Structural characterization of the hemagglutinin receptor specificity from the 2009 H1N1 influenza pandemic. J. Virol. 2012, 86, 982–990. [Google Scholar] [CrossRef] [Green Version]
- Neuman, B.W.; Kiss, G.; Kunding, A.H.; Bhella, D.; Baksh, M.F.; Connelly, S.; Droese, B.; Klaus, J.P.; Makino, S.; Sawicki, S.G.; et al. A structural analysis of M protein in coronavirus assembly and morphology. J. Struct. Biol. 2011, 174, 11–22. [Google Scholar] [CrossRef]
- Schoeman, D.; Fielding, B.C. Coronavirus envelope protein: Current knowledge. Virol. J. 2019, 16, 69. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, T.M.; Buchmeier, M.J. Coronavirus spike proteins in viral entry and pathogenesis. Virology 2001, 279, 371–374. [Google Scholar] [CrossRef] [Green Version]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Q.; Langereis, M.A.; van Vliet, A.L.; Huizinga, E.G.; de Groot, R.J. Structure of coronavirus hemagglutinin-esterase offers insight into corona and influenza virus evolution. Proc. Natl. Acad. Sci. USA 2008, 105, 9065–9069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rota, P.A.; Oberste, M.S.; Monroe, S.S.; Nix, W.A.; Campagnoli, R.; Icenogle, J.P.; Peñaranda, S.; Bankamp, B.; Maher, K.; Chen, M.-H.; et al. Characterization of a novel coronavirus associated with severe acute respiratory syndrome. Science 2003, 300, 1394–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coutard, B.; Valle, C.; de Lamballerie, X.; Canard, B.; Seidah, N.G.; Decroly, E. The spike glycoprotein of the new coronavirus 2019-nCoV contains a furin-like cleavage site absent in CoV of the same clade. Antivir. Res. 2020, 176, 104742. [Google Scholar] [CrossRef]
- Bosch, B.J.; van der Zee, R.; de Haan, C.A.; Rottier, P.J. The coronavirus spike protein is a class I virus fusion protein: Structural and functional characterization of the fusion core complex. J. Virol. 2003, 77, 8801–8811. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.J.; Walls, A.C.; Wang, Z.; Sauer, M.M.; Li, W.; Tortorici, M.A.; Bosch, B.J.; DiMaio, F.; Veesler, D. Structures of MERS-CoV spike glycoprotein in complex with sialoside attachment receptors. Nat. Struct. Mol. Biol. 2019, 26, 1151–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, Y.; Stevens, D.J.; Skehel, J.J.; Wiley, D.C. X-ray structure of the hemagglutinin of a potential H3 avian progenitor of the 1968 Hong Kong pandemic influenza virus. Virology 2003, 309, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.C.; Otwinowski, J.; Thompson, A.J.; Nycholat, C.M.; Nourmohammad, A.; Wilson, I.A. Major antigenic site B of human influenza H3N2 viruses has an evolving local fitness landscape. Nat. Commun. 2020, 11, 1233. [Google Scholar] [CrossRef]
- Wu, N.C.; Zost, S.J.; Thompson, A.J.; Oyen, D.; Nycholat, C.M.; McBride, R.; Paulson, J.C.; Hensley, S.E.; Wilson, I.A. A structural explanation for the low effectiveness of the seasonal influenza H3N2 vaccine. PLoS Pathog. 2017, 13, e1006682. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.C.; Thompson, A.J.; Xie, J.; Lin, C.W.; Nycholat, C.M.; Zhu, X.; Lerner, R.A.; Paulson, J.C.; Wilson, I.A. A complex epistatic network limits the mutational reversibility in the influenza hemagglutinin receptor-binding site. Nat. Commun. 2018, 9, 1264. [Google Scholar] [CrossRef]
- Clustal Omega. Available online: https://www.ebi.ac.uk/Tools/msa/clustalo/ (accessed on 27 September 2020).
- WHO. WHO Pandemic Phase Descriptions and Main Actions by Phase. Available online: https://www.who.int/influenza/resources/documents/pandemic_phase_descriptions_and_actions.pdf (accessed on 1 September 2020).
- Reid, A.H.; Janczewski, T.A.; Lourens, R.M.; Elliot, A.J.; Daniels, R.S.; Berry, C.L.; Oxford, J.S.; Taubenberger, J.K. 1918 influenza pandemic caused by highly conserved viruses with two receptor-binding variants. Emerg. Infect. Dis. 2003, 9, 1249–1253. [Google Scholar] [CrossRef]
- Xu, R.; McBride, R.; Paulson, J.C.; Basler, C.F.; Wilson, I.A. Structure, receptor binding, and antigenicity of influenza virus hemagglutinins from the 1957 H2N2 pandemic. J. Virol. 2010, 84, 1715–1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Poucke, S.; Doedt, J.; Baumann, J.; Qiu, Y.; Matrosovich, T.; Klenk, H.D.; Van Reeth, K.; Matrosovich, M. Role of substitutions in the hemagglutinin in the emergence of the 1968 pandemic influenza virus. J. Virol. 2015, 89, 12211–12216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vries, R.P.; de Vries, E.; Moore, K.S.; Rigter, A.; Rottier, P.J.; de Haan, C.A. Only two residues are responsible for the dramatic difference in receptor binding between swine and new pandemic H1 hemagglutinin. J. Biol. Chem. 2011, 286, 5868–5875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taubenberger, J.K. The origin and virulence of the 1918 “Spanish” influenza virus. Proc. Am. Philos. Soc. 2006, 150, 86–112. [Google Scholar] [PubMed]
