A Review: Aedes-Borne Arboviral Infections, Controls and Wolbachia-Based Strategies

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Aedes-Borne Arboviruses
3. Control Strategies for Aedes-Borne Viral Infections
4. Vector Control Methods for Aedes-Borne Viral Infections
4.1. Established Vector Control Methods
4.2. Experimental Vector Control Methods
5. Wolbachia Control Strategy
5.1. Wolbachia-Based Disruption of Arboviral Transmission
5.2. Wolbachia-Based Vector Population Suppression
6. Wolbachia-Based Field and Experimental Studies
7. Previous Studies on Mathematical Models of Wolbachia
8. Literature Gap, Future Research, and Conclusion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanley, K.A. Origin and Evolution of Viruses; Elsevier: Amsterdam, The Netherlands, 1998; pp. 351–391. [Google Scholar]
- Kamtchum-Tatuene, J.; Makepeace, B.L.; Benjamin, L.; Baylis, M.; Solomon, T. The potential role of Wolbachia in controlling the transmission of emerging human arboviral infections. Curr. Opin. Infect. Dis. 2017, 30, 108–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costard, S.; Mur, L.; Lubroth, J.; Sanchez-Vizcaino, J.; Pfeiffer, D. Epidemiology of African swine fever virus. Virus Res. 2013, 173, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Wasay, M.; Khatri, I.A.; Abd-Allah, F. Arbovirus infections of the nervous system: Current trends and future threats. Neurology 2015, 84, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.G.; Harris, E. Dengue. Lancet 2015, 385, 453–465. [Google Scholar] [CrossRef]
- Simmons, C.P.; Farrar, J.J.; Chau, N.V.V.; Wills, B. Dengue. N. Engl. J. Med. 2012, 366, 1423–1432. [Google Scholar] [CrossRef]
- Charlier, C.; Beaudoin, M.-C.; Couderc, T.; Lortholary, O.; Lecuit, M. Arboviruses and pregnancy: Maternal, fetal, and neonatal effects. Lancet Child. Adolesc. Heal. 2017, 1, 134–146. [Google Scholar] [CrossRef]
- Bergman, A.; Dahl, E.; Lundkvist, Å.; Hesson, J.C. Sindbis Virus Infection in Non-Blood-Fed Hibernating Culex pipiens Mosquitoes in Sweden. Viruses 2020, 12, 1441. [Google Scholar] [CrossRef]
- Snyder, R.E.; Feiszli, T.; Foss, L.; Messenger, S.; Fang, Y.; Barker, C.M.; Reisen, W.K.; Vugia, D.J.; Padgett, K.A.; Kramer, V.L. West Nile virus in California, 2003–2018: A persistent threat. PLoS Negl. Trop. Dis. 2020, 14, e0008841. [Google Scholar] [CrossRef]
- Chiuya, T.; Masiga, D.K.; Falzon, L.C.; Bastos, A.D.; Fèvre, E.M.; Villinger, J. Tick-Borne pathogens, including Crimean-Congo haemorrhagic fever virus, at livestock markets and slaughterhouses in western Kenya. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Deviatkin, A.A.; Kholodilov, I.S.; Vakulenko, Y.A.; Karganova, G.G.; Lukashev, A.N. Tick-Borne Encephalitis Virus: An Emerging Ancient Zoonosis? Viruses 2020, 12, 247. [Google Scholar] [CrossRef] [Green Version]
- Ayhan, N.; Charrel, R.N. An update on Toscana virus distribution, genetics, medical and diagnostic aspects. Clin. Microbiol. Infect. 2020, 26, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Ciota, A.T.; Kramer, L.D. Insights into Arbovirus Evolution and Adaptation from Experimental Studies. Viruses 2010, 2, 2594–2617. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.; Pettersson, J.; Higgs, S.; Charrel, R.; Lamballerie, X.d. Emerging arboviruses: Why today? One Health 2017, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mavian, C.; Dulcey, M.; Munoz, O.; Salemi, M.; Vittor, A.Y.; Capua, I. Islands as Hotspots for Emerging Mosquito-Borne Viruses: A One-Health Perspective. Viruses 2018, 11, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nat. Cell Biol. 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.P.; Luo, Y.; Chen, Y.; Lamers, M.M.; Zhou, Q.; Yang, X.H.; Sanyal, S.; Mok, C.K.P.; Liu, Z.M. Clinical and epidemiological features of the 2014 large-Scale dengue outbreak in Guangzhou city, China. BMC Infect. Dis. 2016, 16, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messina, J.P.; Brady, O.J.; Golding, N.; Kraemer, M.U.G.; Wint, G.R.W.; Ray, S.E.; Pigott, D.M.; Shearer, F.M.; Johnson, K.; Earl, L.; et al. The current and future global distribution and population at risk of dengue. Nat. Microbiol. 2019, 4, 1508–1515. [Google Scholar] [CrossRef]
- Xu, Z.; Bambrick, H.; Frentiu, F.D.; Devine, G.; Yakob, L.; Williams, G.; Hu, W. Projecting the future of dengue under climate change scenarios: Progress, uncertainties and research needs. PLoS Negl. Trop. Dis. 2020, 14, e0008118. [Google Scholar] [CrossRef]
- Beckham, J.D.; Tyler, K.L. Arbovirus Infections. Continu. (Minneap. Minn) 2015, 21, 1599–1611. [Google Scholar] [CrossRef] [Green Version]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [Green Version]
- Rust, R.S. Human Arboviral Encephalitis. Semin. Pediatr. Neurol. 2012, 19, 130–151. [Google Scholar] [CrossRef] [PubMed]
- Wieten, R.W.; Goorhuis, A.; Jonker, E.F.F.; De Bree, G.J.; Visser, A.W.d.; Van Genderen, P.J.J.; Remmerswaal, E.B.M.; Berge, I.J.M.T.; Visser, L.G.; Grobusch, M.P.; et al. 17D yellow fever vaccine elicits comparable long-Term immune responses in healthy individuals and immune-Compromised patients. J. Infect. 2016, 72, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-L.; Chang, J.-K.; Tang, R.-B. Current recommendations for the Japanese encephalitis vaccine. J. Chin. Med. Assoc. 2015, 78, 271–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, L.J. Tetravalent Dengue Vaccine: A Review in the Prevention of Dengue Disease. Drugs 2016, 76, 1301–1312. [Google Scholar] [CrossRef]
- Marston, H.D.; Lurie, N.; Borio, L.L.; Fauci, A.S. Considerations for Developing a Zika Virus Vaccine. N. Engl. J. Med. 2016, 375, 1209–1212. [Google Scholar] [CrossRef]
- Brandler, S.; Tangy, F. Vaccines in Development against West Nile Virus. Viruses 2013, 5, 2384–2409. [Google Scholar] [CrossRef] [Green Version]
- Wressnigg, N.; Van Der Velden, M.V.W.; Portsmouth, D.; Draxler, W.; O’Rourke, M.; Richmond, P.; Hall, S.; McBride, W.J.H.; Redfern, A.; Aaskov, J.; et al. An Inactivated Ross River Virus Vaccine Is Well Tolerated and Immunogenic in an Adult Population in a Randomized Phase 3 Trial. Clin. Vaccine Immunol. 2015, 22, 267–273. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.; Diego, L.; Judith, B. New Approaches to Chikungunya Virus Vaccine Development. Recent Pat. Inflamm. Allergy Drug Discov. 2015, 9, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Buhler, C.; Winkler, V.; Runge-Ranzinger, S.; Boyce, R.; Horstick, O. Environmental methods for dengue vector control–A systematic review and meta-Analysis. PLoS Negl. Trop. Dis. 2019, 13, e0007420. [Google Scholar] [CrossRef]
- Blagrove, M.S.C.; Arias-Goeta, C.; Failloux, A.-B.; Sinkins, S.P. Wolbachia strain wMel induces cytoplasmic incompatibility and blocks dengue transmission in Aedes albopictus. Proc. Natl. Acad. Sci. USA 2011, 109, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Crain, P.R.; Mains, J.W.; Suh, E.; Huang, Y.; Crowley, P.H.; Dobson, S.L. Wolbachia infections that reduce immature insect survival: Predicted impacts on population replacement. BMC Evol. Biol. 2011, 11, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joubert, D.A.; Walker, T.; Carrington, L.B.; De Bruyne, J.T.; Kien, D.H.T.; Hoang, N.L.T.; Chau, N.V.V.; Iturbe-Ormaetxe, I.; Simmons, C.P.; O’Neill, S.L. Establishment of a Wolbachia Superinfection in Aedes aegypti Mosquitoes as a Potential Approach for Future Resistance Management. PLoS Pathog. 2016, 12, e1005434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, G.A.; Hoffmann, A.A.; Maciel-De-Freitas, R.; Villela, D.A.M. Aedes aegypti insecticide resistance underlies the success (and failure) of Wolbachia population replacement. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nazni, W.A.; Hoffmann, A.A.; NoorAfizah, A.; Cheong, Y.L.; Mancini, M.V.; Golding, N.; Kamarul, G.M.R.; Arif, M.A.K.; Thohir, H.; NurSyamimi, H.; et al. Establishment of Wolbachia Strain wAlbB in Malaysian Populations of Aedes aegypti for Dengue Control. Curr. Biol. 2019, 29, 4241–4248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Reilly, K.M.; Hendrickx, E.; Kharisma, D.D.; Wilastonegoro, N.N.; Carrington, L.B.; Elyazar, I.R.F.; Kucharski, A.J.; Lowe, R.; Flasche, S.; Pigott, D.M.; et al. Estimating the burden of dengue and the impact of release of wMel Wolbachia-Infected mosquitoes in Indonesia: A modelling study. BMC Med. 2019, 17, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, R.; Lee, S.; Lana, R.M.; Codeço, C.T.; Castro, M.C.; Pascual, M. Emerging arboviruses in the urbanized Amazon rainforest. BMJ 2020, 371, m4385. [Google Scholar] [CrossRef]
- Madewell, Z.J. Arboviruses and Their Vectors. South Med. J. 2020, 113, 520–523. [Google Scholar] [CrossRef]
