Immunogenicity and Protection against Mycobacterium caprae Challenge in Goats Vaccinated with BCG and Revaccinated after One Year

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Vaccination Schedule and Experimental Infection
2.2. Ethics Statement
2.3. In Vitro IFN-γ Release Assay (IGRA)
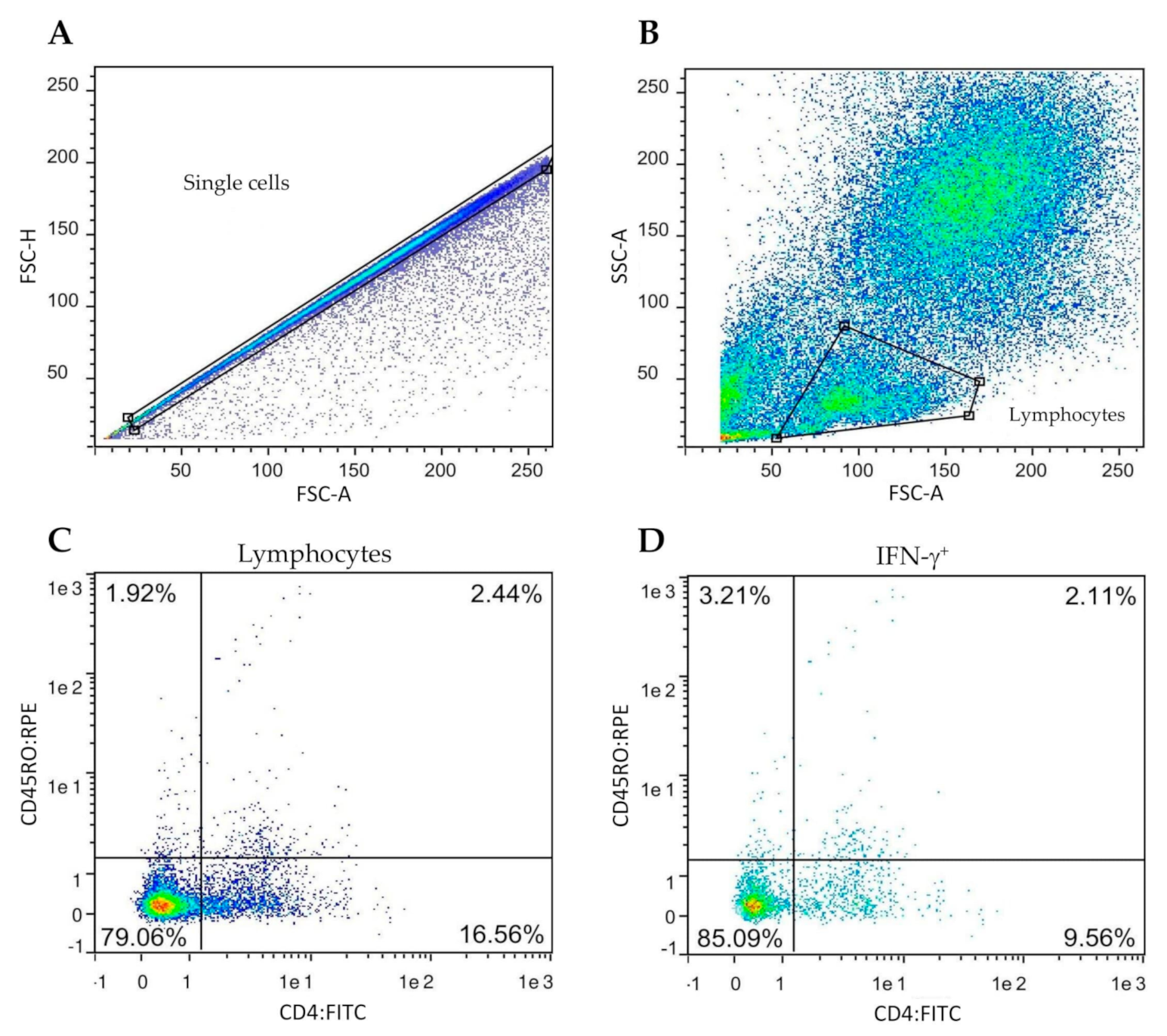
2.4. Flow Cytometry and Intracellular IFN-γ Staining
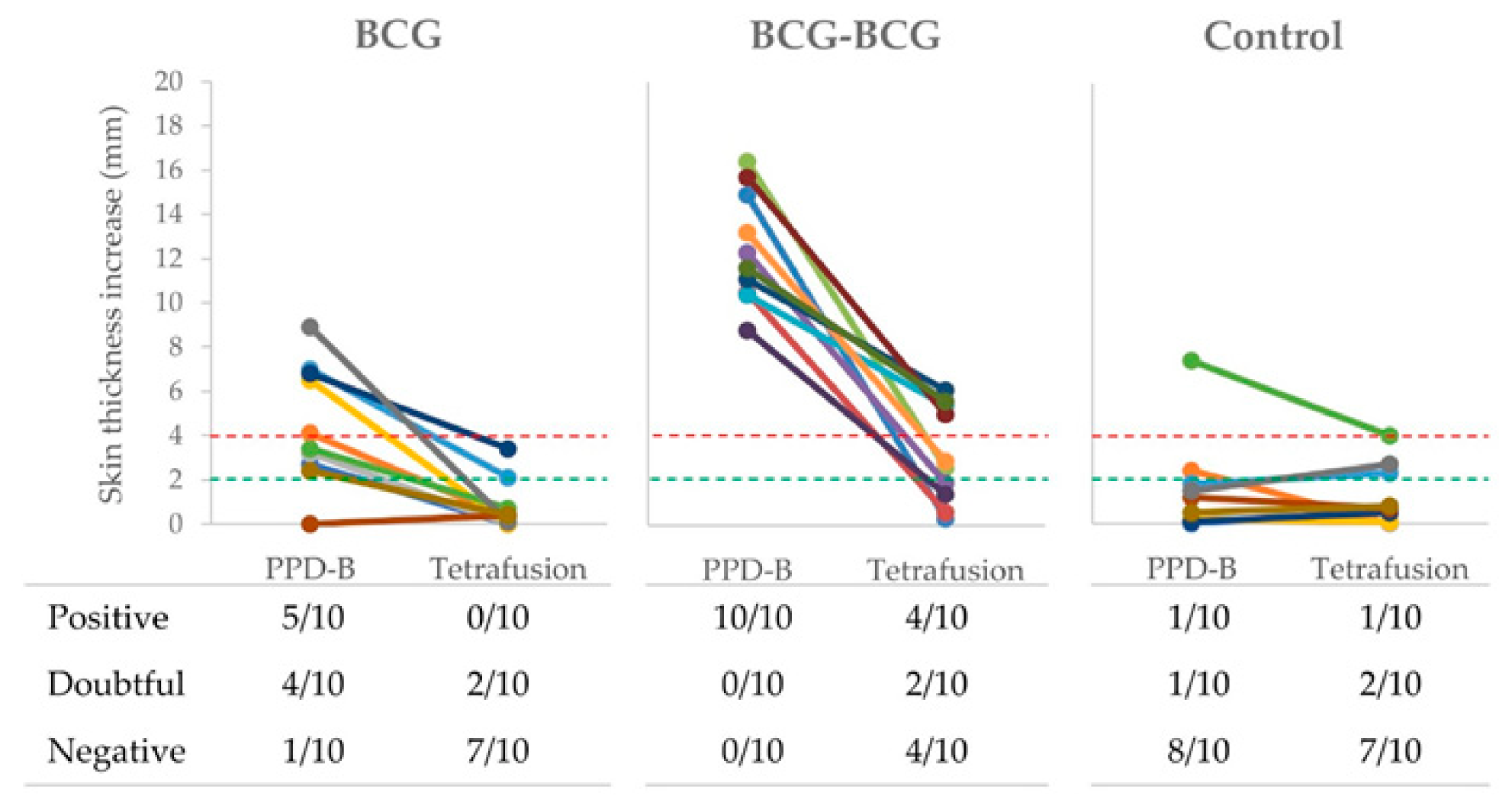
2.5. Skin Tests
2.6. Post-Mortem Examination
2.7. M. caprae DNA Burden Assessment by qPCR
2.8. Data Analysis
3. Results
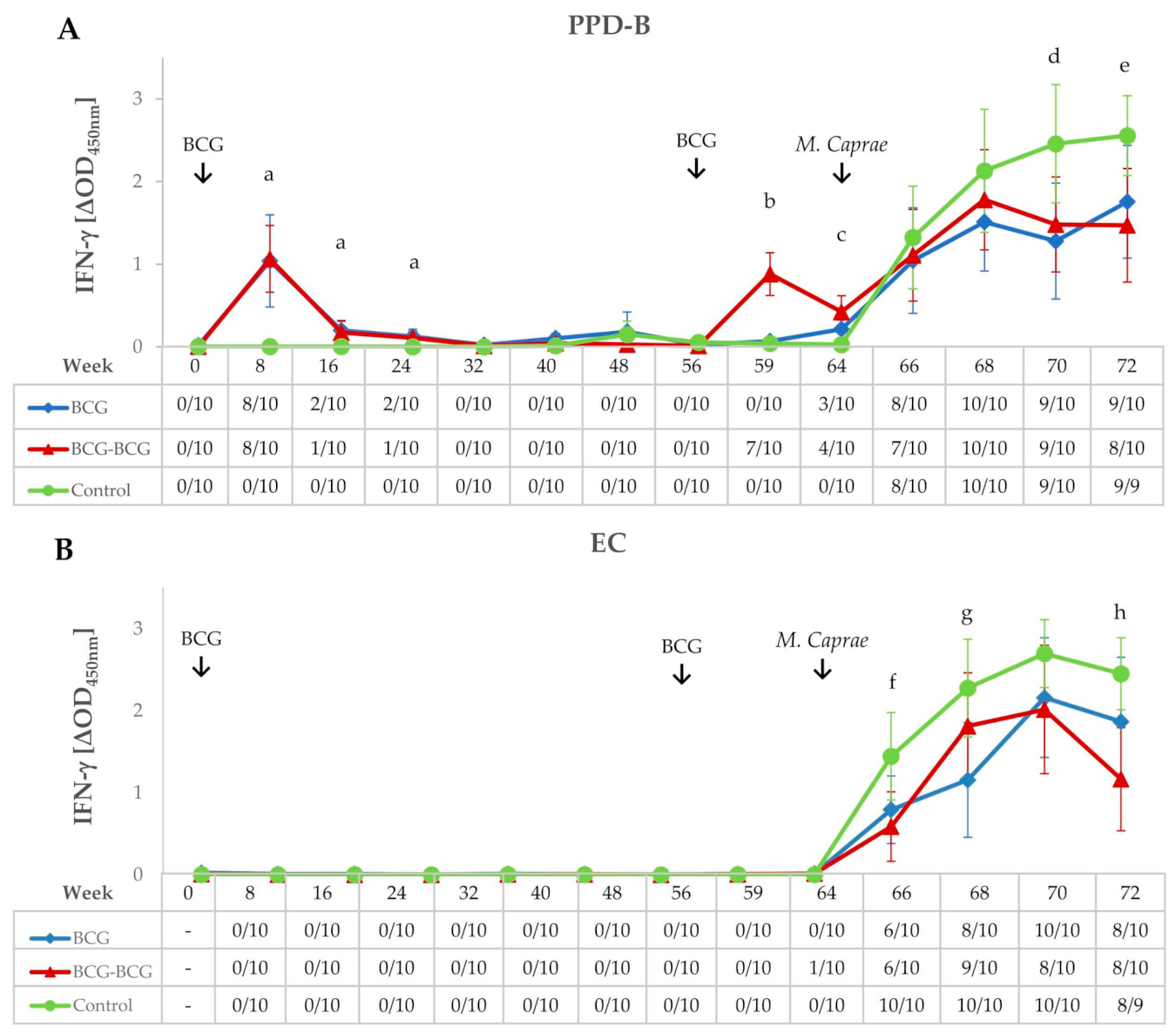
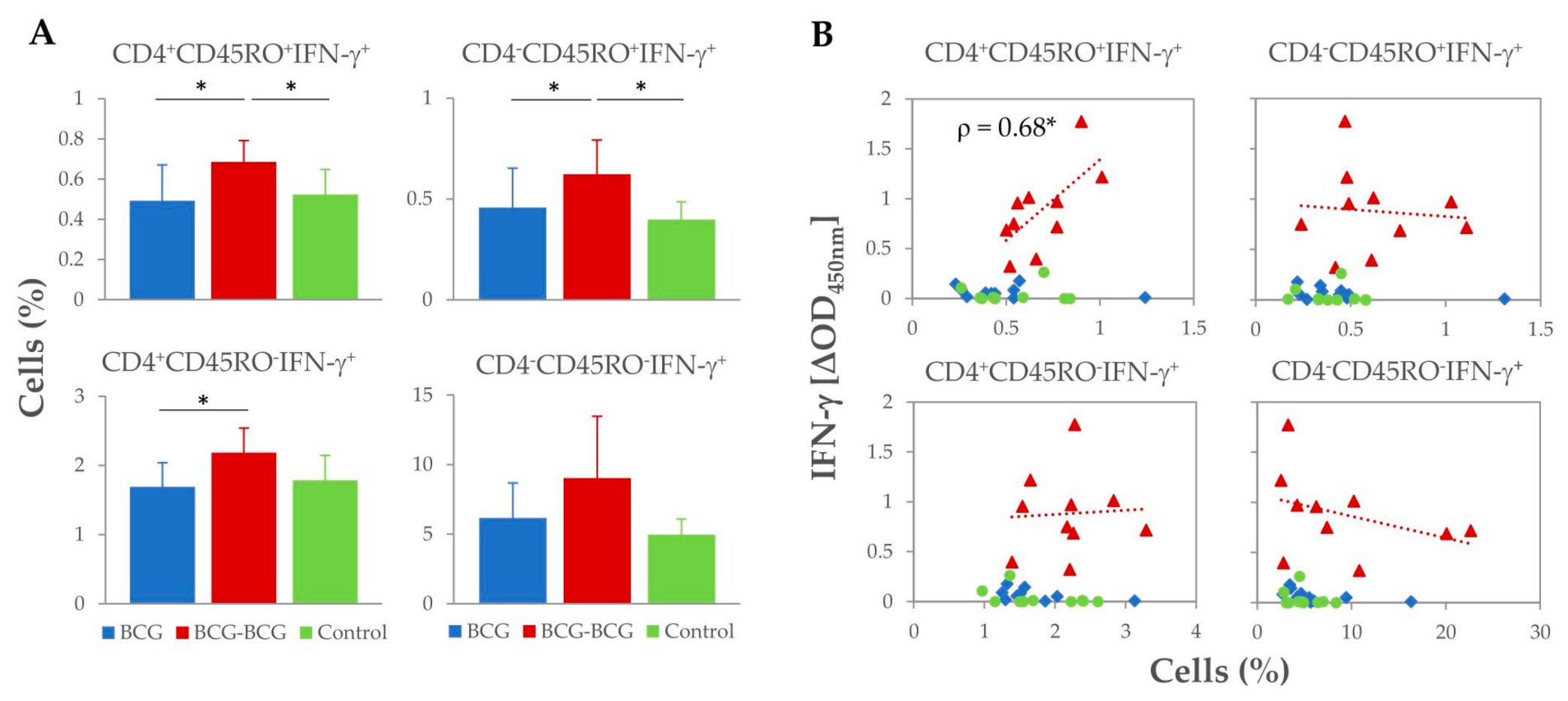
3.1. Immunological Responses after Vaccination and Revaccination
3.2. IFN-γ Responses After M. caprae Challenge
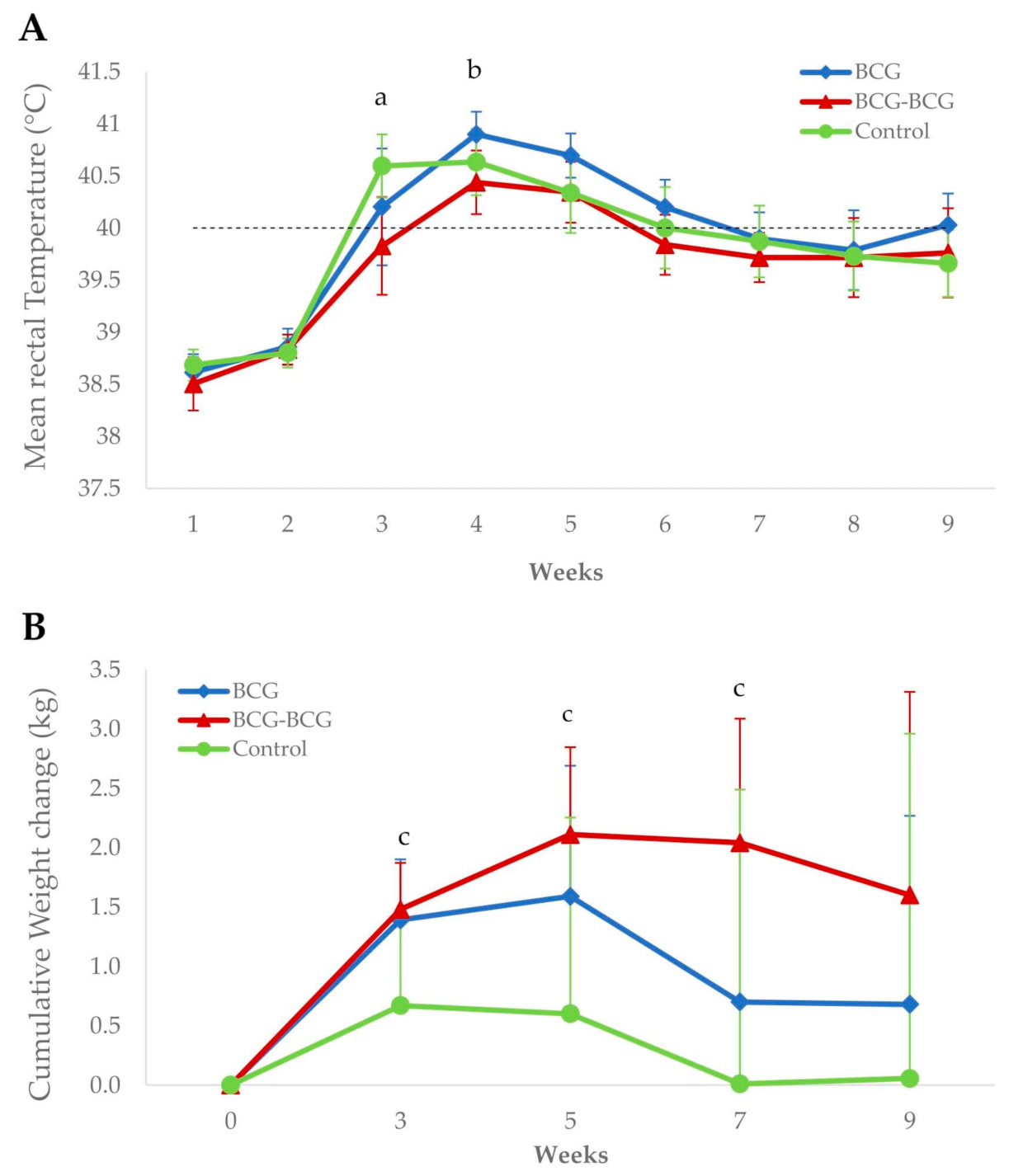
3.3. Clinical Signs and Body Condition Post-Challenge
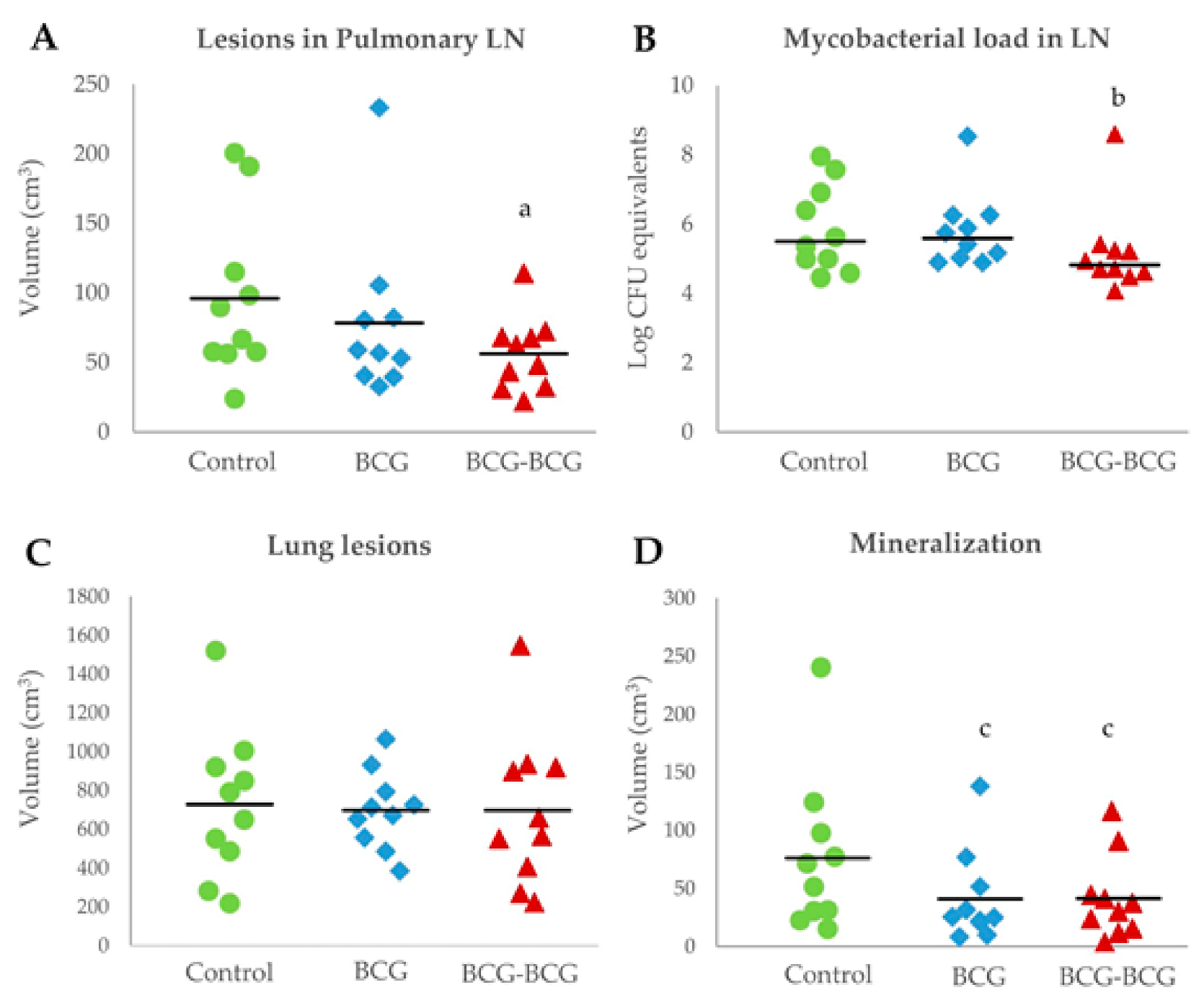
3.4. Post-Mortem Findings
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report 2019; World Health Organization: Geneva, Switzerland, 2019; Licence: CC BY-NC-SA 3.0 IGO; ISBN 9789241565714. [Google Scholar]
- Olea-Popelka, F.; Muwonge, A.; Perera, A.; Dean, A.S.; Mumford, E.; Erlacher-Vindel, E.; Forcella, S.; Silk, B.J.; Ditiu, L.; El Idrissi, A.; et al. Zoonotic tuberculosis in human beings caused by Mycobacterium bovis—A call for action. Lancet Infect. Dis. 2017, 17, e21–e25. [Google Scholar] [CrossRef] [Green Version]
- Arrieta-Villegas, C.; Perálvarez, T.; Vidal, E.; Puighibet, Z.; Moll, X.; Canturri, A.; Sevilla, I.A.; Espada, Y.; Juste, R.A.; Domingo, M.; et al. Efficacy of parenteral vaccination against tuberculosis with heat-inactivated Mycobacterium bovis in experimentally challenged goats. PLoS ONE 2018, 13, e0196948. [Google Scholar] [CrossRef] [PubMed]