- Lin, Y.P.; Xiong, X.; Wharton, S.A.; Martin, S.R.; Coombs, P.J.; Vachieri, S.G.; Christodoulou, E.; Walker, P.A.; Liu, J.; Skehel, J.J.; et al. Evolution of the receptor binding properties of the influenza A(H3N2) hemagglutinin. Proc. Natl. Acad. Sci. USA 2012, 109, 21474–21479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrasekaran, A.; Srinivasan, A.; Raman, R.; Viswanathan, K.; Raguram, S.; Tumpey, T.M.; Sasisekharan, V.; Sasisekharan, R. Glycan topology determines human adaptation of avian H5N1 virus hemagglutinin. Nat. Biotechnol. 2008, 26, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; de Vries, R.P.; Grant, O.C.; Thompson, A.J.; McBride, R.; Tsogtbaatar, B.; Lee, P.S.; Razi, N.; Wilson, I.A.; Woods, R.J.; et al. Recent H3N2 viruses have evolved specificity for extended, branched human-type receptors, conferring potential for increased avidity. Cell Host Microbe 2017, 21, 23–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tharakaraman, K.; Jayaraman, A.; Raman, R.; Viswanathan, K.; Stebbins, N.W.; Johnson, D.; Shriver, Z.; Sasisekharan, V.; Sasisekharan, R. Glycan receptor binding of the influenza A virus H7N9 hemagglutinin. Cell 2013, 153, 1486–1493. [Google Scholar] [CrossRef] [Green Version]
- Uiprasertkul, M.; Puthavathana, P.; Sangsiriwut, K.; Pooruk, P.; Srisook, K.; Peiris, M.; Nicholls, J.M.; Chokephaibulkit, K.; Vanprapar, N.; Auewarakul, P. Influenza A H5N1 replication sites in humans. Emerg. Infect. Dis. 2005, 11, 1036–1041. [Google Scholar] [CrossRef]
- Shieh, W.J.; Blau, D.M.; Denison, A.M.; Deleon-Carnes, M.; Adem, P.; Bhatnagar, J.; Sumner, J.; Liu, L.; Patel, M.; Batten, B.; et al. 2009 pandemic influenza A (H1N1): Pathology and pathogenesis of 100 fatal cases in the United States. Am. J. Pathol. 2010, 177, 166–175. [Google Scholar] [CrossRef]
- Jia, N.; Byrd-Leotis, L.; Matsumoto, Y.; Gao, C.; Wein, A.N.; Lobby, J.L.; Kohlmeier, J.E.; Steinhauer, D.A.; Cummings, R.D. The human lung glycome reveals novel glycan ligands for influenza A virus. Sci. Rep. 2020, 10, 5320. [Google Scholar] [CrossRef] [PubMed]
- Walther, T.; Karamanska, R.; Chan, R.W.; Chan, M.C.; Jia, N.; Air, G.; Hopton, C.; Wong, M.P.; Dell, A.; Malik Peiris, J.S.; et al. Glycomic analysis of human respiratory tract tissues and correlation with influenza virus infection. PLoS Pathog. 2013, 9, e1003223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Clinical Management of Human Infection with Pandemic (H1N1) 2009: Revised Guidance. Available online: https://www.who.int/csr/resources/publications/swineflu/clinical_management_h1n1.pdf (accessed on 1 September 2020).
- Kuiken, T.; Taubenberger, J.K. Pathology of human influenza revisited. Vaccine 2008, 26 (Suppl. S4), 59–66. [Google Scholar] [CrossRef] [Green Version]
- Rota, P.A.; Wallis, T.R.; Harmon, M.W.; Rota, J.S.; Kendal, A.P.; Nerome, K. Cocirculation of two distinct evolutionary lineages of influenza type B virus since 1983. Virology 1990, 175, 59–68. [Google Scholar] [CrossRef]
- WHO. Summary of Neuraminidase Amino acid Substitutions Associated with Reduced Inhibition by Neuraminidase Inhibitors. Available online: https://www.who.int/influenza/gisrs_laboratory/antiviral_susceptibility/NAI_Reduced_Susceptibility_Marker_Table_WHO.pdf?ua=1 (accessed on 1 September 2020).
- Magesh, S.; Sriwilaijaroen, N.; Moriya, S.; Ando, H.; Miyagi, T.; Suzuki, Y.; Ishida, H.; Kiso, M. Evaluation of a set of C9 N-acyl Neu5Ac2en mimetics as viral sialidase selective inhibitors. Int. J. Med. Chem. 2011, 2011, 539245. [Google Scholar]
- Sriwilaijaroen, N.; Magesh, S.; Imamura, A.; Ando, H.; Ishida, H.; Sakai, M.; Ishitsubo, E.; Hori, T.; Moriya, S.; Ishikawa, T.; et al. A novel potent and highly specific inhibitor against influenza viral N1-N9 neuraminidases: Insight into neuraminidase-inhibitor interactions. J. Med. Chem. 2016, 59, 4563–4577. [Google Scholar] [CrossRef]
- Vavricka, C.J.; Liu, Y.; Kiyota, H.; Sriwilaijaroen, N.; Qi, J.; Tanaka, K.; Wu, Y.; Li, Q.; Li, Y.; Yan, J.; et al. Influenza neuraminidase operates via a nucleophilic mechanism and can be targeted by covalent inhibitors. Nat. Commun. 2013, 4, 1491. [Google Scholar] [CrossRef]
- Sriwilaijaroen, N.; Fukumoto, S.; Kumagai, K.; Hiramatsu, H.; Odagiri, T.; Tashiro, M.; Suzuki, Y. Antiviral effects of Psidium guajava Linn. (guava) tea on the growth of clinical isolated H1N1 viruses: Its role in viral hemagglutination and neuraminidase inhibition. Antivir. Res. 2012, 94, 139–146. [Google Scholar] [CrossRef]
- Sriwilaijaroen, N.; Wilairat, P.; Hiramatsu, H.; Takahashi, T.; Suzuki, T.; Ito, M.; Ito, Y.; Tashiro, M.; Suzuki, Y. Mechanisms of the action of povidone-iodine against human and avian influenza A viruses: Its effects on hemagglutination and sialidase activities. Virol. J. 2009, 6, 124. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.M. Studies on survival of influenza virus between epidemics and antigenic variants of the virus. Am. J. Public Health Nations Health 1949, 39, 171–178. [Google Scholar] [CrossRef]
- Matsuzaki, Y.; Abiko, C.; Mizuta, K.; Sugawara, K.; Takashita, E.; Muraki, Y.; Suzuki, H.; Mikawa, M.; Shimada, S.; Sato, K.; et al. A nationwide epidemic of influenza C virus infection in Japan in 2004. J. Clin. Microbiol. 2007, 45, 783–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homma, M.; Ohyama, S.; Katagiri, S. Age distribution of the antibody to type C influenza virus. Microbiol. Immunol. 1982, 26, 639–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrler, G.; Durkop, I.; Becht, H.; Klenk, H.D. The glycoprotein of influenza C virus is the haemagglutinin, esterase and fusion factor. J. Gen. Virol. 1988, 69 Pt 4, 839–846. [Google Scholar] [CrossRef]