- Causa, R.; Ochoa-Diaz-Lopez, H.; Dor, A.; Rodríguez-León, F.; Solís-Hernández, R.; Pacheco-Soriano, A.L. Emerging arboviruses (dengue, chikungunya, and Zika) in Southeastern Mexico: Influence of socio-Environmental determinants on knowledge and practices. Cad. Saúde Pública 2020, 36, e00110519. [Google Scholar] [CrossRef]
- Silva, P.A.N.d.; Ito, C.R.M.; Barbosa, M.S.; Santos, M.d.O.; Carneiro, L.C. Arboviruses (chikungunya, dengue, and Zika) associated with ophthalmic changes: A focus on aqueous fluid and vitreous humor. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 39, 827–833. [Google Scholar] [CrossRef]
- Kularatne, S. Dengue fever. BMJ 2015, 351, h4661. [Google Scholar] [CrossRef]
- Chan, M.; Johansson, M.A. The Incubation Periods of Dengue Viruses. PLoS ONE 2012, 7, e50972. [Google Scholar] [CrossRef] [PubMed]
- Brady, O.J.; Gething, P.W.; Bhatt, S.; Messina, J.P.; Brownstein, J.S.; Hoen, A.G.; Moyes, C.L.; Farlow, A.W.; Scott, T.W.; Hay, S.I. Refining the Global Spatial Limits of Dengue Virus Transmission by Evidence-Based Consensus. PLoS Negl. Trop. Dis. 2012, 6, e1760. [Google Scholar] [CrossRef]
- Dick, G.W.A.; Kitchen, S.F.; Haddow, A.J. Zika Virus (I). Isolations and serological specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Bogoch, I.I.; Brady, O.J.; Kraemer, M.U.G.; German, M.J.; Creatore, M.I.; Kulkarni, M.A.; Brownstein, J.S.; Mekaru, S.R.; Hay, S.I.; Groot, E.; et al. Anticipating the international spread of Zika virus from Brazil. Lancet 2016, 387, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Guerbois, M.; Fernandez-Salas, I.; Azar, S.R.; Danis-Lozano, R.; Alpuche-Aranda, C.M.; Leal, G.; Garcia-Malo, I.R.; Diaz-Gonzalez, E.E.; Casas-Martinez, M.; Rossi, S.L.; et al. Outbreak of Zika Virus Infection, Chiapas State, Mexico, 2015, and First Confirmed Transmission byAedes aegyptiMosquitoes in the Americas. J. Infect. Dis. 2016, 214, 1349–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamel, R.; Liégeois, F.; Wichit, S.; Pompon, J.; Diop, F.; Talignani, L.; Thomas, F.; Després, P.; Yssel, H.; Missé, D. Zika virus: Epidemiology, clinical features and host-Virus interactions. Microbes Infect. 2016, 18, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Musso, D.; Nhan, T.; Robin, E.; Roche, C.; Bierlaire, D.; Zisou, K.; Yan, A.S.; Cao-Lormeau, V.M.; Broult, J. Potential for Zika virus transmission through blood transfusion demonstrated during an outbreak in French Polynesia, November 2013 to February 2014. Eurosurveillance 2014, 19, 20761. [Google Scholar] [CrossRef] [Green Version]
- Brooks, J.T.; Friedman, A.; Kachur, R.E.; LaFlam, M.; Peters, P.J.; Jamieson, D.J. Update: Interim Guidance for Prevention of Sexual Transmission of Zika Virus-United States. Morb. Mortal. Wkly. Rep. 2016, 65, 745–747. [Google Scholar] [CrossRef] [Green Version]
- Motta, I.J.; Spencer, B.R.; Silva, S.G.C.d.; Arruda, M.B.; Dobbin, J.A.; Gonzaga, Y.B.; Arcuri, I.P.; Tavares, R.C.; Atta, E.H.; Fernandes, R.F.; et al. Evidence for Transmission of Zika Virus by Platelet Transfusion. N. Engl. J. Med. 2016, 375, 1101–1103. [Google Scholar] [CrossRef]
- Brady, O.J.; Osgood-Zimmerman, A.; Kassebaum, N.J.; Ray, S.E.; De Araújo, V.E.M.; Da Nóbrega, A.A.; Frutuoso, L.C.V.; Lecca, R.C.R.; Stevens, A.; De Oliveira, B.Z.; et al. The association between Zika virus infection and microcephaly in Brazil 2015–2017: An observational analysis of over 4 million births. PLoS Med. 2019, 16, e1002755. [Google Scholar] [CrossRef]
- WHO. Fact Sheets: Chikungunya; World Health Organization, 2017; Available online: https://www.who.int/news-room/fact-sheets/detail/chikungunya (accessed on 15 June 2020).
- Parola, P.; De Lamballerie, X.; Jourdan, J.; Rovery, C.; Vaillant, V.; Minodier, P.; Brouqui, P.; Flahault, A.; Raoult, D.; Charrel, R.N. Novel Chikungunya Virus Variant in Travelers Returning from Indian Ocean Islands. Emerg. Infect. Dis. 2006, 12, 1493–1499. [Google Scholar] [CrossRef] [PubMed]
- Couderc, T.; Gangneux, N.; Caro, V.; Ducloux, B.; Tolou, H.; Chrétien, F.; Luong, T.L.; Lecuit, M.; Grandadam, M. Chikungunya Virus Infection of Corneal Grafts. J. Infect. Dis. 2012, 206, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Lenglet, Y.; Barau, G.; Robillard, P.Y.; Randrianaivo, H.; Michault, A.; Bouveret, A.; Gerardin, P.; Boumahni, B.; Touret, Y.; Kauffmann, E.; et al. Chikungunya infection in pregnancy: Evidence for intrauterine infection in pregnant women and vertical transmission in the parturient. Survey of the Reunion Island outbreak. J. Gynecol. Obstet. Biol. Rep. (Paris) 2006, 35, 578–583. [Google Scholar] [CrossRef]
- Mutebi, J.-P.; Wang, H.; Li, L.; Bryant, J.E.; Barrett, A.D.T. Phylogenetic and Evolutionary Relationships among Yellow Fever Virus Isolates in Africa. J. Virol. 2001, 75, 6999–7008. [Google Scholar] [CrossRef] [Green Version]
- Mathai, D.; Vasanthan, A.G. State of the globe: Yellow fever is still around and active! J. Glob. Infect. Dis. 2009, 1, 4–6. [Google Scholar] [CrossRef]
- Rogers, D.; Wilson, A.J.; Hay, S.I.; Graham, A. The Global Distribution of Yellow Fever and Dengue. Adv. Parasitol. 2006, 62, 181–220. [Google Scholar] [CrossRef] [Green Version]
- Bae, H.-G.; Drosten, C.; Emmerich, P.; Colebunders, R.; Hantson, P.; Pest, S.; Parent, M.; Schmitz, H.; Warnat, M.-A.; Niedrig, M. Analysis of two imported cases of yellow fever infection from Ivory Coast and The Gambia to Germany and Belgium. J. Clin. Virol. 2005, 33, 274–280. [Google Scholar] [CrossRef]
- Xiao, S.-Y.; Zhang, H.; Guzman, H.; Tesh, R.B. Experimental Yellow Fever Virus Infection in the Golden Hamster (Mesocricetus auratus). II. Pathology. J. Infect. Dis. 2001, 183, 1437–1444. [Google Scholar] [CrossRef] [Green Version]
- Tesh, R.; Guzman, H.; Rosa, A.P.A.T.d.; Vasconcelos, P.F.C.; Dias, L.B.; Bunnell, J.E.; Zhang, H.; Xiao, S.-Y. Experimental Yellow Fever Virus Infection in the Golden Hamster (Mesocricetus auratus). I. Virologic, Biochemical, and Immunologic Studies. J. Infect. Dis. 2001, 183, 1431–1436. [Google Scholar] [CrossRef] [Green Version]
- Barba-Spaeth, G.; Dejnirattisai, W.; Rouvinski, A.; Vaney, M.-C.; Medits, I.; Sharma, A.; Simon-Lorière, E.; Sakuntabhai, A.; Cao-Lormeau, V.-M.; Haouz, A.; et al. Structural basis of potent Zika–Dengue virus antibody cross-Neutralization. Nat. Cell Biol. 2016, 536, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, R. Antibody-Dependent Enhancement of Viral Infections. In Dynamics of Immune Activation in Viral Diseases; Springer: Singapore, 2020; pp. 9–41. [Google Scholar] [CrossRef] [Green Version]
- Smatti, M.K.; Al Thani, A.A.; Yassine, H.M. Viral-Induced Enhanced Disease Illness. Front. Microbiol. 2018, 9, 2991. [Google Scholar] [CrossRef] [Green Version]
- Fukusumi, M.; Arashiro, T.; Arima, Y.; Matsui, T.; Shimada, T.; Kinoshita, H.; Arashiro, A.; Takasaki, T.; Sunagawa, T.; Oishi, K. Dengue Sentinel Traveler Surveillance: Monthly and Yearly Notification Trends among Japanese Travelers, 2006–2014. PLoS Negl. Trop. Dis. 2016, 10, e0004924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguiar, M.; Stollenwerk, N.; Halstead, S.B. The Impact of the Newly Licensed Dengue Vaccine in Endemic Countries. PLoS Negl. Trop. Dis. 2016, 10, e0005179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priyamvada, L.; Hudson, W.; Ahmed, R.; Wrammert, J. Humoral cross-Reactivity between Zika and dengue viruses: Implications for protection and pathology. Emerg. Microbes Infect. 2017, 6, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudolph, K.E.; Moloney, R.M.; Kmush, B.; Cummings, D.A.T.; Lessler, J. Incubation Periods of Mosquito-Borne Viral Infections: A Systematic Review. Am. J. Trop. Med. Hyg. 2014, 90, 882–891. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, T.J.; Panda, P.K.; Wolford, R.W. Dengue Fever; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Goh, G.K.-M.; Dunker, A.K.; Foster, J.A.; Uversky, V.N. Zika and Flavivirus Shell Disorder: Virulence and Fetal Morbidity. Biomolecules 2019, 9, 710. [Google Scholar] [CrossRef] [Green Version]
- Brasil, P.; Calvet, G.A.; Siqueira, A.M.; Wakimoto, M.; Sequeira, P.C.d.; Nobre, A.; Quintana, M.D.S.B.; Mendonça, M.C.L.d.; Lupi, O.; Souza, R.V.d.; et al. Zika Virus Outbreak in Rio de Janeiro, Brazil: Clinical Characterization, Epidemiological and Virological Aspects. PLoS Negl. Trop. Dis. 2016, 10, e0004636. [Google Scholar] [CrossRef]
- Chen, L.H. Zika Virus Infection in a Massachusetts Resident After Travel to Costa Rica: A Case Report. Ann. Intern. Med. 2016, 164, 574–576. [Google Scholar] [CrossRef] [Green Version]
- CDC. Centers for Disease Control and Prevention: Zika Virus, Symptoms Testing & Treatment. 2019. Available online: https://www.cdc.gov/zika/symptoms/treatment.html (accessed on 21 October 2019).