- Cano-Terriza, D.; Risalde, M.A.; Rodríguez-Hernández, P.; Napp, S.; Fernández-Morente, M.; Moreno, I.; Bezos, J.; Fernández-Molera, V.; Sáez, J.L.; García-Bocanegra, I. Epidemiological surveillance of Mycobacterium tuberculosis complex in extensively raised pigs in the south of Spain. Prev. Vet. Med. 2018, 159, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Buddle, B.M.; Vordermeier, H.M.; Chambers, M.A.; de Klerk-Lorist, L.M. Efficacy and safety of BCG vaccine for control of tuberculosis in domestic livestock and wildlife. Front. Vet. Sci. 2018, 5, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, W.R.; Palmer, M.V.; Buddle, B.M.; Vordermeier, H.M. Bovine tuberculosis vaccine research: Historical perspectives and recent advances. Vaccine 2012, 30, 2611–2622. [Google Scholar] [CrossRef]
- Pérez De Val, B.; Vidal, E.; Villarreal-Ramos, B.; Gilbert, S.C.; Andaluz, A.; Moll, X.; Martín, M.; Nofrarías, M.; McShane, H.; Vordermeier, H.M.; et al. A multi-antigenic adenoviral-vectored vaccine improves BCG-induced protection of goats against pulmonary tuberculosis infection and prevents disease progression. PLoS ONE 2013, 8, e81317. [Google Scholar] [CrossRef]
- Roy, A.; Tomé, I.; Romero, B.; Lorente-Leal, V.; Infantes-Lorenzo, J.A.; Domínguez, M.; Martín, C.; Aguiló, N.; Puentes, E.; Rodríguez, E.; et al. Evaluation of the immunogenicity and efficacy of BCG and MTBVAC vaccines using a natural transmission model of tuberculosis. Vet. Res. 2019, 50, 82. [Google Scholar] [CrossRef] [Green Version]
- Kipnis, A.; Irwin, S.; Izzo, A.A.; Basaraba, R.J.; Orme, I.M. Memory T Lymphocytes Generated by Mycobacterium bovis BCG Vaccination Reside within a CD4 CD44lo CD62 Ligandhi Population. Infect. Immun. 2005, 73, 7759–7764. [Google Scholar] [CrossRef] [Green Version]