- Wang, M.; Veit, M. Hemagglutinin-esterase-fusion (HEF) protein of influenza C virus. Protein Cell 2016, 7, 28–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrler, G.; Rott, R.; Klenk, H.D.; Muller, H.P.; Shukla, A.K.; Schauer, R. The receptor-destroying enzyme of influenza C virus is neuraminate-O-acetylesterase. EMBO J. 1985, 4, 1503–1506. [Google Scholar] [CrossRef]
- Hause, B.M.; Ducatez, M.; Collin, E.A.; Ran, Z.; Liu, R.; Sheng, Z.; Armien, A.; Kaplan, B.; Chakravarty, S.; Hoppe, A.D.; et al. Isolation of a novel swine influenza virus from Oklahoma in 2011 which is distantly related to human influenza C viruses. PLoS Pathog. 2013, 9, e1003176. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Qi, J.; Khedri, Z.; Diaz, S.; Yu, H.; Chen, X.; Varki, A.; Shi, Y.; Gao, G.F. Correction: An open receptor-binding cavity of hemagglutinin-esterase-fusion glycoprotein from newly-identified influenza D virus: Basis for its broad cell tropism. PLoS Pathog. 2016, 12, e1005505. [Google Scholar] [CrossRef] [Green Version]
- Bardor, M.; Nguyen, D.H.; Diaz, S.; Varki, A. Mechanism of uptake and incorporation of the non-human sialic acid N-glycolylneuraminic acid into human cells. J. Biol. Chem. 2005, 280, 4228–4237. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, K.; Becker, W.B.; Chanock, R.M. Growth in suckling-mouse brain of “IBV-like” viruses from patients with upper respiratory tract disease. Proc. Natl. Acad. Sci. USA 1967, 58, 2268–2273. [Google Scholar] [CrossRef] [Green Version]
- St-Jean, J.R.; Jacomy, H.; Desforges, M.; Vabret, A.; Freymuth, F.; Talbot, P.J. Human respiratory coronavirus OC43: Genetic stability and neuroinvasion. J. Virol. 2004, 78, 8824–8834. [Google Scholar] [CrossRef] [Green Version]
- Woo, P.C.; Lau, S.K.; Chu, C.M.; Chan, K.H.; Tsoi, H.W.; Huang, Y.; Wong, B.H.; Poon, R.W.; Cai, J.J.; Luk, W.K.; et al. Characterization and complete genome sequence of a novel coronavirus, coronavirus HKU1, from patients with pneumonia. J. Virol. 2005, 79, 884–895. [Google Scholar] [CrossRef] [Green Version]
- Decaro, N.; Lorusso, A. Novel human coronavirus (SARS-CoV-2): A lesson from animal coronaviruses. Vet. Microbiol. 2020, 244, 108693. [Google Scholar] [CrossRef] [PubMed]
- Vlasak, R.; Luytjes, W.; Spaan, W.; Palese, P. Human and bovine coronaviruses recognize sialic acid-containing receptors similar to those of influenza C viruses. Proc. Natl. Acad. Sci. USA 1988, 85, 4526–4529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Dong, W.; Milewska, A.; Golda, A.; Qi, Y.; Zhu, Q.K.; Marasco, W.A.; Baric, R.S.; Sims, A.C.; Pyrc, K.; et al. Human coronavirus HKU1 spike protein uses O-acetylated sialic acid as an attachment receptor determinant and employs hemagglutinin-esterase protein as a receptor-destroying enzyme. J. Virol. 2015, 89, 7202–7213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, Y.; Nagao, Y.; Kato, H.; Matsumoto, M.; Nerome, K.; Nakajima, K.; Nobusawa, E. Human influenza A virus hemagglutinin distinguishes sialyloligosaccharides in membrane-associated gangliosides as its receptor which mediates the adsorption and fusion processes of virus infection. Specificity for oligosaccharides and sialic acids and the sequence to which sialic acid is attached. J. Biol. Chem. 1986, 261, 17057–17061. [Google Scholar] [PubMed]
- Tortorici, M.A.; Walls, A.C.; Lang, Y.; Wang, C.; Li, Z.; Koerhuis, D.; Boons, G.J.; Bosch, B.J.; Rey, F.A.; de Groot, R.J.; et al. Structural basis for human coronavirus attachment to sialic acid receptors. Nat. Struct. Mol. Biol. 2019, 26, 481–489. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Tomlinson, A.C.; Wong, A.H.; Zhou, D.; Desforges, M.; Talbot, P.J.; Benlekbir, S.; Rubinstein, J.L.; Rini, J.M. The human coronavirus HCoV-229E S-protein structure and receptor binding. Elife 2019, 8, e51230. [Google Scholar] [CrossRef]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef] [Green Version]
- Oefner, C.; D’Arcy, A.; Mac Sweeney, A.; Pierau, S.; Gardiner, R.; Dale, G.E. High-resolution structure of human apo dipeptidyl peptidase IV/CD26 and its complex with 1-[({2-[(5-iodopyridin-2-yl)amino]-ethyl}amino)-acetyl]-2-cyano-(S)-pyrrolidine. Acta Cryst. 2003, D59, 1206–1212. [Google Scholar] [CrossRef] [Green Version]
- Kirchdoerfer, R.N.; Cottrell, C.A.; Wang, N.; Pallesen, J.; Yassine, H.M.; Turner, H.L.; Corbett, K.S.; Graham, B.S.; McLellan, J.S.; Ward, A.B. Pre-fusion structure of a human coronavirus spike protein. Nature 2016, 531, 118–121. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wu, Y.; Tefsen, B.; Shi, Y.; Gao, G.F. Bat-derived influenza-like viruses H17N10 and H18N11. Trends Microbiol. 2014, 22, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Ciminski, K.; Ran, W.; Gorka, M.; Lee, J.; Malmlov, A.; Schinkothe, J.; Eckley, M.; Murrieta, R.A.; Aboellail, T.A.; Campbell, C.L.; et al. Bat influenza viruses transmit among bats but are poorly adapted to non-bat species. Nat. Microbiol. 2019, 4, 2298–2309. [Google Scholar] [CrossRef] [PubMed]