- Waggoner, J.J.; Gresh, L.; Vargas, M.J.; Ballesteros, G.; Tellez, Y.; Soda, K.J.; Sahoo, M.K.; Nuñez, A.; Balmaseda, A.; Harris, E.; et al. Viremia and Clinical Presentation in Nicaraguan Patients Infected with Zika Virus, Chikungunya Virus, and Dengue Virus. Clin. Infect. Dis. 2016, 63, 1584–1590. [Google Scholar] [CrossRef]
- Fourié, T.; Grard, G.; Leparc-Goffart, I.; Briolant, S.; Fontaine, A. Variability of Zika Virus Incubation Period in Humans. Open Forum Infect. Dis. 2018, 5, 261. [Google Scholar] [CrossRef]
- Krow-Lucal, E.R.; Biggerstaff, B.J.; Staples, J.E. Estimated Incubation Period for Zika Virus Disease. Emerg. Infect. Dis. 2017, 23, 841–845. [Google Scholar] [CrossRef] [PubMed]
- Winokur, O.C.; Main, B.J.; Nicholson, J.; Barker, C.M. Impact of temperature on the extrinsic incubation period of Zika virus in Aedes aegypti. PLoS Negl. Trop. Dis. 2020, 14, e0008047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffy, M.R.; Chen, T.-H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; DuBray, C.; et al. Zika Virus Outbreak on Yap Island, Federated States of Micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, V.K.; Duan, B.; Reid, S.P. Chikungunya Virus: Pathophysiology, Mechanism, and Modeling. Viruses 2017, 9, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, S.C.; Lecuit, M. Chikungunya Virus and the Global Spread of a Mosquito-Borne Disease. N. Engl. J. Med. 2015, 372, 1231–1239. [Google Scholar] [CrossRef] [Green Version]
- Javelle, E.; Tiong, T.H.; Leparc-Goffart, I.; Savini, H.; Simon, F. Inflammation of the external ear in acute chikungunya infection: Experience from the outbreak in Johor Bahru, Malaysia, 2008. J. Clin. Virol. 2014, 59, 270–273. [Google Scholar] [CrossRef]
- Miner, J.J.; Yeang, H.X.A.; Fox, J.M.; Taffner, S.; Malkova, O.N.; Oh, S.T.; Kim, A.H.J.; Diamond, M.S.; Lenschow, D.J.; Yokoyama, W.M. Brief Report: Chikungunya Viral Arthritis in the United States: A Mimic of Seronegative Rheumatoid Arthritis. Arthritis Rheumatol. 2015, 67, 1214–1220. [Google Scholar] [CrossRef] [Green Version]
- Nayar, S.K.; Noridah, O.; Paranthaman, V.; Ranjit, K.; Norizah, I.; Chem, Y.K.; Mustafa, B.; Chua, K.B. Co-Infection of dengue virus and chikungunya virus in two patients with acute febrile illness. Med. J. Malays. 2007, 62, 335–336. [Google Scholar]
- Dubrulle, M.; Mousson, L.; Moutailler, S.; Vazeille, M.; Failloux, A.-B. Chikungunya Virus and Aedes Mosquitoes: Saliva Is Infectious as soon as Two Days after Oral Infection. PLoS ONE 2009, 4, e5895. [Google Scholar] [CrossRef]
- Nakkhara, P.; Chongsuvivatwong, V.; Thammapalo, S. Risk factors for symptomatic and asymptomatic chikungunya infection. Trans. R. Soc. Trop. Med. Hyg. 2013, 107, 789–796. [Google Scholar] [CrossRef]
- Kamgang, B.; Vazeille, M.; Yougang, A.P.; Tedjou, A.N.; Wilson-Bahun, T.A.; Mousson, L.; Wondji, C.S.; Failloux, A.-B. Potential of Aedes albopictus and Aedes aegypti (Diptera: Culicidae) to transmit yellow fever virus in urban areas in Central Africa. Emerg. Microbes Infect. 2019, 8, 1636–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnett, E.D. Yellow Fever: Epidemiology and Prevention. Clin. Infect. Dis. 2007, 44, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Klitting, R.; Gould, E.A.; Paupy, C.; Lamballerie, X.d. What Does the Future Hold for Yellow Fever Virus? (I). Genes 2018, 9, 291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo, A.B.d.; Silva, M.D.P.C.D.; Bertani, G.R.; Gil, L.H.V.G.; Magalhães, M.C.F.; Cordeiro, M.T.; Marques, E.T.A.; Carvalho, E.M.F.d.; Braga-Neto, U.M. Description of a Prospective 17DD Yellow Fever Vaccine Cohort in Recife, Brazil. Am. J. Trop. Med. Hyg. 2011, 85, 739–747. [Google Scholar] [CrossRef] [Green Version]
- Sbrana, E.; Xiao, S.Y.; Guzman, H.; Ye, M.; Rosa, A.P.T.d.; Tesh, R.B. Efficacy of post-Exposure treatment of yel-Low fever with ribavirin in a hamster model of the disease. Am. J. Trop Med. Hyg. 2004, 71, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Gould, L.H.; Osman, M.S.; Farnon, E.C.; Griffith, K.S.; Godsey, M.S.; Karch, S.; Mulenda, B.; Kholy, A.E.; Grandesso, F.; Radiguès, X.d.; et al. An outbreak of yellow fever with concurrent chikungunya virus transmission in South Kordofan, Sudan, 2005. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 1247–1254. [Google Scholar] [CrossRef]
- Johansson, M.A.; Biggerstaff, B.J.; Arana-Vizcarrondo, N.; Staples, J.E. Incubation Periods of Yellow Fever Virus. Am. J. Trop. Med. Hyg. 2010, 83, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.A.; Vasconcelos, P.F.C.; Staples, J.E. The whole iceberg: Estimating the incidence of yellow fever virus infection from the number of severe cases. Trans. R. Soc. Trop. Med. Hyg. 2014, 108, 482–487. [Google Scholar] [CrossRef] [Green Version]
- Roiz, D.; Wilson, A.L.; Scott, T.W.; Fonseca, D.M.; Jourdain, F.; Müller, P.; Velayudhan, R.; Corbel, V. Integrated Aedes management for the control of Aedes-borne diseases. PLoS Negl. Trop. Dis. 2018, 12, e0006845. [Google Scholar] [CrossRef] [Green Version]
- Rather, I.A.; Parray, H.A.; Lone, J.B.; Paek, W.K.; Lim, J.; Bajpai, V.K.; Park, Y.-H. Prevention and Control Strategies to Counter Dengue Virus Infection. Front. Cell. Infect. Microbiol. 2017, 7, 336. [Google Scholar] [CrossRef] [Green Version]
- Whitehorn, J.; Yacoub, S.; Anders, K.L.; Macareo, L.R.; Cassetti, M.C.; Van, V.C.N.; Shi, P.-Y.; Wills, B.; Simmons, C.P. Dengue Therapeutics, Chemoprophylaxis, and Allied Tools: State of the Art and Future Directions. PLoS Negl. Trop. Dis. 2014, 8, e3025. [Google Scholar] [CrossRef] [PubMed]
- Hladish, T.J.; Pearson, C.A.B.; Ben Toh, K.; Rojas, D.P.; Manrique-Saide, P.; Vazquez-Prokopec, G.M.; Halloran, M.E.; Longini, I.M. Designing effective control of dengue with combined interventions. Proc. Natl. Acad. Sci. USA 2020, 117, 3319–3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilder-Smith, A.; Flasche, S.; Smith, P.G. Vaccine-attributable severe dengue in the Philippines. Lancet 2019, 394, 2151–2152. [Google Scholar] [CrossRef] [Green Version]
- Heydari, N.; Larsen, D.A.; Neira, M.; Ayala, E.B.; Fernandez, P.; Adrian, J.; Rochford, R.; Stewart-Ibarra, A.M. Household Dengue Prevention Interventions, Expenditures, and Barriers to Aedes aegypti Control in Machala, Ecuador. Int. J. Environ. Res. Public Heal. 2017, 14, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.H.; Kozarsky, P.E.; Freedman, D.O. Medical Considerations before International Travel. N. Engl. J. Med. 2016, 375, e32. [Google Scholar] [CrossRef] [PubMed]
- Rapley, L.P.; Russell, R.C.; Montgomery, B.L.; Ritchie, S.A. The Effects of Sustained Release Metofluthrin on the Biting, Movement, and Mortality of Aedes aegypti in a Domestic Setting. Am. J. Trop. Med. Hyg. 2009, 81, 94–99. [Google Scholar] [CrossRef]
- Achee, N.L.; Gould, F.; Perkins, T.A.; Reiner, R.C., Jr.; Morrison, A.C.; Ritchie, S.A.; Gubler, D.J.; Teyssou, R.; Scott, T.W. A Critical Assessment of Vector Control for Dengue Prevention. PLoS Negl. Trop. Dis. 2015, 9, e0003655. [Google Scholar] [CrossRef]