- Orme, I.M.; Henao-Tamayo, M.I. Trying to see the forest through the trees: Deciphering the nature of memory immunity to Mycobacterium tuberculosis. Front. Immunol. 2018, 9, 461. [Google Scholar] [CrossRef] [Green Version]
- Thom, M.L.; McAulay, M.; Vordermeier, H.M.; Clifford, D.; Hewinson, R.G.; Villarreal-Ramos, B.; Hope, J.C. Duration of Immunity against Mycobacterium bovis following Neonatal Vaccination with Bacillus Calmette-Guérin Danish: Significant Protection against Infection at 12, but Not 24, Months. Clin. Vaccine Immunol. 2012, 19, 1254–1260. [Google Scholar] [CrossRef] [Green Version]
- Parlane, N.A.; Shu, D.; Subharat, S.; Wedlock, D.N.; Rehm, B.H.A.; De Lisle, G.W.; Buddle, B.M. Revaccination of cattle with bacille Calmette-Guérin two years after first vaccination when immunity has waned, boosted protection against challenge with Mycobacterium bovis. PLoS ONE 2014, 9, e106519. [Google Scholar] [CrossRef] [Green Version]
- Vordermeier, H.M.; Jones, G.J.; Buddle, B.M.; Hewinson, R.G. Development of immune-diagnostic reagents to diagnose bovine tuberculosis in cattle. Vet. Immunol. Immunopathol. 2016, 181, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Balseiro, A.; Altuzarra, R.; Vidal, E.; Moll, X.; Espada, Y.; Sevilla, I.A.; Domingo, M.; Garrido, J.M.; Juste, R.A.; Prieto, M.; et al. Assessment of BCG and inactivated Mycobacterium bovis vaccines in an experimental tuberculosis infection model in sheep. PLoS ONE 2017, 12, e0180546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez de Val, B.; Vidal, E.; López-Soria, S.; Marco, A.; Cervera, Z.; Martín, M.; Mercader, I.; Singh, M.; Raeber, A.; Domingo, M. Assessment of safety and interferon gamma responses of Mycobacterium bovis BCG vaccine in goat kids and milking goats. Vaccine 2016, 34, 881–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez De Val, B.; López-Soria, S.; Nofrarías, M.; Martín, M.; Vordermeier, H.M.; Villarreal-Ramos, B.; Romera, N.; Escobar, M.; Solanes, D.; Cardona, P.J.; et al. Experimental model of tuberculosis in the domestic goat after endobronchial infection with Mycobacterium caprae. Clin. Vaccine Immunol. 2011, 18, 1872–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrieta-Villegas, C.; Allepuz, A.; Grasa, M.; Martín, M.; Cervera, Z.; Mercader, I.; López-Soria, S.; Domingo, M.; Pérez de Val, B. Long-term efficacy of BCG vaccination in goat herds with a high prevalence of tuberculosis. Sci. Rep. 2020, 10, 20369. [Google Scholar] [CrossRef] [PubMed]
- Vidal, E.; Arrieta-Villegas, C.; Grasa, M.; Mercader, I.; Domingo, M.; Pérez de Val, B. Field evaluation of the efficacy of Mycobacterium bovis BCG vaccine against tuberculosis in goats. BMC Vet. Res. 2017, 13, 252. [Google Scholar] [CrossRef] [Green Version]
- Pathak, S.; Awuh, J.A.; Leversen, N.A.; Flo, T.H.; Åsjø, B. Counting mycobacteria in infected human cells and mouse tissue: A comparison between qPCR and CFU. PLoS ONE 2012, 7, e34931. [Google Scholar] [CrossRef] [Green Version]
- Wangoo, A.; Johnson, L.; Gough, J.; Ackbar, R.; Inglut, S.; Hicks, D.; Spencer, Y.; Hewinson, G.; Vordermeier, M. Advanced granulomatous lesions in Mycobacterium bovis-infected cattle are associated with increased expression of type I procollagen, gammadelta (WC1+) T cells and CD 68+ cells. J. Comp. Pathol. 2005, 133, 223–234. [Google Scholar] [CrossRef]