- Tyrrell, D.A.; Bynoe, M.L. Cultivation of a novel type of common-cold virus in organ cultures. Br. Med. J. 1965, 1, 1467–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamre, D.; Procknow, J.J. A new virus isolated from the human respiratory tract. Proc. Soc. Exp. Biol. Med. 1966, 121, 190–193. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Eckerle, I.; Memish, Z.A.; Liljander, A.M.; Dijkman, R.; Jonsdottir, H.; Juma Ngeiywa, K.J.; Kamau, E.; Younan, M.; Al Masri, M.; et al. Link of a ubiquitous human coronavirus to dromedary camels. Proc. Natl. Acad. Sci. USA 2016, 113, 9864–9869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wentworth, D.E.; Holmes, K.V. Molecular determinants of species specificity in the coronavirus receptor aminopeptidase N (CD13): Influence of N-linked glycosylation. J. Virol. 2001, 75, 9741–9752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walls, A.C.; Tortorici, M.A.; Frenz, B.; Snijder, J.; Li, W.; Rey, F.A.; DiMaio, F.; Bosch, B.J.; Veesler, D. Glycan shield and epitope masking of a coronavirus spike protein observed by cryo-electron microscopy. Nat. Struct. Mol. Biol. 2016, 23, 899–905. [Google Scholar] [CrossRef]
- Yuan, Y.; Cao, D.; Zhang, Y.; Ma, J.; Qi, J.; Wang, Q.; Lu, G.; Wu, Y.; Yan, J.; Shi, Y.; et al. Cryo-EM structures of MERS-CoV and SARS-CoV spike glycoproteins reveal the dynamic receptor binding domains. Nat. Commun. 2017, 8, 15092. [Google Scholar] [CrossRef]
- Corti, D.; Lanzavecchia, A. Broadly neutralizing antiviral antibodies. Annu. Rev. Immunol. 2013, 31, 705–742. [Google Scholar] [CrossRef]
- van der Hoek, L.; Pyrc, K.; Jebbink, M.F.; Vermeulen-Oost, W.; Berkhout, R.J.; Wolthers, K.C.; Wertheim-van Dillen, P.M.; Kaandorp, J.; Spaargaren, J.; Berkhout, B. Identification of a new human coronavirus. Nat. Med. 2004, 10, 368–373. [Google Scholar] [CrossRef]
- Esper, F.; Weibel, C.; Ferguson, D.; Landry, M.L.; Kahn, J.S. Evidence of a novel human coronavirus that is associated with respiratory tract disease in infants and young children. J. Infect. Dis. 2005, 191, 492–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donaldson, E.F.; Haskew, A.N.; Gates, J.E.; Huynh, J.; Moore, C.J.; Frieman, M.B. Metagenomic analysis of the viromes of three North American bat species: Viral diversity among different bat species that share a common habitat. J. Virol. 2010, 84, 13004–13018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynh, J.; Li, S.; Yount, B.; Smith, A.; Sturges, L.; Olsen, J.C.; Nagel, J.; Johnson, J.B.; Agnihothram, S.; Gates, J.E.; et al. Evidence supporting a zoonotic origin of human coronavirus strain NL63. J. Virol. 2012, 86, 12816–12825. [Google Scholar] [CrossRef] [Green Version]
- Corman, V.M.; Muth, D.; Niemeyer, D.; Drosten, C. Hosts and sources of endemic human coronaviruses. Adv. Virus Res. 2018, 100, 163–188. [Google Scholar] [PubMed]
- Xu, R.-H.; He, J.-F.; Evans, M.R.; Peng, G.-W.; Field, H.E.; Yu, D.-W.; Lee, C.-K.; Luo, H.-M.; Lin, W.-S.; Lin, P.; et al. Epidemiologic clues to SARS origin in China. Emerg. Infect. Dis. 2004, 10, 1031. [Google Scholar] [CrossRef] [PubMed]
- Lam, W.K.; Zhong, N.S.; Tan, W.C. Overview on SARS in Asia and the world. Respirology 2003, 8, S2–S5. [Google Scholar] [CrossRef]
- Song, H.D.; Tu, C.C.; Zhang, G.W.; Wang, S.Y.; Zheng, K.; Lei, L.C.; Chen, Q.X.; Gao, Y.W.; Zhou, H.Q.; Xiang, H.; et al. Cross-host evolution of severe acute respiratory syndrome coronavirus in palm civet and human. Proc. Natl. Acad. Sci. USA 2005, 102, 2430–2435. [Google Scholar] [CrossRef] [Green Version]
- Liang, G.; Chen, Q.; Xu, J.; Liu, Y.; Lim, W.; Peiris, J.S.M.; Anderson, L.J.; Ruan, L.; Li, H.; Kan, B.; et al. Laboratory diagnosis of four recent sporadic cases of community-acquired SARS, Guangdong province, China. Emerg. Infect. Dis. 2004, 10, 1774. [Google Scholar] [CrossRef]
- WHO. Severe Acute Respiratory Syndrome (SARS). Available online: https://www.who.int/csr/don/archive/disease/severe_acute_respiratory_syndrome/en/ (accessed on 1 September 2020).
- Guan, Y.; Zheng, B.J.; He, Y.Q.; Liu, X.L.; Zhuang, Z.X.; Cheung, C.L.; Luo, S.W.; Li, P.H.; Zhang, L.J.; Guan, Y.J.; et al. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science 2003, 302, 276–278. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.F.; Shi, Z.; Zhang, S.; Field, H.; Daszak, P.; Eaton, B.T. Review of bats and SARS. Emerg. Infect. Dis. 2006, 12, 1834–1840. [Google Scholar] [CrossRef]
- Poon, L.L.; Chu, D.K.; Chan, K.H.; Wong, O.K.; Ellis, T.M.; Leung, Y.H.; Lau, S.K.; Woo, P.C.; Suen, K.Y.; Yuen, K.Y.; et al. Identification of a novel coronavirus in bats. J. Virol. 2005, 79, 2001–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Shi, Z.; Yu, M.; Ren, W.; Smith, C.; Epstein, J.H.; Wang, H.; Crameri, G.; Hu, Z.; Zhang, H.; et al. Bats are natural reservoirs of SARS-like coronaviruses. Science 2005, 310, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Hou, Y.; Shen, J.; Huang, Y.; Martin, W.; Cheng, F. Network-based drug repurposing for novel coronavirus 2019-nCoV/SARS-CoV-2. Cell Discov. 2020, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Gandhi, R.T.; Lynch, J.B.; del Rio, C. Mild or moderate Covid-19. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- WHO. Coronavirus Disease (COVID-19) Weekly Epidemiological Update and Weekly Operational Update. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports/?gclid=EAIaIQobChMIsdzRkq3o6QIV1zMrCh2kNA1REAAYASACEgJFnPD_BwE (accessed on 1 September 2020).