- Hunter, P. Challenges and options for disease vector control: The outbreak of Zika virus in South America and increasing insecticide resistance among mosquitoes have rekindled efforts for controlling disease vectors. EMBO Rep. 2016, 17, 1370–1373. [Google Scholar] [CrossRef] [Green Version]
- Horstick, O.; Boyce, R.; Runge-Ranzinger, S. Dengue vector control: Assessing what works? Southeast Asian J. Trop Med. Public Health 2017, 48, 181–195. [Google Scholar]
- Flores, H.; Neill, S.L.O. Controlling vector-Borne diseases by releasing modified mosquitoes. Nat. Rev. Microbiol. 2018, 16, 508–518. [Google Scholar] [CrossRef]
- Yen, P.-S.; Failloux, A.-B. A Review: Wolbachia-Based Population Replacement for Mosquito Control Shares Common Points with Genetically Modified Control Approaches. Pathogens 2020, 9, 404. [Google Scholar] [CrossRef] [PubMed]
- Choi, L.; Majambere, S.; Wilson, A.L. Larviciding to prevent malaria transmission. Cochrane Database Syst. Rev. 2019, 8, CD012736. [Google Scholar] [CrossRef] [PubMed]
- Lima, E.P.; Paiva, M.H.; Araujo, A.P.d.; Silva, E.V.d.; Silva, U.M.d.; Oliveira, L.N.d.; Santana, A.E.; Barbosa, C.N.; Neto, C.C.d.P.; Goulart, M.O.; et al. Insecticide resistance in Aedes aegypti populations from Ceará, Brazil. Parasites Vectors 2011, 4, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dusfour, I.; Vontas, J.; David, J.-P.; Weetman, D.; Fonseca, D.M.; Corbel, V.; Raghavendra, K.; Coulibaly, M.B.; Martins, A.J.; Kasai, S.; et al. Management of insecticide resistance in the major Aedes vectors of arboviruses: Advances and challenges. PLoS Negl. Trop. Dis. 2019, 13, e0007615. [Google Scholar] [CrossRef] [PubMed]
- Karunamoorthi, K. Vector control: A cornerstone in the malaria elimination campaign. Clin. Microbiol. Infect. 2011, 17, 1608–1616. [Google Scholar] [CrossRef] [Green Version]
- Han, W.W.; Lazaro, A.; McCall, P.J.; George, L.; Runge-Ranzinger, S.; Toledo, J.; Velayudhan, R.; Horstick, O. Efficacy and community effectiveness of larvivorous fish for dengue vector control. Trop. Med. Int. Heal. 2015, 20, 1239–1256. [Google Scholar] [CrossRef]
- Udayanga, L.; Ranathunge, T.; Iqbal, M.C.M.; Abeyewickreme, W.; Hapugoda, M.D. Predatory efficacy of five locally available copepods on Aedes larvae under laboratory settings: An approach towards bio-Control of dengue in Sri Lanka. PLoS ONE 2019, 14, e0216140. [Google Scholar] [CrossRef] [Green Version]
- Tissera, H.A.; Samaraweera, P.C.; Jayamanne, B.D.W.; Janaki, M.D.S.; Chulasiri, M.P.P.U.; Rodrigo, C.; Fernando, S.D. Use of Bacillus thuringiensis israelensis in integrated vector control of Aedes sp. in Sri Lanka: A prospective controlled effectiveness study. Trop. Med. Int. Heal. 2017, 23, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Lazaro, A.; Han, W.W.; Manrique-Saide, P.; George, L.; Velayudhan, R.; Toledo, J.; Ranzinger, S.R.; Horstick, O. Community effectiveness of copepods for dengue vector control: Systematic review. Trop. Med. Int. Heal. 2015, 20, 685–706. [Google Scholar] [CrossRef]
- Alphey, L.; McKemey, A.; Nimmo, D.; Oviedo, M.N.; Lacroix, R.; Matzen, K.; Beech, C. Genetic control ofAedesmosquitoes. Pathog. Glob. Heal. 2013, 107, 170–179. [Google Scholar] [CrossRef] [Green Version]
- Atyame, C.M.; Cattel, J.; Lebon, C.; Flores, O.; Dehecq, J.-S.; Weill, M.; Gouagna, L.C.; Tortosa, P. Wolbachia-Based Population Control Strategy Targeting Culex quinquefasciatus Mosquitoes Proves Efficient under Semi-Field Conditions. PLoS ONE 2015, 10, e0119288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Staples, D.; Diaz-Fleischer, F.; Montoya, P. The Sterile Insect Technique: Success and Perspectives in the Neotropics. Néotrop. Èntomol. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Kittayapong, P.; Ninphanomchai, S.; Limohpasmanee, W.; Chansang, C.; Chansang, U.; Mongkalangoon, P. Combined sterile insect technique and incompatible insect technique: The first proof-Of-Concept to suppress Aedes aegypti vector populations in semi-Rural settings in Thailand. PLoS Negl. Trop. Dis. 2019, 13, e0007771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustafa, M.S.; Bansal, A.; Rastogi, V. Flightless Aedes mosquitoes in dengue control. Med. J. Armed Forces India 2011, 67, 192–193. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.-N.; Rodriguez, S.D.; Gonzales, K.K.; Vulcan, J.; Cordova, J.J.; Mitra, S.; Adams, C.G.; Moses-Gonzales, N.; Tam, N.; Cluck, J.W.; et al. Toward Implementation of Mosquito Sterile Insect Technique: The Effect of Storage Conditions on Survival of MaleAedes aegyptiMosquitoes (Diptera: Culicidae) During Transport. J. Insect Sci. 2018, 18. [Google Scholar] [CrossRef] [Green Version]
- Dickens, B.L.; Yang, J.; Cook, A.R.; Carrasco, L.R. Time to Empower Release of Insects Carrying a Dominant Lethal and Wolbachia Against Zika. Open Forum Infect. Dis. 2016, 3, ofw103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-Payá, D.; Flis, I.; Anderson, M.; Hawes, P.; Li, M.; Akbari, O.S.; Basu, S.; Alphey, L. Targeting female flight for genetic control of mosquitoes. PLoS Negl. Trop. Dis. 2020, 14, e0008876. [Google Scholar] [CrossRef]
- Fu, G.; Lees, R.S.; Nimmo, D.; Aw, D.; Jin, L.; Gray, P.; Berendonk, T.U.; White-Cooper, H.; Scaife, S.; Phuc, H.K.; et al. Female-Specific flightless phenotype for mosquito control. Proc. Natl. Acad. Sci. USA 2010, 107, 4550–4554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resnik, D.B. Ethical issues in field trials of genetically modified disease-Resistant mosquitoes. Dev. World Bioeth. 2012, 14, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, R.G.; Denton, J.A.; Santucci, F.; Bryk, J.; Reed, F.A. Scientific Standards and the Regulation of Genetically Modified Insects. PLoS Negl. Trop. Dis. 2012, 6, e1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Yang, T.; Kandul, N.P.; Bui, M.; Gamez, S.; Raban, R.R.; Bennett, J.; Sánchez C, H.M.; Lanzaro, G.C.; Schmidt, H.; et al. Development of a confinable gene drive system in the human disease vector Aedes aegypti. Elife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Terradas, G.; McGraw, E.A. Using genetic variation in Aedes aegypti to identify candidate anti-Dengue virus genes. BMC Infect. Dis. 2019, 19, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, A.A.; Montgomery, B.L.; Popovici, J.; Iturbeormaetxe, I.; Johnson, P.H.; Muzzi, F.; Greenfield, M.; Durkan, M.; Leong, Y.S.; Dong, Y.; et al. Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature 2011, 476, 454–457. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Ross, P.A.; Rašić, G. Wolbachia strains for disease control: Ecological and evolutionary considerations. Evol. Appl. 2015, 8, 751–768. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H. Biology of Wolbachia. Annu. Rev. Èntomol. 1997, 42, 587–609. [Google Scholar] [CrossRef] [Green Version]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How many species are infected with Wolbachia? –A statistical analysis of current data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Rainey, S.M.; Shah, P.; Kohl, A.; Dietrich, I. Understanding the Wolbachia-Mediated inhibition of arboviruses in mosquitoes: Progress and challenges. J. Gen. Virol. 2014, 95, 517–530. [Google Scholar] [CrossRef]