- Buddle, B.M.; Wards, B.J.; Aldwell, F.E.; Collins, D.M.; De Lisle, G.W. Influence of sensitisation to environmental mycobacteria on subsequent vaccination against bovine tuberculosis. Vaccine 2002, 20, 1126–1133. [Google Scholar] [CrossRef]
- Andersen, P.; Doherty, T.M. The success and failure of BCG—Implications for a novel tuberculosis vaccine. Nat. Rev. Microbiol. 2005, 3, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Young, S.L.; Slobbe, L.; Wilson, R.; Buddle, B.M.; De Lisle, G.W.; Buchan, G.S. Environmental strains of Mycobacterium avium interfere with immune responses associated with Mycobacterium bovis BCG vaccination. Infect. Immun. 2007, 75, 2833–2840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whittaker, E.; Nicol, M.P.; Zar, H.J.; Tena-Coki, N.G.; Kampmann, B. Age-related waning of immune responses to BCG in healthy children supports the need for a booster dose of BCG in TB endemic countries. Sci. Rep. 2018, 8, 15309. [Google Scholar] [CrossRef]
- Buddle, B.M.; Wedlock, D.N.; Parlane, N.A.; Corner, L.A.L.; De Lisle, G.W.; Skinner, M.A. Revaccination of Neonatal Calves with Mycobacterium bovis BCG Reduces the Level of Protection against Bovine Tuberculosis Induced by a Single Vaccination. Infect. Immun. 2003, 71, 6411–6419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, P.; Smedegaard, B. CD4+ T-cell subsets that mediate immunological memory to Mycobacterium tuberculosis infection in mice. Infect. Immun. 2000, 68, 621–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sallusto, F.; Lenig, D.; Förster, R.; Lipp, M.; Lanzavecchia, A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 1999, 401, 708–712. [Google Scholar] [CrossRef] [PubMed]