- Lau, S.K.P.; Luk, H.K.H.; Wong, A.C.P.; Li, K.S.M.; Zhu, L.; He, Z.; Fung, J.; Chan, T.T.Y.; Fung, K.S.C.; Woo, P.C.Y. Possible bat origin of severe acute respiratory syndrome coronavirus 2. Emerg. Infect. Dis. 2020, 26, 1542. [Google Scholar] [CrossRef]
- Ji, W.; Wang, W.; Zhao, X.; Zai, J.; Li, X. Cross-species transmission of the newly identified coronavirus 2019-nCoV. J. Med. Virol. 2020, 92, 433–440. [Google Scholar] [CrossRef]
- Xia, X. Extreme genomic CpG deficiency in SARS-CoV-2 and evasion of host antiviral defense. Mol. Biol. Evol. 2020. [Google Scholar] [CrossRef]
- Mallapaty, S. Coronavirus can infect cats—Dogs, not so much. Nature 2020. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS–coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, T.T.; Jia, N.; Zhang, Y.W.; Shum, M.H.; Jiang, J.F.; Zhu, H.C.; Tong, Y.G.; Shi, Y.X.; Ni, X.B.; Liao, Y.S.; et al. Identifying SARS-CoV-2-related coronaviruses in Malayan pangolins. Nature 2020, 583, 282–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, K.; Li, W.; Peng, G.; Li, F. Crystal structure of NL63 respiratory coronavirus receptor-binding domain complexed with its human receptor. Proc. Natl. Acad. Sci. USA 2009, 106, 19970–19974. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Li, W.; Farzan, M.; Harrison, S.C. Structure of SARS coronavirus spike receptor-binding domain complexed with receptor. Science 2005, 309, 1864–1868. [Google Scholar] [CrossRef]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Sui, J.; Huang, I.C.; Kuhn, J.H.; Radoshitzky, S.R.; Marasco, W.A.; Choe, H.; Farzan, M. The S proteins of human coronavirus NL63 and severe acute respiratory syndrome coronavirus bind overlapping regions of ACE2. Virology 2007, 367, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.; Chen, L.; Peng, G.; Zhou, W.; Pennell, C.A.; Mansky, L.M.; Geraghty, R.J.; Li, F. A virus-binding hot spot on human angiotensin-converting enzyme 2 is critical for binding of two different coronaviruses. J. Virol. 2011, 85, 5331–5337. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhang, C.; Sui, J.; Kuhn, J.H.; Moore, M.J.; Luo, S.; Wong, S.K.; Huang, I.C.; Xu, K.; Vasilieva, N.; et al. Receptor and viral determinants of SARS-coronavirus adaptation to human ACE2. EMBO J. 2005, 24, 1634–1643. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Zhao, Y.B.; Wang, Q.; Li, J.Y.; Zhou, Z.J.; Liao, C.H.; Ge, X.Y. Predicting the angiotensin converting enzyme 2 (ACE2) utilizing capability as the receptor of SARS-CoV-2. Microbes Infect. 2020, 22, 221–225. [Google Scholar] [CrossRef]
- Luan, J.; Lu, Y.; Jin, X.; Zhang, L. Spike protein recognition of mammalian ACE2 predicts the host range and an optimized ACE2 for SARS-CoV-2 infection. Biochem. Biophys. Res. Commun. 2020, 526, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Greenough, T.C.; Moore, M.J.; Vasilieva, N.; Somasundaran, M.; Sullivan, J.L.; Farzan, M.; Choe, H. Efficient replication of severe acute respiratory syndrome coronavirus in mouse cells is limited by murine angiotensin-converting enzyme 2. J. Virol. 2004, 78, 11429–11433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata, N.; Iwata, N.; Hasegawa, H.; Fukushi, S.; Yokoyama, M.; Harashima, A.; Sato, Y.; Saijo, M.; Morikawa, S.; Sata, T. Participation of both host and virus factors in induction of severe acute respiratory syndrome in F344 rats infected with SARS coronavirus. J. Virol. 2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, X.; Chen, J.; Wang, X.; Zhang, F.; Liu, Y. Age- and gender-related difference of ACE2 expression in rat lung. Life Sci. 2006, 78, 2166–2171. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, X.; Liu, W.; Gan, M.; Zhang, L.; Wang, J.; Zhang, Z.; Zhu, A.; Li, F.; Sun, J.; et al. Discovery of a subgenotype of human coronavirus NL63 associated with severe lower respiratory tract infection in China, 2018. Emerg. Microbes Infect. 2020, 9, 246–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.; Fouchier, R.A. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef]
- de Groot, R.J.; Baker, S.C.; Baric, R.S.; Brown, C.S.; Drosten, C.; Enjuanes, L.; Fouchier, R.A.; Galiano, M.; Gorbalenya, A.E.; Memish, Z.A.; et al. Middle East respiratory syndrome coronavirus (MERS-CoV): Announcement of the coronavirus study group. J. Virol. 2013, 87, 7790–7792. [Google Scholar] [CrossRef] [Green Version]
- Smits, S.L.; Raj, V.S.; Pas, S.D.; Reusken, C.B.; Mohran, K.; Farag, E.A.; Al-Romaihi, H.E.; AlHajri, M.M.; Haagmans, B.L.; Koopmans, M.P. Reliable typing of MERS-CoV variants with a small genome fragment. J. Clin. Virol. 2015, 64, 83–87. [Google Scholar] [CrossRef] [Green Version]
- Raj, V.S.; Mou, H.; Smits, S.L.; Dekkers, D.H.W.; Müller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.A.A.; Zaki, A.; Fouchier, R.A.M.; et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 2013, 495, 251–254. [Google Scholar] [CrossRef] [Green Version]
- Shang, J.; Wan, Y.; Liu, C.; Yount, B.; Gully, K.; Yang, Y.; Auerbach, A.; Peng, G.; Baric, R.; Li, F. Structure of mouse coronavirus spike protein complexed with receptor reveals mechanism for viral entry. PLoS Pathog. 2020, 16, e1008392. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Shi, X.; Jiang, L.; Zhang, S.; Wang, D.; Tong, P.; Guo, D.; Fu, L.; Cui, Y.; Liu, X.; et al. Structure of MERS-CoV spike receptor-binding domain complexed with human receptor DPP4. Cell Res. 2013, 23, 986–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIE. Avian Influenza Vaccination. Available online: https://www.oie.int/doc/ged/D4410.PDF (accessed on 1 September 2020).