- Walker, T.G.; Johnson, P.H.; Moreira, L.A.; Iturbeormaetxe, I.; Frentiu, F.D.; McMeniman, C.J.; Leong, Y.S.; Dong, Y.; Axford, J.K.; Kriesner, P.; et al. The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nat. Cell Biol. 2011, 476, 450–453. [Google Scholar] [CrossRef]
- Kriesner, P.; Hoffmann, A.A.; Lee, S.F.; Turelli, M.; Weeks, A.R. Rapid Sequential Spread of Two Wolbachia Variants in Drosophila simulans. PLoS Pathog. 2013, 9, e1003607. [Google Scholar] [CrossRef] [Green Version]
- Huigens, M.E.; Luck, R.F.; Klaassen, R.H.G.; Maas, M.F.P.M.; Timmermans, M.J.T.N.; Stouthamer, R. Infectious parthenogenesis. Nat. Cell Biol. 2000, 405, 178–179. [Google Scholar] [CrossRef]
- Zabalou, S.; Riegler, M.; Theodorakopoulou, M.; Stauffer, C.; Savakis, C.; Bourtzis, K. Wolbachia-Induced cytoplasmic incompatibility as a means for insect pest population control. Proc. Natl. Acad. Sci. USA 2004, 101, 15042–15045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinkins, S.P.; Braig, H.R.; O’Neill, S.L. Wolbachia superinfections and the expression of cytoplasmic incompatibility. Proc. R. Soc. B: Boil. Sci. 1995, 261, 325–330. [Google Scholar] [CrossRef]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia Symbiont in Aedes aegypti Limits Infection with Dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertig, M.; Wolbach, S.B. Studies on Rickettsia-Like Micro-Organisms in Insects. J. Med. Res. 1924, 44, 329–374.7. [Google Scholar] [PubMed]
- Zhou, W.; Rousset, F.; O’Neill, S.L. Phylogeny and PCR-Based classification of Wolbachia strains using wsp gene sequences. Proc. Biol. Sci. 1998, 265, 509–515. [Google Scholar] [CrossRef]
- Ant, T.H.; Herd, C.S.; Geoghegan, V.; Hoffmann, A.A.; Sinkins, S.P. The Wolbachia strain wAu provides highly efficient virus transmission blocking in Aedes aegypti. PLoS Pathog. 2018, 14, e1006815. [Google Scholar] [CrossRef]
- Ulrich, J.N.; Beier, J.C.; Devine, G.J.; Hugo, L.E. Heat Sensitivity of wMel Wolbachia during Aedes aegypti Development. PLoS Negl. Trop. Dis. 2016, 10, e0004873. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Le Nguyen, H.; Nguyen, T.Y.; Vu, S.N.; Tran, N.D.; Le, T.N.; Vien, Q.M.; Bui, T.C.; Le, H.T.; Kutcher, S.; et al. Field evaluation of the establishment potential of wmelpop Wolbachia in Australia and Vietnam for dengue control. Parasites Vectors 2015, 8, 563. [Google Scholar] [CrossRef] [Green Version]
- Mancini, M.V.; Herd, C.S.; Ant, T.H.; Murdochy, S.M.; Sinkins, S.P. Wolbachia strain wAu efficiently blocks arbovirus transmission in Aedes albopictus. PLoS Negl. Trop. Dis. 2020, 14, e0007926. [Google Scholar] [CrossRef]
- Hurk, A.F.v.d.; Hall-Mendelin, S.; Pyke, A.T.; Frentiu, F.D.; McElroy, K.; Day, A.; Higgs, S.; O’Neill, S.L. Impact of Wolbachia on Infection with Chikungunya and Yellow Fever Viruses in the Mosquito Vector Aedes aegypti. PLoS Negl. Trop. Dis. 2012, 6, e1892. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, N.M.; Kien, D.T.H.; Clapham, H.; Aguas, R.; Trung, V.T.; Chau, T.N.B.; Popovici, J.; Ryan, P.A.; O’Neill, S.L.; McGraw, E.A.; et al. Modeling the impact on virus transmission ofWolbachia-Mediated blocking of dengue virus infection ofAedes aegypti. Sci. Transl. Med. 2015, 7, 279ra37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeap, H.L.; Mee, P.; Walker, T.W.; Weeks, A.R.; Oneill, S.L.; Johnson, P.R.S.; Ritchie, S.A.; Richardson, K.M.; Doig, C.J.; Endersby, N.M.; et al. Dynamics of the “Popcorn” Wolbachia Infection in Outbred Aedes aegypti Informs Prospects for Mosquito Vector Control. Genetics 2011, 187, 583–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasgon, J.L.; Scott, T.W. Wolbachia and cytoplasmic incompatibility in the California Culex pipiens mosquito species complex: Parameter estimates and infection dynamics in natural populations. Genetics 2003, 165, 2029–2038. [Google Scholar]
- Almeida, F.d.; Moura, A.S.; Cardoso, A.F.; Winter, C.E.; Bijovsky, A.T.; Suesdek, L. Effects of Wolbachia on fitness of Culex quinquefasciatus (Diptera; Culicidae). Infect. Genet. Evol. 2011, 11, 2138–2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyer, K.A.; Jaenike, J. Evolutionarily stable infection by a male-killing endosymbiont in Drosophila innubila: Molecular evidence from the host and parasite genomes. Genetics 2004, 168, 1443–1455. [Google Scholar] [CrossRef] [Green Version]
- Axford, J.K.; Callahan, A.G.; Hoffmann, A.A.; Yeap, H.L.; Ross, P.A. Fitness of wAlbB Wolbachia Infection in Aedes aegypti: Parameter Estimates in an Outcrossed Background and Potential for Population Invasion. Am. J. Trop. Med. Hyg. 2016, 94, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Parzych, K.R.; Klionsky, D.J. An Overview of Autophagy: Morphology, Mechanism, and Regulation. Antioxid. Redox Signal. 2014, 20, 460–473. [Google Scholar] [CrossRef] [Green Version]
- Voronin, D.; Cook, D.A.N.; Steven, A.; Taylor, M.J. Autophagy regulates Wolbachia populations across diverse symbiotic associations. Proc. Natl. Acad. Sci. USA 2012, 109, E1638–E1646. [Google Scholar] [CrossRef] [Green Version]
- Krejbich-Trotot, P.; Gay, B.; Li-Pat-Yuen, G.; Hoarau, J.-J.; Jaffar-Bandjee, M.-C.; Briant, L.; Gasque, P.; Denizot, M. Chikungunya triggers an autophagic process which promotes viral replication. Virol. J. 2011, 8, 432. [Google Scholar] [CrossRef] [Green Version]
- Gill, A.C.; Darby, A.C.; Makepeace, B.L. Iron Necessity: The Secret of Wolbachia’s Success? PLoS Negl. Trop. Dis. 2014, 8, e3224. [Google Scholar] [CrossRef] [Green Version]
- Zug, R.; Hammerstein, P. Bad guys turned nice? A critical assessment of Wolbachia mutualisms in arthropod hosts. Biol. Rev. Camb. Philos. Soc. 2015, 90, 89–111. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.; Jupatanakul, N.; Dimopoulos, G. Mosquito Immunity against Arboviruses. Viruses 2014, 6, 4479–4504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rancès, E.; Ye, Y.H.; Woolfit, M.; McGraw, E.A.; O’Neill, S.L. The Relative Importance of Innate Immune Priming in Wolbachia-Mediated Dengue Interference. PLoS Pathog. 2012, 8, e1002548. [Google Scholar] [CrossRef] [Green Version]
- Jupatanakul, N.; Sim, S.; Angleró-Rodríguez, Y.I.; Souza-Neto, J.; Das, S.; Poti, K.E.; Rossi, S.L.; Bergren, N.; Vasilakis, N.; Dimopoulos, G. Engineered Aedes aegypti JAK/STAT Pathway-Mediated Immunity to Dengue Virus. PLoS Negl. Trop. Dis. 2017, 11, e0005187. [Google Scholar] [CrossRef] [PubMed]
- Vargas, V.; Cime-Castillo, J.; Lanz-Mendoza, H. Immune priming with inactive dengue virus during the larval stage of Aedes aegypti protects against the infection in adult mosquitoes. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.; Kenny, N.; Eyles, D.W.; Moreira, L.A.; O’Neill, S.L.; Asgari, S. Infection with the wMel and wMelPop strains of Wolbachia leads to higher levels of melanization in the hemolymph of Drosophila melanogaster, Drosophila simulans and Aedes aegypti. Dev. Comp. Immunol. 2011, 35, 360–365. [Google Scholar] [CrossRef]