- Maggioli, M.F.; Palmer, M.V.; Thacker, T.C.; Vordermeier, H.M.; Waters, W.R. Characterization of effector and memory T cell subsets in the immune response to bovine tuberculosis in cattle. PLoS ONE 2015, 10, e0122571. [Google Scholar] [CrossRef] [Green Version]
- Blunt, L.; Hogarth, P.J.; Kaveh, D.A.; Webb, P.; Villarreal-Ramos, B.; Vordermeier, H.M. Phenotypic characterization of bovine memory cells responding to mycobacteria in IFNγ enzyme linked immunospot assays. Vaccine 2015, 33, 7276–7282. [Google Scholar] [CrossRef] [Green Version]
- Weir, R.E.; Gorak-Stolinska, P.; Floyd, S.; Lalor, M.K.; Stenson, S.; Branson, K.; Blitz, R.; Ben-Smith, A.; Fine, P.E.M.; Dockrell, H.M. Persistence of the immune response induced by BCG vaccination. BMC Infect. Dis. 2008, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Uzal, F.A.; Bodero, D.A.V.; Kelly, W.R.; Nielsen, K. Variability of serum antibody responses of goat kids to a commercial Clostridium perfringens epsilon toxoid vaccine. Vet. Rec. 1998. [Google Scholar] [CrossRef]
- Tizard, I.R. Chapter 17—Sheep and Goat Vaccines. In Vaccines for Veterinarians; Elsevier: Amsterdam, The Netherlands, 2020; pp. 215–224.e1. [Google Scholar] [CrossRef]
- Lacasta, D.; Ferrer, L.M.; Ramos, J.J.; González, J.M.; Ortín, A.; Fthenakis, G.C. Vaccination schedules in small ruminant farms. Vet. Microbiol. 2015, 181, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Bezos, J.; Casal, C.; Puentes, E.; Díez-Guerrier, A.; Romero, B.; Aguiló, N.; de Juan, L.; Martín, C.; Domínguez, L. Evaluation of the immunogenicity and diagnostic interference caused by M. tuberculosis SO2 vaccination against tuberculosis in goats. Res. Vet. Sci. 2015, 103, 73–79. [Google Scholar] [CrossRef]