- Yang, L.; Zhao, X.; Li, X.; Bo, H.; Li, D.; Liu, J.; Wang, D. Case report for human infection with a highly pathogenic avian influenza A(H5N6) virus in Beijing, China 2019. Biosaf. Health 2020, 2, 49–52. [Google Scholar] [CrossRef]
- Yu, D.; Xiang, G.; Zhu, W.; Lei, X.; Li, B.; Meng, Y.; Yang, L.; Jiao, H.; Li, X.; Huang, W.; et al. The re-emergence of highly pathogenic avian influenza H7N9 viruses in humans in mainland China, 2019. Eurosurveillance 2019, 24, 1900273. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Guan, L.; Shi, J.; Kong, X.; Ma, S.; Zhang, Y.; Yin, X.; He, X.; Liu, L.; Suzuki, Y.; Li, C.; et al. H3N2 avian influenza viruses detected in live poultry markets in China bind to human-type receptors and transmit in guinea pigs and ferrets. Emerg. Microbes Infect. 2019, 8, 1280–1290. [Google Scholar] [CrossRef] [Green Version]
- Sriwilaijaroen, N.; Suzuki, Y. A simple viral neuraminidase-based detection for high-throughput screening of viral hemagglutinin-host receptor specificity. Methods Mol. Biol. 2014, 1200, 107–120. [Google Scholar] [PubMed]
- Stevens, J.; Blixt, O.; Paulson, J.C.; Wilson, I.A. Glycan microarray technologies: Tools to survey host specificity of influenza viruses. Nat. Rev. Microbiol. 2006, 4, 857–864. [Google Scholar] [CrossRef]
- Nakakita, S.I.; Sriwilaijaroen, N.; Suzuki, Y.; Hirabayashi, J. Preparation and detection of glycan-binding activity of influenza virus. Methods Mol. Biol. 2020, 2132, 567–583. [Google Scholar]
- Watanabe, Y.; Ito, T.; Ibrahim, M.S.; Arai, Y.; Hotta, K.; Phuong, H.V.M.; Hang, N.L.K.; Mai, L.Q.; Soda, K.; Yamaoka, M.; et al. A novel immunochromatographic system for easy-to-use detection of group 1 avian influenza viruses with acquired human-type receptor binding specificity. Biosens. Bioelectron. 2015, 65, 211–219. [Google Scholar] [CrossRef]
- Sun, H.; Xiao, Y.; Liu, J.; Wang, D.; Li, F.; Wang, C.; Li, C.; Zhu, J.; Song, J.; Sun, H.; et al. Prevalent Eurasian avian-like H1N1 swine influenza virus with 2009 pandemic viral genes facilitating human infection. Proc. Natl. Acad. Sci. USA 2020, 117, 17204–17210. [Google Scholar] [CrossRef] [PubMed]
- Rajao, D.S.; Gauger, P.C.; Anderson, T.K.; Lewis, N.S.; Abente, E.J.; Killian, M.L.; Perez, D.R.; Sutton, T.C.; Zhang, J.; Vincent, A.L. Novel reassortant human-like H3N2 and H3N1 influenza A viruses detected in pigs are virulent and antigenically distinct from swine viruses endemic to the United States. J. Virol. 2015, 89, 11213–11222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, Y.; Ibrahim, M.S.; Ellakany, H.F.; Kawashita, N.; Mizuike, R.; Hiramatsu, H.; Sriwilaijaroen, N.; Takagi, T.; Suzuki, Y.; Ikuta, K. Acquisition of human-type receptor binding specificity by new H5N1 influenza virus sublineages during their emergence in birds in Egypt. PLoS Pathog. 2011, 7, e1002068. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Deng, G.; Kong, H.; Gu, C.; Ma, S.; Yin, X.; Zeng, X.; Cui, P.; Chen, Y.; Yang, H.; et al. H7N9 virulent mutants detected in chickens in China pose an increased threat to humans. Cell Res. 2017, 27, 1409–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, T.; Lim, W.; Suzuki, T.; Suzuki, Y.; Kida, H.; Nishimura, S.I.; Tashiro, M. Characterization of a human H9N2 influenza virus isolated in Hong Kong. Vaccine 2001, 20, 125–133. [Google Scholar] [CrossRef]
- Belser, J.A.; Blixt, O.; Chen, L.M.; Pappas, C.; Maines, T.R.; Van Hoeven, N.; Donis, R.; Busch, J.; McBride, R.; Paulson, J.C.; et al. Contemporary North American influenza H7 viruses possess human receptor specificity: Implications for virus transmissibility. Proc. Natl. Acad. Sci. USA 2008, 105, 7558–7563. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Siddique, R.; Shereen, M.A.; Ali, A.; Liu, J.; Bai, Q.; Bashir, N.; Xue, M. Emergence of a novel coronavirus, severe acute respiratory syndrome coronavirus 2: Biology and therapeutic options. J. Clin. Microbiol. 2020, 58. [Google Scholar] [CrossRef] [Green Version]
- Azhar, E.I.; El-Kafrawy, S.A.; Farraj, S.A.; Hassan, A.M.; Al-Saeed, M.S.; Hashem, A.M.; Madani, T.A. Evidence for camel-to-human transmission of MERS coronavirus. N. Engl. J. Med. 2014, 370, 2499–2505. [Google Scholar] [CrossRef]
- Li, X.; Giorgi, E.E.; Marichannegowda, M.H.; Foley, B.; Xiao, C.; Kong, X.-P.; Chen, Y.; Gnanakaran, S.; Korber, B.; Gao, F. Emergence of SARS-CoV-2 through recombination and strong purifying selection. Sci. Adv. 2020, 6, eabb9153. [Google Scholar] [CrossRef]
- Sriwilaijaroen, N.; Suzuki, Y. Hemagglutinin inhibitors are potential future anti-influenza drugs for mono- and combination therapies. Methods Mol. Biol. 2020, 2132, 547–565. [Google Scholar]
- Sriwilaijaroen, N.; Suzuki, K.; Takashita, E.; Hiramatsu, H.; Kanie, O.; Suzuki, Y. 6SLN-lipo PGA specifically catches (coats) human influenza virus and synergizes neuraminidase-targeting drugs for human influenza therapeutic potential. J. Antimicrob. Chemother. 2015, 70, 2797–2809. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Genus | Genome | Type | Host | IAV Subtype or IBV1/ICV/IDV Lineage | Disease | Receptor-binding Protein | Host Receptor | Ref. | |
|---|---|---|---|---|---|---|---|---|---|---|
| Birds | Wild birds (frequently found in waterfowls and shorebirds) | H1–H16 and N1–N9 HP: H5NX, X = NA subtypes | No or mild disease HP: isolated from dead wild birds | HA | Terminal Neu5Acα2,3Gal Some gull/tern H16: α2,6Neu5Ac ≥ α2,3Neu5Ac Internal Duck type: non-fucosylate, SiaLec, 3′SLN Gull type: fucosylated, SiaLex and 6-sulfo-SiaLex Gull/tern H16: 6′SLDN ≥ LSTb, but some 6-sulfo-SiaLex > 6′SLDN, LSTb | [23,24] | ||||