- Rodriguez-Andres, J.; Rani, S.; Varjak, M.; Chase-Topping, M.E.; Beck, M.H.; Ferguson, M.C.; Schnettler, E.; Fragkoudis, R.; Barry, G.; Merits, A.; et al. Phenoloxidase Activity Acts as a Mosquito Innate Immune Response against Infection with Semliki Forest Virus. PLoS Pathog. 2012, 8, e1002977. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.; Frentiu, F.D.; Moreira, L.A.; O’Neill, S.L.; Asgari, S. Wolbachia uses host microRNAs to manipulate host gene expression and facilitate colonization of the dengue vector Aedes aegypti. Proc. Natl. Acad. Sci. USA 2011, 108, 9250–9255. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.S.; Webster, J.A.; Madzokere, E.T.; Skinner, E.B.; Herrero, L.J. Mosquito antiviral defense mechanisms: A delicate balance between innate immunity and persistent viral infection. Parasites Vectors 2019, 12, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Saldaña, M.A.; Etebari, K.; Hart, C.E.; Widen, S.G.; Wood, T.G.; Thangamani, S.; Asgari, S.; Hughes, G.L. Zika virus alters the microRNA expression profile and elicits an RNAi response in Aedes aegypti mosquitoes. PLoS Negl. Trop. Dis. 2017, 11, e0005760. [Google Scholar] [CrossRef]
- Campbell, C.L.; Harrison, T.; Hess, A.M.; Ebel, G.D. MicroRNA levels are modulated inAedes aegyptiafter exposure to Dengue-2. Insect Mol. Biol. 2013, 23, 132–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMeniman, C.J.; Lane, R.V.; Cass, B.N.; Fong, A.W.C.; Sidhu, M.; Wang, Y.-F.; O’Neill, S.L. Stable Introduction of a Life-Shortening Wolbachia Infection into the Mosquito Aedes aegypti. Science 2009, 323, 141–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duron, O.; Bernard, C.; Unal, S.; Berthomieu, A.; Berticat, C.; Weill, M. Tracking factors modulating cytoplasmic incompatibilities in the mosquito Culex pipiens. Mol. Ecol. 2006, 15, 3061–3071. [Google Scholar] [CrossRef]
- Turelli, M.; Hoffmann, A.A. Cytoplasmic Incompatibility in Drosophila Simulans: Dynamics and Parameter Estimates from Natural Populations. Genetics 1995, 140, 1319–1338. [Google Scholar] [PubMed]
- Rousset, F.; Raymond, M. Cytoplasmic incompatibility in insects: Why sterilize females? Trends Ecol. Evol. 1991, 6, 54–57. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Clancy, D.J.; Merton, E. Cytoplasmic Incompatibility in Australian Populations of Drosophi-La-Melanogaster. Genetics 1994, 136, 993–999. [Google Scholar]
- Flor, M.; Hammerstein, P.; Telschow, A. Wolbachia-Induced unidirectional cytoplasmic incompatibility and the stability of infection polymorphism in parapatric host populations. J. Evol. Biol. 2007, 20, 696–706. [Google Scholar] [CrossRef]
- Telschow, A.; Flor, M.; Kobayashi, Y.; Hammerstein, P.; Werren, J.H. Wolbachia-Induced Unidirectional Cytoplasmic Incompatibility and Speciation: Mainland-Island Model. PLoS ONE 2007, 2, e701. [Google Scholar] [CrossRef] [Green Version]
- Branca, A.; Vavre, F.; Silvain, J.-F.; Dupas, S. Maintenance of adaptive differentiation by Wolbachia induced bidirectional cytoplasmic incompatibility: The importance of sib-Mating and genetic systems. BMC Evol. Biol. 2009, 9, 185. [Google Scholar] [CrossRef] [Green Version]
- Sicard, M.; Bouchon, D.; Ceyrac, L.; Raimond, R.; Thierry, M.; Clec’H, W.L.; Marcadé, I.; Caubet, Y.; Grève, P. Bidirectional cytoplasmic incompatibility caused by Wolbachia in the terrestrial isopod Porcellio dilatatus. J. Invertebr. Pathol. 2014, 121, 28–36. [Google Scholar] [CrossRef]
- Zhong, Y.; Li, Z.-X. Bidirectional Cytoplasmic Incompatibility Induced by Cross-Order Transfection of Wolbachia: Implications for Control of the Host Population. Microb. Ecol. 2014, 68, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Dorigatti, I.; McCormack, C.; Nedjati-Gilani, G.; Ferguson, N.M. Using Wolbachia for Dengue Control: Insights from Modelling. Trends Parasitol. 2018, 34, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Ogunlade, S.T.; Adekunle, A.I.; Meehan, M.T.; Rojas, D.P.; McBryde, E.S. Modeling the potential of wAu-Wolbachia strain invasion in mosquitoes to control Aedes-Borne arboviral infections. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.A.; Ritchie, S.A.; Axford, J.K.; Hoffmann, A.A. Loss of cytoplasmic incompatibility in Wolbachia-Infected Aedes aegypti under field conditions. PLoS Negl. Trop. Dis. 2019, 13, e0007357. [Google Scholar] [CrossRef]
- Adekunle, A.I.; Meehan, M.T.; McBryde, E.S. Mathematical analysis of a Wolbachia invasive model with imperfect maternal transmission and loss of Wolbachia infection. Infect. Dis. Model. 2019, 4, 265–285. [Google Scholar] [CrossRef]
- Dutton, T.J.; Sinkins, S.P. Strain-Specific quantification of Wolbachia density in Aedes albopictus and effects of larval rearing conditions. Insect Mol. Biol. 2004, 13, 317–322. [Google Scholar] [CrossRef]
- Hague, M.T.J.; Mavengere, H.; Matute, D.R.; Cooper, B.S. Environmental and Genetic Contributions to Imperfect wMel-Like Wolbachia Transmission and Frequency Variation. Genetics 2020, 215, 1117–1132. [Google Scholar] [CrossRef]
- Meany, M.K.; Conner, W.R.; Richter, S.V.; Bailey, J.A.; Turelli, M.; Cooper, B.S. Loss of cytoplasmic incompatibility and minimal fecundity effects explain relatively low Wolbachia frequencies in Drosophila mauritiana. Evolution 2019, 73, 1278–1295. [Google Scholar] [CrossRef] [Green Version]
- Hedges, L.M.; Brownlie, J.C.; O’Neill, S.L.; Johnson, K.N. Wolbachia and Virus Protection in Insects. Science 2008, 322, 702. [Google Scholar] [CrossRef]
- Shaw, A.E.; Veronesi, E.; Maurin, G.; Ftaich, N.; Guiguen, F.; Rixon, F.; Ratinier, M.; Mertens, P.; Carpenter, S.; Palmarini, M.; et al. Drosophila melanogaster as a Model Organism for Bluetongue Virus Replication and Tropism. J. Virol. 2012, 86, 9015–9024. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, L.A.; Ferreira, Á.; Ashburner, M. The Bacterial Symbiont Wolbachia Induces Resistance to RNA Viral Infections in Drosophila melanogaster. PLoS Biol. 2008, 6, e1000002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kambris, Z.; Cook, P.E.; Phuc, H.K.; Sinkins, S.P. Immune Activation by Life-Shortening Wolbachia and Reduced Filarial Competence in Mosquitoes. Science 2009, 326, 134–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, L.; Plichart, C.; Sang, A.C.; Brelsfoard, C.L.; Bossin, H.C.; Dobson, S.L. Open release of male mosquitoes in-Fected with a wolbachia biopesticide: Field performance and infection containment. PLoS Negl. Trop. Dis. 2012, 6, e1797. [Google Scholar]
- Rasic, G.; Filipovic, I.; Weeks, A.R.; Hoffmann, A.A. Genome-Wide SNPs lead to strong signals of geographic structure and relatedness patterns in the major arbovirus vector, Aedes aegypti. BMC Genom. 2014, 15, 275. [Google Scholar] [CrossRef] [Green Version]
- Frentiu, F.D.; Zakir, T.; Walker, T.; Popovici, J.; Pyke, A.T.; Hurk, A.V.D.; McGraw, E.A.; O’Neill, S.L. Limited Dengue Virus Replication in Field-Collected Aedes aegypti Mosquitoes Infected with Wolbachia. PLoS Negl. Trop. Dis. 2014, 8, e2688. [Google Scholar] [CrossRef] [Green Version]
- WHO. World Health Organization Mosquito Control: Can. It Stop Zika at Source? WHO: Geneva, Switzerland, 2016; Available online: http://www.who.int/emergencies/zika-virus/articles/mosquito-control/en/ (accessed on 17 October 2019).