- van der Heijden, E.M.D.L.V.; Chileshe, J.; Vernooij, J.C.M.; Gortazar, C.; Juste, R.A.; Sevilla, I.; Crafford, J.E.; Rutten, V.P.M.G.; Michel, A.L. Immune response profiles of calves following vaccination with live BCG and inactivated Mycobacterium bovis vaccine candidates. PLoS ONE 2017, 12, e0188448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, G.J.; Whelan, A.; Clifford, D.; Coad, M.; Vordermeier, H.M. Improved skin test for differential diagnosis of bovine tuberculosis by the addition of Rv3020c-derived peptides. Clin. Vaccine Immunol. 2012, 19, 620–622. [Google Scholar] [CrossRef] [PubMed]
- Millington, K.A.; Fortune, S.M.; Low, J.; Garces, A.; Hingley-Wilson, S.M.; Wickremasinghe, M.; Kon, O.M.; Lalvani, A. Rv3615c is a highly immunodominant RD1 (Region of difference 1)-dependent secreted antigen specific for Mycobacterium tuberculosis infection. Proc. Natl. Acad. Sci. USA 2011, 108, 5730–5735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, S.; Jones, G.; Veerasami, M.; Steinbach, S.; Holder, T.; Zewude, A.; Fromsa, A.; Ameni, G.; Easterling, L.; Bakker, D.; et al. A defined antigen skin test for the diagnosis of bovine tuberculosis. Sci. Adv. 2019, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Post-Mortem Parameter | Body Weight Change | Rectal Temperature | |||||
|---|---|---|---|---|---|---|---|
| W 3 | W 5 | W 7 | W 9 | W 3 | W 4 | W 5 | |
| M. caprae CFU equivalents | −0.061 | −0.221 | −0.234 | −0.3035 | 0.234 | 0.27 | 0.336 |
| Vol Lesions in LN | −0.414 * | −0.511 ** | −0.576 *** | −0.542 ** | 0.444 * | 0.428 * | 0.435 * |
| Vol Lung Lesions | −0.319 | −0.482 ** | −0.587 *** | −0.692 *** | 0.277 | 0.456 * | 0.548 ** |