| Domestic birds (poultry) | H1–H13, N1–N9 | LP: mild disease HP: multi-organ systemic disease | HA | Terminal Neu5Acα2,3Gal Internal HP higher affinity than LP: 3′SLN, SiaLec All LP and HP: 3′SLN > SiaLex, SiaLec > SiaLea, 3′SLN ~ SiaLec H5 poultry: 6-sulfo-3′SLN H7H7 from all species: 6-sulfo-3′SLN, 6-sulfo-SiaLex > nonsulfated SGPs | [25,26,27,28,29,30,31,32,33,34,35,36,37] | |||||
| Domestic ducks | HP: H5N1, H5N2, H5N3, H5N5, H5N6, H5N8, H7N2, H7N9 | |||||||||
| Other poultry | HP: H5N1, H5N2, H5N3, H5N6, H5N8, H5N9, H7N1, H7N2, H7N3, H7N4, H7N7, H7N9 | |||||||||
| Alphainfluenzavirus | Other animals | Pigs | H1N1, H1N2, H2N3, H3N1, H3N2, HPH5N1 | Respiratory disease | HA | Neu5Acα2,6Gal > Neu5Acα2,3Gal Neu5AcGM3, Neu5GcGM3 | [4] [38,39,40,41] | |||
| Horses | H7N7, H3N8 | Respiratory disease | HA | Mainly Neu5Gcα2,3Gal, Sialyllacto-N-tetraose a, SiaLex | [42,43] | |||||
| 8 (−)ssRNA segments | Dogs | H3N8, H3N2, H3N1, H5N2, HPAI H5N1, H1N1 | Respiratory disease | HA | Canine H3N8: 6-sulfo-3′SLN and Neu5Acα2,3Galβ- | [38] [43,44,45,46] | ||||
| Cats | H7N2, H3N2, HPH5N1 | Respiratory disease | HA | Feline H7N2: Siaα2,3Gal | [38,47] [48] | |||||
| A | Ferrets (a model for human infection and transmission) | Triple reassortant H1N1 SIV, A(H1N1)pdm09 | Respiratory disease | HA | Expected to bind to Neu5Acα2,6Gal | [49,50] | ||||
| Occasionally in numerous species of mammals such as whales, seals, tigers, and minks | Seal H7N7 & whale H13N9 Tiger HPH5N1 Mink H9N2 | Respiratory disease | HA | Seal H7N7 & whale H13N9: Siaα2,3Gal Tiger HPH5N1 HA with Q226 and G228 (H3 numbering): predicted to prefer Siaα2,3Gal Mink H9N2 HA with 226L: increased binding to Siaα2,6Gal | [51,52,53,54,55,56] | |||||
| Orthomyxoviridae | Zoonosis | Poultry, pigs and other animals | H1, H2, H3, H5 (HP & LP), H6, H7 (HP & LP), H9, H10 | HP: severe systemic disease Other zoonotic infections: ranging from mild upper respiratory tract infection to severe pneumonia and death. Possible to cause conjunctivitis, GI symptoms, encephalitis and encephalopathy | HA | Most still retains previous host-receptor specificity | [16] [57,58,59] | |||
| Humans | 1918-derived H1N1 (extinct), 1957-derived H2N2 (extinct), 1968-derived H3N2, 2009-derived H1N1 | Pandemic and continued as seasonal flu: Primarily cause respiratory disease (usually pdm more severe, including pneumonia) | HA | Terminal Neu5Acα2,6Gal Internal 6′S(LN)n Early human-adapted viruses: n = 1, 2, 3, etc. Long-term circulating viruses: n ≥ 3 | [16,19] [60,61] | |||||
| Bats | Fruit-eating bats in Guatemala | Sl-BatH17N10 Fruit-eating bats in Peru: Ap-batH18N11 | H17: liver, intestine, lung and kidney tissues, and rectal swabs H18: intestine tissue and rectal swabs | HA | MHC-II HLA-DR | [62,63] | ||||
| Fruit-eating bats in Egypt | Distinct from all known IAVs. HAs, closest to mallard H9 viruses with 73% aa identity | Viruses in bat oral swabs > in rectal swabs Able to replicate in mouse lungs | HA | Terminal Neu5Acα2,3Gal > Neu5Acα2,6Gal | [64] | |||||
| Betainfluenzavirus | 8 (-)ssRNA segments | B | Humans, sporadically in seals, pigs, horses, pheasants and dogs. | Influenza B viruses (IBVs) 2 lineages - Victoria lineage B/Victoria/2/87-like viruses - Yamagata lineage: B/Yamagata/16/88-like viruses | Seasonal flu | HA | Wild types of both Yamagata (HA with F95 and N194) and Victoria (HA with G141, R162 and D196) lineages: Neu5Acα2,6Gal>Neu5Acα2,3Gal 2001–2007 clinical isolates in Taiwan: 83% of Yamagata-like strains prefer α2,6Sia; 17% prefer both α2,3Sia and α2,6Sia. 54% of Victoria-like strains prefer both α2,3Sia and α2,6Sia; 25% prefer sulfated glycan, either β-Gal-3-sulfate or 6-HSO3-Galβ1,4GlcNAc; 21% prefer α2,6Sia. Dual α2,3Sia and α2,6Sia-binding viruses seem to be associated with bronchopneumonia and gastrointestinal symptoms. | [65,66,67,68] | ||
| Gammainfluenzavirus | 7 (−)ssRNA segments | C | Humans (particularly in infants and children), pigs, cattle | Influenza C viruses (ICVs) 6 lineages: C/Taylor, C/Mississippi, C/Aichi, C/Yamagata, C/Kanagawa and C/Sao Paulo | Usually mild flu | HEF | Terminal C/Johannesburg/1/66 (C/Aichi lineage): Neu5,9Ac2α2,6Gal | [69,70,71,72] | ||
| Deltainfluenzavirus | 7 (−)ssRNA segments | D | Pigs, cattle, 91% seropositivity found among persons working with cattle | Influenza D viruses (IDVs) 3 lineages: D/OK, D/660 and D/Japanese D/Japanese lineage contains 2 sublineages: D/Yama2016 and D/Yama2019 | Usually mild flu | HEF | Terminal D/OK: Neu5,9Ac2 and Neu5Gc9Ac either linked to α2,6Gal or α2,3Gal D/660: Neu5,9Ac2α2,6Gal, Neu5Gc9Acα2,6Gal and Neu5Gc9Acα2,3Gal | [15,72] [73] | ||
| Family | Subfamily | Genus | Genome | Lineage | Host | CoV | Disease | Receptor-binding Protein | Host Receptor | Ref. | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Primary Receptor | Attachment Factor | ||||||||||
| Alphacoronavirus (gr. 1) | Single linear (+)ssRNA | Bats | Ro(HKU10)-, Hi(HKU10)-, Rh(HKU2)-, Sc(512)-, Mi(1A, 1B, HKU8)- BatCoV | Unknown | [74] | ||||||
| Other animals | FeCoV I & II (nonvirulent form of FCoV), CCoV, TGEV, PEDV | Gastroenteritis | S1 | S1B (S1-CTD) Aminopeptidase N (APN; CD13), not for PEDV and FeCoV I | S1A (S1-NTD) α2,3Neu5Gc >Neu5Ac (TGEV traveling to a target site), Neu5Ac > Neu5Gc (PEDV) | [75,76,77,78,79,80] | |||||