- Pan, X.; Pike, A.; Joshi, D.; Bian, G.; McFadden, M.J.; Lu, P.; Liang, X.; Zhang, F.; Raikhel, A.S.; Xiaoling, P. The bacterium Wolbachia exploits host innate immunity to establish a symbiotic relationship with the dengue vector mosquito Aedes aegypti. ISME J. 2018, 12, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Wang, Y.; He, K.; Yang, Q.; Gong, M.; Ji, M.; Chen, L. Wolbachia limits pathogen infections through induction of host innate immune responses. PLoS ONE 2020, 15, e0226736. [Google Scholar] [CrossRef] [Green Version]
- Turley, A.P.; Moreira, L.A.; O’Neill, S.L.; McGraw, E.A. Wolbachia Infection Reduces Blood-Feeding Success in the Dengue Fever Mosquito, Aedes aegypti. PLoS Negl. Trop. Dis. 2009, 3, e516. [Google Scholar] [CrossRef]
- Sutton, E.R.; Harris, S.R.; Parkhill, J.; Sinkins, S.P. Comparative genome analysis of Wolbachia strain wAu. BMC Genom. 2014, 15, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Suhrcke, M.; Stuckler, D.; Suk, J.E.; Desai, M.; Senek, M.; McKee, M.; Tsolova, S.; Basu, S.; Abubakar, I.; Hunter, P.; et al. The Impact of Economic Crises on Communicable Disease Transmission and Control: A Systematic Review of the Evidence. PLoS ONE 2011, 6, e20724. [Google Scholar] [CrossRef]
- Anderson, R.M.; May, R.M.; Gupta, S. Non-Iinear phenomena in host—Parasite interactions. Parasitology 1989, 99, S59–S79. [Google Scholar] [CrossRef] [PubMed]
- May, R.M. Infectious Disease: Can We Avert a Lethal Flu Pandemic? Curr. Biol. 2005, 15, R922–R924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hethcote, H.W.; Driessche, P.V.D. Two SIS epidemiologic models with delays. J. Math. Biol. 2000, 40, 3–26. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.M.; May, R.M. Population biology of infectious diseases: Part I. Nat. Cell Biol. 1979, 280, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Siettos, C.; Russo, L. Mathematical modeling of infectious disease dynamics. Virulence 2013, 4, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Driessche, P.V.D.; Watmough, J. Reproduction numbers and sub-Threshold endemic equilibria for compartmental models of disease transmission. Math. Biosci. 2002, 180, 29–48. [Google Scholar] [CrossRef]
- Campo-Duarte, D.E.; Vasilieva, O.; Cardona-Salgado, D.; Svinin, M. Optimal control approach for establishing wMelPop Wolbachia infection among wild Aedes aegypti populations. J. Math. Biol. 2018, 76, 1907–1950. [Google Scholar] [CrossRef]
- Hughes, H.; Britton, N.F. Modelling the Use of Wolbachia to Control Dengue Fever Transmission. Bull. Math. Biol. 2013, 75, 796–818. [Google Scholar] [CrossRef] [Green Version]
- Ndii, M.Z.; Hickson, R.I.; Mercer, G.N. Modelling the Introduction of Wolbachia into Aedes Aegypti Mosquitoes to Reduce Dengue Transmission. ANZIAM J. 2012, 53, 213–227. [Google Scholar] [CrossRef] [Green Version]
- Qu, Z.; Xue, L.; Hyman, J.M. Modeling the Transmission of Wolbachia in Mosquitoes for Controlling Mosquito-Borne Diseases. SIAM J. Appl. Math. 2018, 78, 826–852. [Google Scholar] [CrossRef] [Green Version]
- Schraiber, J.G.; Kaczmarczyk, A.N.; Kwok, R.; Park, M.; Silverstein, R.; Rutaganira, F.U.; Aggarwal, T.; Schwemmer, M.A.; Hom, C.L.; Grosberg, R.K.; et al. Constraints on the use of lifespan-Shortening Wolbachia to control dengue fever. J. Theor. Biol. 2012, 297, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Telschow, A.; Yamamura, N.; Werren, J.H. Bidirectional cytoplasmic incompatibility and the stable coexistence of two Wolbachia strains in parapatric host populations. J. Theor. Biol. 2005, 235, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Manore, C.A.; Thongsripong, P.; Hyman, J.M. Two-Sex mosquito model for the persistence ofWolbachia. J. Biol. Dyn. 2017, 11, 216–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, B.; Tang, M.; Yu, J.; Qiu, J. Wolbachia spreading dynamics in mosquitoes with imperfect maternal transmission. J. Math. Biol. 2018, 76, 235–263. [Google Scholar] [CrossRef] [PubMed]
- Gumel, A.B. Causes of backward bifurcations in some epidemiological models. J. Math. Anal. Appl. 2012, 395, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.C.D.S.; Maia, L.M.S.; De Souza, V.C.; Gonzaga, A.M.; De Azevedo, V.C.; Martins, L.R.; Pavoni, J.H.C.; Naveca, F.G.; Slhessarenko, R.D. Arbovirus investigation in patients from Mato Grosso during Zika and Chikungunya virus introdution in Brazil, 2015–2016. Acta Trop. 2019, 190, 395–402. [Google Scholar] [CrossRef]