| Animals with Extrapulmonary Lesions | Localization of Lesions | |||||
|---|---|---|---|---|---|---|
| Spleen | Liver | Ln Ms 1 | Kidney | Ln RF 2 | ||
| BCG | 7/10 | 5 | 2 | 4 | 2 | 0 |
| BCG-BCG | 7/10 | 6 | 2 | 4 | 1 | 4 |
| Control | 9/10 | 8 | 4 | 8 | 4 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arrieta-Villegas, C.; Vidal, E.; Martín, M.; Verdés, J.; Moll, X.; Espada, Y.; Singh, M.; Villarreal-Ramos, B.; Domingo, M.; Pérez de Val, B. Immunogenicity and Protection against Mycobacterium caprae Challenge in Goats Vaccinated with BCG and Revaccinated after One Year. Vaccines 2020, 8, 751. https://doi.org/10.3390/vaccines8040751
Arrieta-Villegas C, Vidal E, Martín M, Verdés J, Moll X, Espada Y, Singh M, Villarreal-Ramos B, Domingo M, Pérez de Val B. Immunogenicity and Protection against Mycobacterium caprae Challenge in Goats Vaccinated with BCG and Revaccinated after One Year. Vaccines. 2020; 8(4):751. https://doi.org/10.3390/vaccines8040751
Chicago/Turabian StyleArrieta-Villegas, Claudia, Enric Vidal, Maite Martín, Judit Verdés, Xavier Moll, Yvonne Espada, Mahavir Singh, Bernardo Villarreal-Ramos, Mariano Domingo, and Bernat Pérez de Val. 2020. "Immunogenicity and Protection against Mycobacterium caprae Challenge in Goats Vaccinated with BCG and Revaccinated after One Year" Vaccines 8, no. 4: 751. https://doi.org/10.3390/vaccines8040751
APA StyleArrieta-Villegas, C., Vidal, E., Martín, M., Verdés, J., Moll, X., Espada, Y., Singh, M., Villarreal-Ramos, B., Domingo, M., & Pérez de Val, B. (2020). Immunogenicity and Protection against Mycobacterium caprae Challenge in Goats Vaccinated with BCG and Revaccinated after One Year. Vaccines, 8(4), 751. https://doi.org/10.3390/vaccines8040751