| FIPV (virulent form of FCoV) | Peritonitis, severe wasting | S1 | fAPN for type II, not type I FIPV | Coreceptor for both types I and II: DC-SIGN | [81,82] | ||||||
| PRCoV | Respiratory disease | S1-CTD | pAPN (res. 366–369, 727–790) | [83] | |||||||
| Humans | HCoV-229E, HCoV-NL63 | Generally URTI | S1-CTD | NL63: hACE2 229E: hAPN (res. 208, 241–319) | [84] | ||||||
| Rodents | ChRCoV (HKU24) | Unknown | [85] | ||||||||
| RCoV | Respiratory disease, sialodacryoadenitis | HE | Unknown | HE (type II) 4-O-Ac-Sias, 4,5-di-N-acetylneuraminic acid α-methylglycoside (α-4-N-Ac-Sia) | [86,87] | ||||||
| MHV | Gastroenteritis, hepatitis, encephalomyelitis | S1-NTD, HE | S1-NTD Murine CEACAM1 | HE Strain DVIM: 9-O-Ac-Sias Strains S, JHM: 4-O-Ac-Sias | [88,89] | ||||||
| A | Other animals | BCoV, EqCoV(ECoV) | Enteritis, respiratory disease | S1-NTD, HE | S1-NTD 9-O-acetylated sialoglycans (unknown for EqCoV) | HE (type I) 9-O-acetylated sialoglycans, 7,9-di-O-acetyl Sia | [86,90] | ||||
| PHEV | Vomiting, wasting, encephalomyelitis | S1, HE | S1 (res. 291–548) Porcine NCAM S1-NTD 9-O-Ac-Sia | HE 9-O-Ac-Sias | [91,92,93] | ||||||
| Humans | HCoV-OC43, HCoV-HKU1 | Generally URTI | S1-NTD, HE | S1-NTD Neu5,9Ac2 | HE Neu5,9Ac2 Progressive loss of binding during circulation in humans OC43: accumulation of mutations HKU1: massive deletions | [92,94] | |||||
| Bats | SARSr-Rh(HKU3)-, SARSr-Rp-, SARSr-Rs-, SARSr-Rf-BatCoV, Bat-SL-RaTG13, Bat-SL-RmYN02 | Viruses in anal/fecal swabs | RsACE2, 3 aa substitutions in RpACE2: shown to be used by SARS-CoV | [95,96,97] | |||||||
| Betacoronavirus (gr. 2) | Single linear (+)ssRNA | Other animals | Masked palm civet, raccoon dog: SARS-CoV-like virus SARS-CoV real-time RT-PCR-positive animals: red fox, Sikkim rat, wild boar, etc. Pangolin: SARS-CoV-2-like CoV SARS-CoV-2 RT-PCR-positive animals: dog, cat, tiger, etc. | Palm civets: viruses in rectal and/or throat swabs Pangolins: viruses from lung tissues Domestic cats, dogs: viruses in oral/nasal specimens | S1-CTD | Host ACE2 Prototype: viral S gene has no SNV. | [98,99,100,101,102,103] | ||||
| Coronaviridae | Orthocoronavirinae | B | Zoonosis | SARS-CoV | SARS in humans: atypical pneumonia (type I pneumocytes), ARDS, gastroenteritis | S1 | S1-CTD hACE2 - Nov 2002–Jan 2003 (early epidemic, HP, animal/human-to-human by close contact): viral S gene contains 17–22 SNVs. - Feb–Jul 2003 (epidemic, HP, human-to-human by close contact): 25–27 SNVs - Dec 2003–May 2004 (re-appearance, LP, animal-to-human): 2–7 SNVs | DC-SIGN, DC-SIGNR and LSECtin: their roles are less clear. | [100] [104] [105] | ||
| Pandemic | SARS-CoV-2 | COVID-19: URTI, LRTI (non-life-threatening pneumonia, severe pneumonia with ARDS) | S1-CTD | hACE2 (efficient and sustained spread among humans) | [9] | ||||||
| Bats | Ty(HKU4)-, Hp(HKU25)-, Pi(HKU5)-BatCoV, NeoCoV | Virus in feces | S1 | S1-CTD Ty- and Hp-BatCoVs, but not Pi-BatCoV, can bind to hDPP4. | S1-NTD Binds to the intestinal epithelium in common pipistrelle bats but not in serotine bats and frugivorous bats. | [106,107,108] | |||||
| C | Dromedary camels | dromedary MERS-CoV | Virus/RNA in nasal swabs and lung tissue samples. An experimental study: mild upper respiratory tract disease | S1 | S1-CTD dromedary DDP4 found in various organs, particularly in nasal turbinate, trachea and lungs | S1-NTD α2,3-sialic acid The S1A domain binds to the dromedary nasal epithelium but not to the porcine/rabbit nasal epithelium. | [108,109,110,111] | ||||
| Zoonosis | MERS-CoV | Atypical pneumonia | S1 | S1-CTD hDDP4 (CD26) Note: Intercellular spread of the virus through cell-cell fusion does not require DDP4 receptors but requires TMPRSS2 activity. | S1-NTD Short and long α2,3Neu5Ac > α2,6Neu5Ac glycans The S1A domain binds to the human alveolar epithelium (type II pneumocytes). | [104] [108] [112] [113] | |||||
| D | Bats | Ro-BatCoV (HKU9) | Unknown | [74] | |||||||
| Gammacoronavirus (gr.3) | Single linear (+)ssRNA | Birds | Infectious bronchitis virus (IBV2) | Bronchitis, nephritis, reproductive problems | S1-NTD | IBV: α2,3-linked sialic acids type 1 lactosamines IBV strain Beaudette: heparan sulfate | [114] [115] | ||||
| TuCoV(TCoV), GfCoV | Enteritis | S1 | Nonsialylated type 2 poly-N-acetyl-lactosamines | [114] | |||||||
| Deltacoronavirus (a new group) | Single linear (+)ssRNA | Birds | FalCoV, HouCoV, PiCoV | Unknown | [116] | ||||||
| Other animals | PorCoV (PDCoV) | Diarrhea in newborn piglets | S1 | pAPN in porcine alveolar macrophages but not necessary for infection of lung-derived fibroblast cells | [77] | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sriwilaijaroen, N.; Suzuki, Y. Host Receptors of Influenza Viruses and Coronaviruses—Molecular Mechanisms of Recognition. Vaccines 2020, 8, 587. https://doi.org/10.3390/vaccines8040587
Sriwilaijaroen N, Suzuki Y. Host Receptors of Influenza Viruses and Coronaviruses—Molecular Mechanisms of Recognition. Vaccines. 2020; 8(4):587. https://doi.org/10.3390/vaccines8040587
Chicago/Turabian StyleSriwilaijaroen, Nongluk, and Yasuo Suzuki. 2020. "Host Receptors of Influenza Viruses and Coronaviruses—Molecular Mechanisms of Recognition" Vaccines 8, no. 4: 587. https://doi.org/10.3390/vaccines8040587
APA StyleSriwilaijaroen, N., & Suzuki, Y. (2020). Host Receptors of Influenza Viruses and Coronaviruses—Molecular Mechanisms of Recognition. Vaccines, 8(4), 587. https://doi.org/10.3390/vaccines8040587