- Eckerle, I.; Briciu, V.; Ergonul, O.; Lupse, M.; Papa, A.; Radulescu, A.; Tsiodras, S.; Tsitou, C.; Drosten, C.; Nussenblatt, V.; et al. Emerging souvenirs—Clinical presentation of the returning traveller with imported arbovirus infections in Europe. Clin. Microbiol. Infect. 2018, 24, 240–245. [Google Scholar] [CrossRef]
- Eder, M.; Cortes, F.; Filha, N.T.D.S.; De França, G.V.A.; DeGroote, S.; Braga, M.C.; Ridde, V.; Martelli, C.M.T. Scoping review on vector-Borne diseases in urban areas: Transmission dynamics, vectorial capacity and co-Infection. Infect. Dis. Poverty 2018, 7, 1–24. [Google Scholar] [CrossRef]
- Iwashita, H.; Higa, Y.; Futami, K.; Lutiali, P.A.; Njenga, S.M.; Nabeshima, T.; Minakawa, N. Mosquito arbovirus survey in selected areas of Kenya: Detection of insect-Specific virus. Trop. Med. Heal. 2018, 46, 1–15. [Google Scholar] [CrossRef]
- Ross, P.A.; Wiwatanaratanabutr, I.; Axford, J.K.; White, V.L.; Endersby-Harshman, N.M.; Hoffmann, A.A. Wolbachia Infections in Aedes aegypti Differ Markedly in Their Response to Cyclical Heat Stress. PLoS Pathog. 2017, 13, e1006006. [Google Scholar] [CrossRef] [Green Version]



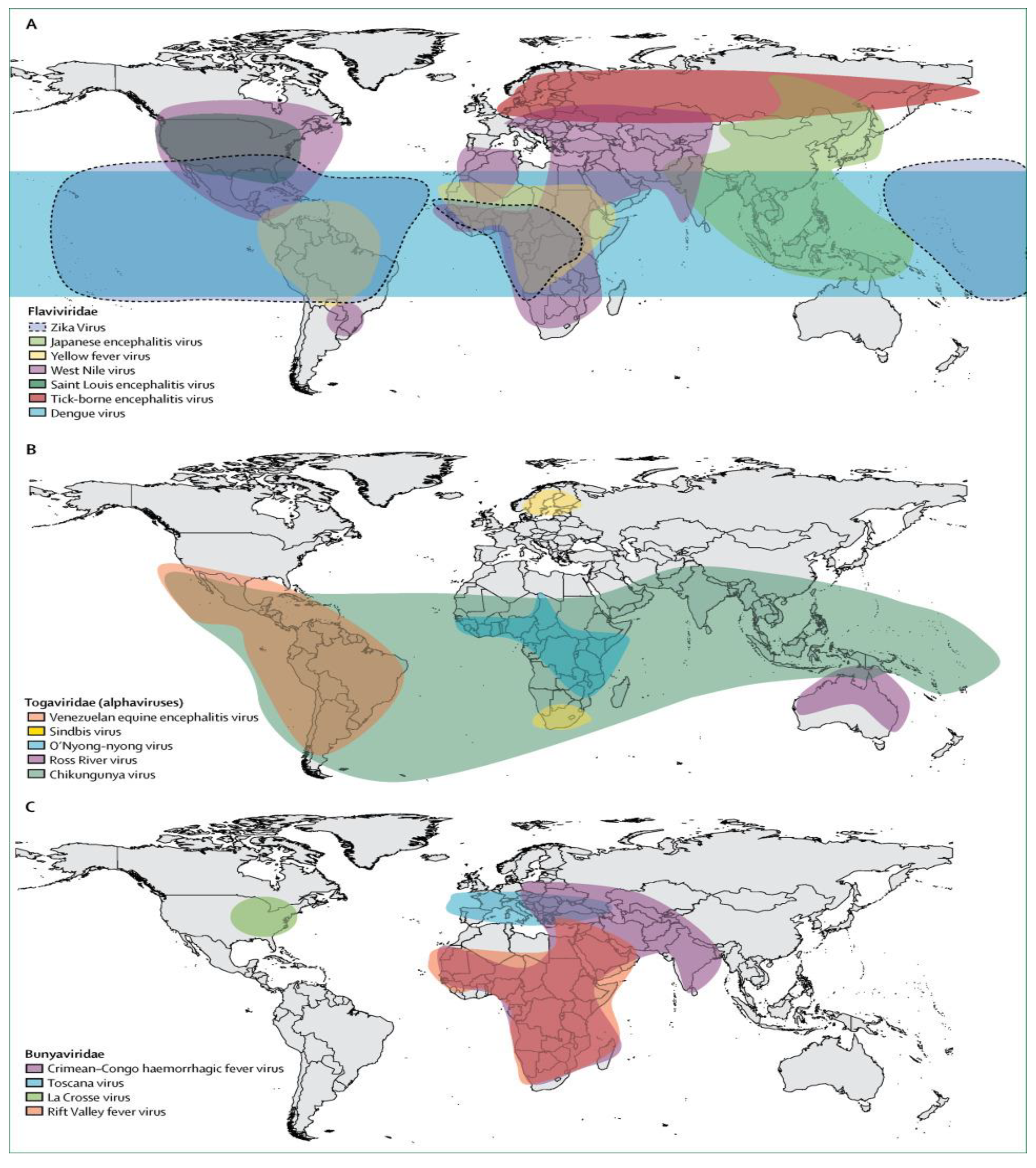{kind=link}
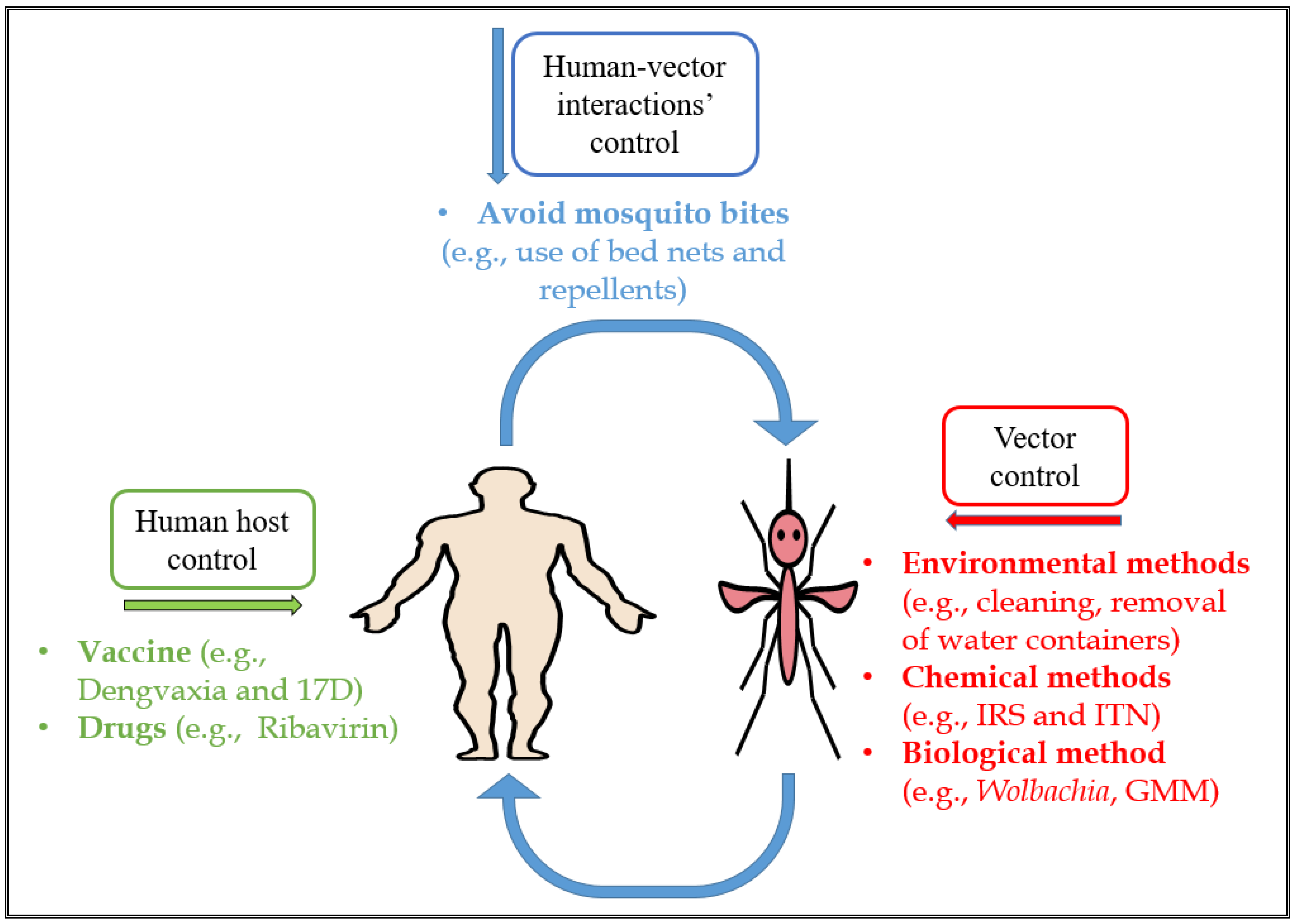{kind=link}
| Aedes-Borne Arboviruses | Virus Type | Transmitted By | Symptoms | Supportive Treatment | Coinfection with Other Arboviruses | Intrinsic Incubation Period (Days) | Extrinsic Incubation Period (Days) | Asymptomatic Proportion in Infected Humans (%) |
|---|---|---|---|---|---|---|---|---|
| Dengue | Flavivirus [14] | Ae. Aegypti Ae. albopictus [5,6] | Sudden high grade fever, Headache, Nausea, Arthralgia, Eye and Muscle pain [65] | DENV vaccine and drug administration [66] | Yes (e.g., DENV and ZIKV) [67] | Median: 5.3 [68] Mean: 5.9 [42] | Mean: 15 (at 25 °C) 6.5 (at 30 °C) [42] | 75 [69] |
| Zika | Flavivirus [70] | Ae. aegypti Ae. albopictus Human (via blood transfusion) [45,46,51] | Fever, Conjunctivitis, Muscle pain, Headache, Joint pain, Rash and Microcephaly [71,72] | Fluid intake and drug administration (such as acetaminophen) [73] | Yes (e.g., ZIKV and CHIKV) [74] | Median: 6.8 [75] 6.2 [76] | Median: 5.1 (at 30 °C) 9.6 (at 26 °C) 24.2 (at 21 °C) [77] | 80 [78] |
| Chikungunya | Alphavirus [79] | Ae. aegypti Ae. albopictus [80] | High fever, Joint pain, Myalgia, Arthritis, Conjunctivitis, and Dermatologic manifestations [81,82] | Plenty of rest, Fluid intake and Acetaminophen [52,80] | Yes (e.g., CHIKV and DENV) [83] | Median: 3.0 [68] | Median: 2 [84] | Approx. 18 to 28 [85] |
| Yellow Fever | Flavivirus [56] | Ae. aegypti Ae. Albopictus [86] | Headache, Nausea, Vomiting, Fever, Dizziness and Joint pain [87,88] | YFV vaccine and Ribavirin [89,90] | Yes (e.g., YFV and CHIKV) [91] | Median: 4.3 [92] 4.4 [68] | Median: 10 (at 25 °C) [92] | 55 [93] |
| Wolbachia Strain | Origin | Means of Control of Arboviral Transmission (CI, MT, WIR, VB, F) | References |
|---|---|---|---|
| wAu | Drosophila simulans/Fruit fly | (No, Yes, High, High, Partial) | [141,144] |
| wMel | Drosophila melanogaster/Fruit fly | (Yes, Yes, Low, Partial, Partial) | [31,133,145] |
| wAlbA | Aedes albopictus/Asian Tiger mosquito | (Yes, Yes, Medium, Partial, High) | [140,141] |
| wAlbB | Aedes albopictus/Asian Tiger mosquito | (Yes, Yes, Medium, High, Partial) | [129,141] |
| wMelPop | Drosophila melanogaster/Fruit fly | (Yes, Yes, Low, High, High) | [141,146,147] |
| wPip | Culex pipiens/Mosquito | (Yes, Yes, -, Low, Low) | [129,148,149] |
| wRi | Drosophila simulans (Riverside)/Fly | (Yes, Yes, -, Partial, Low) | [129,140] |
| wInn | Drosophila innubila/Vinegar fly | (Yes [only males], Yes, -, Partial, Low) | [129,150] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogunlade, S.T.; Meehan, M.T.; Adekunle, A.I.; Rojas, D.P.; Adegboye, O.A.; McBryde, E.S. A Review: Aedes-Borne Arboviral Infections, Controls and Wolbachia-Based Strategies. Vaccines 2021, 9, 32. https://doi.org/10.3390/vaccines9010032
Ogunlade ST, Meehan MT, Adekunle AI, Rojas DP, Adegboye OA, McBryde ES. A Review: Aedes-Borne Arboviral Infections, Controls and Wolbachia-Based Strategies. Vaccines. 2021; 9(1):32. https://doi.org/10.3390/vaccines9010032
Chicago/Turabian StyleOgunlade, Samson T., Michael T. Meehan, Adeshina I. Adekunle, Diana P. Rojas, Oyelola A. Adegboye, and Emma S. McBryde. 2021. "A Review: Aedes-Borne Arboviral Infections, Controls and Wolbachia-Based Strategies" Vaccines 9, no. 1: 32. https://doi.org/10.3390/vaccines9010032
APA StyleOgunlade, S. T., Meehan, M. T., Adekunle, A. I., Rojas, D. P., Adegboye, O. A., & McBryde, E. S. (2021). A Review: Aedes-Borne Arboviral Infections, Controls and Wolbachia-Based Strategies. Vaccines, 9(1), 32. https://doi.org/10.3390/vaccines9010032