
Correlation of Influenza B Haemagglutination Inhibiton, Single-Radial Haemolysis and Pseudotype-Based Microneutralisation Assays for Immunogenicity Testing of Seasonal Vaccines

 , ,
, ,  ,
, 
Abstract
:1. Introduction
1.1. Serological Assays for Influenza B
1.2. Monoclonal Antibodies
1.3. Study Aims
2. Materials and Methods
2.1. Plasmids
2.2. Serum Samples
2.3. Pseudotype Production
2.4. Pseudotype-Based Microneutralisation Assay
2.5. Haemagglutination Inhibition Assay
2.6. Single Radial Haemolysis Assay
2.7. mAbs and Controls
2.8. Statistical Analysis
3. Results
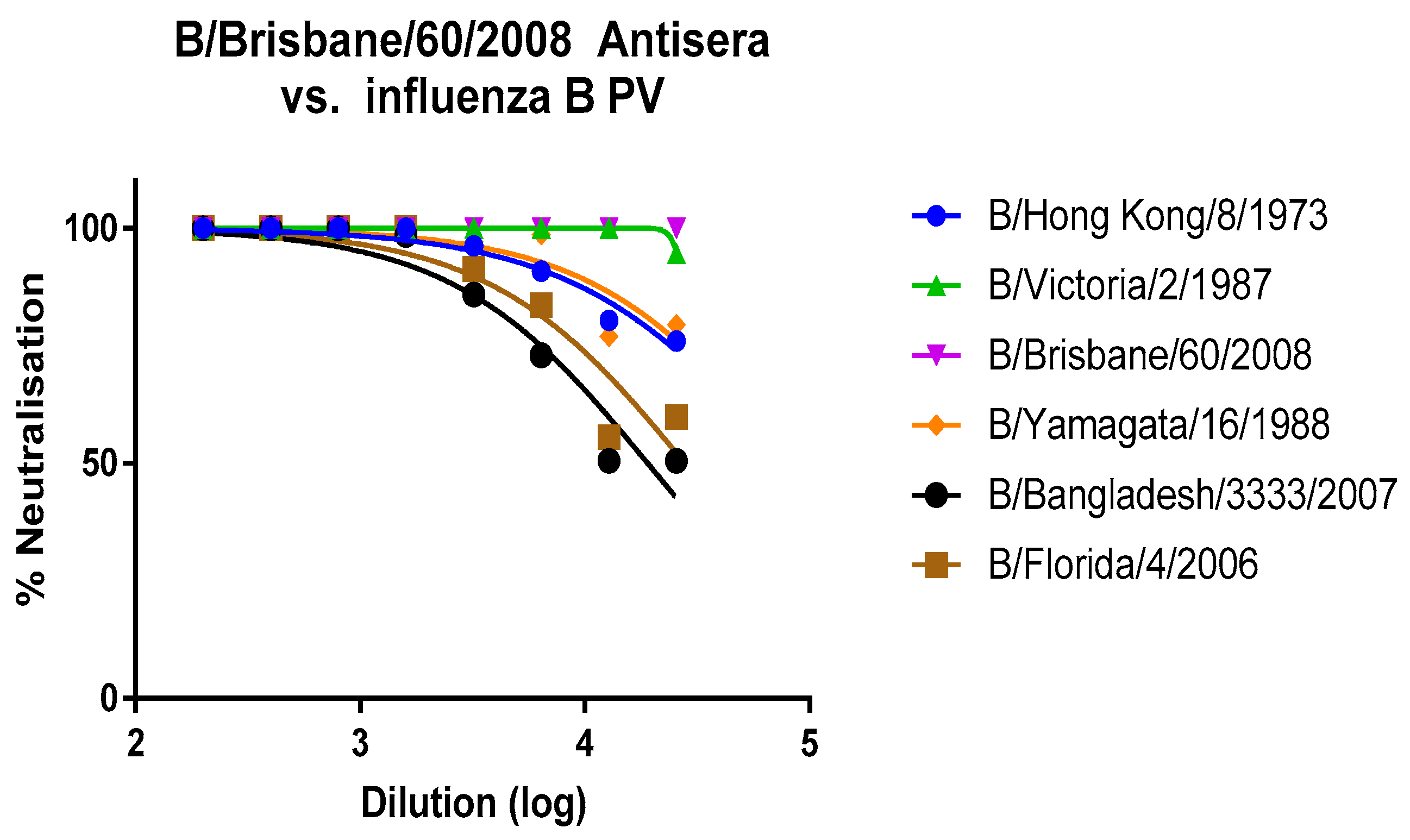
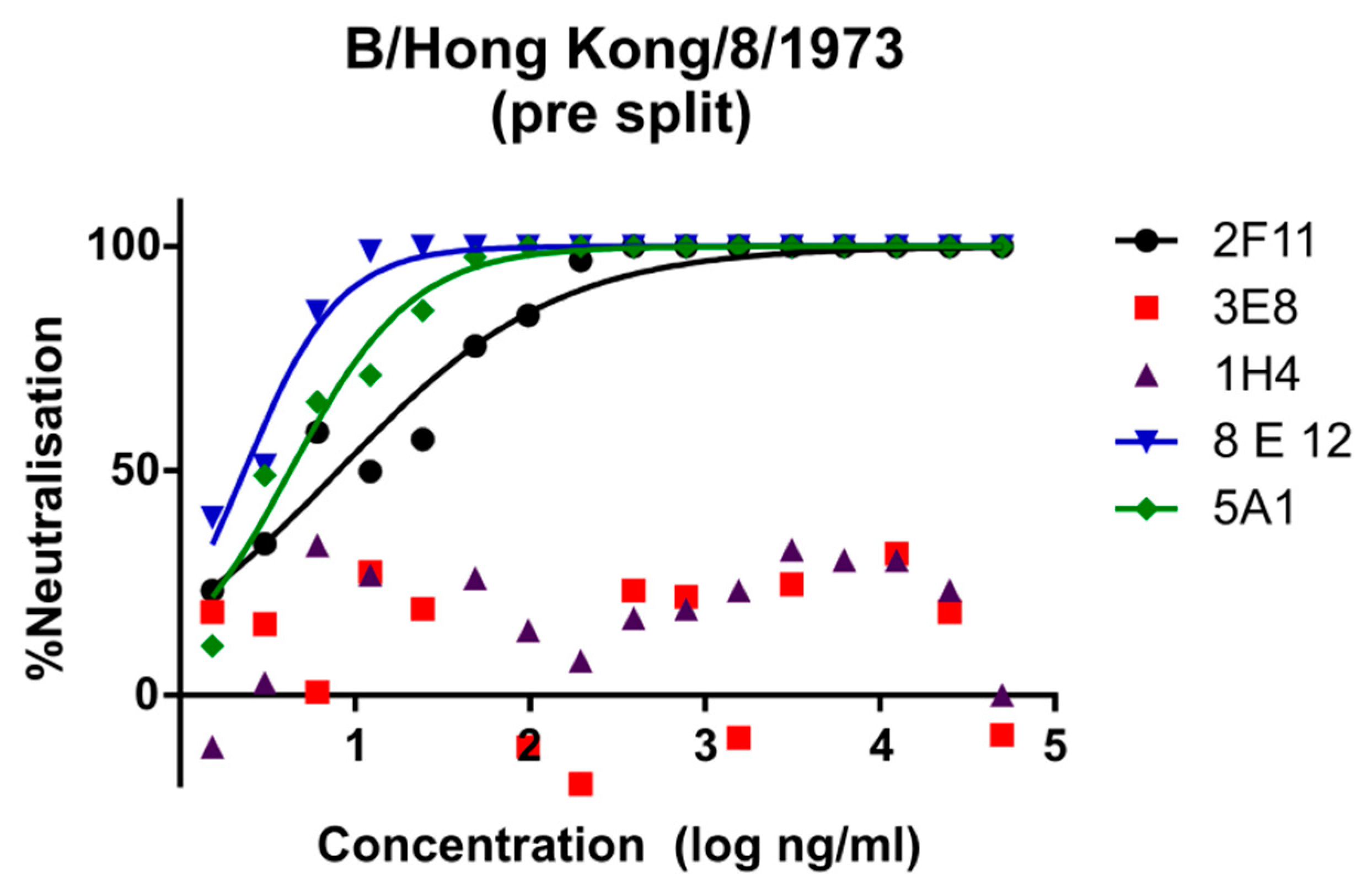
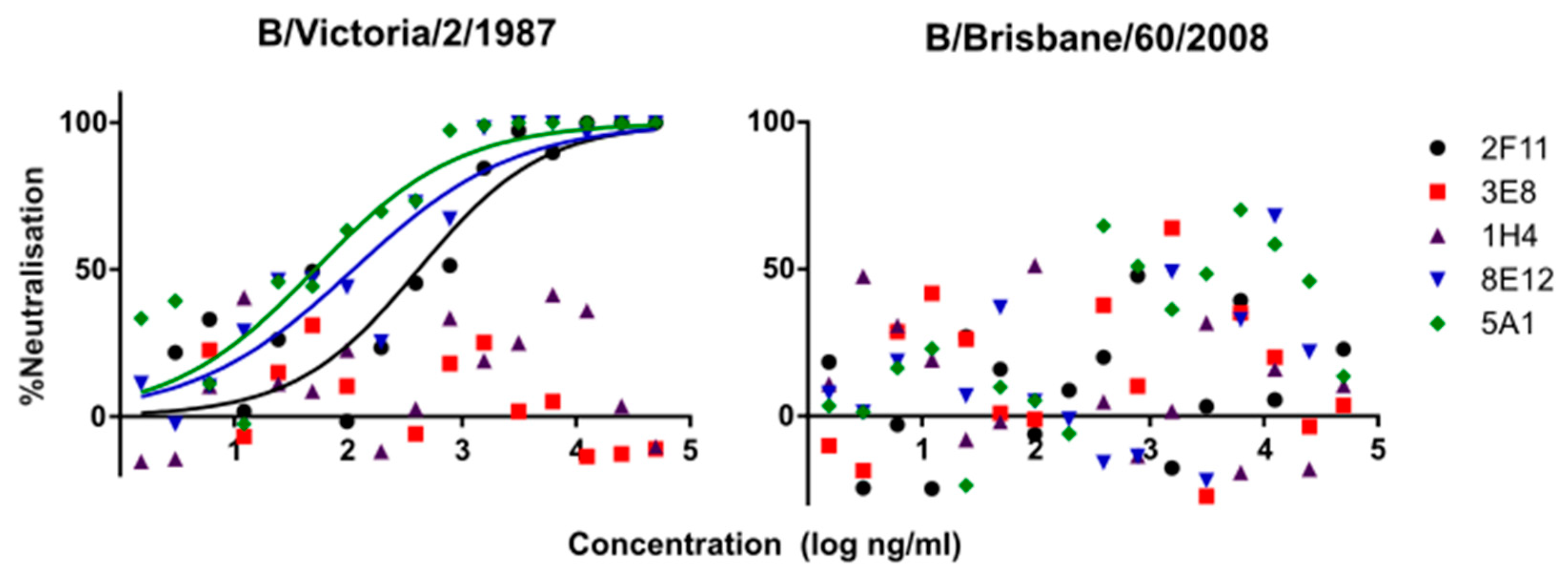
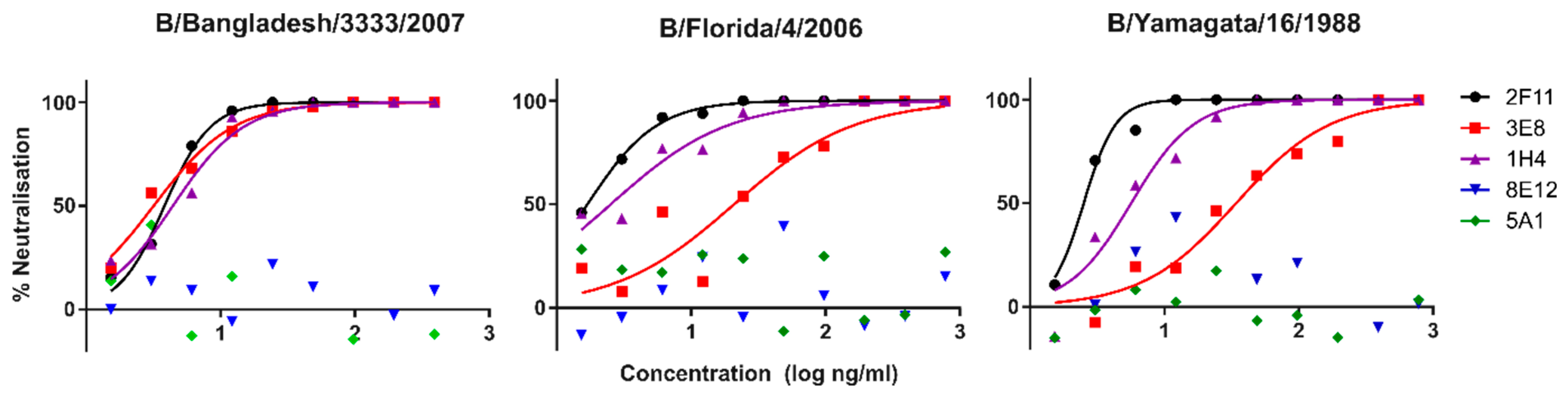
3.1. Lineage Specific and Cross-Reacting mAbs Neutralise Influenza B PV
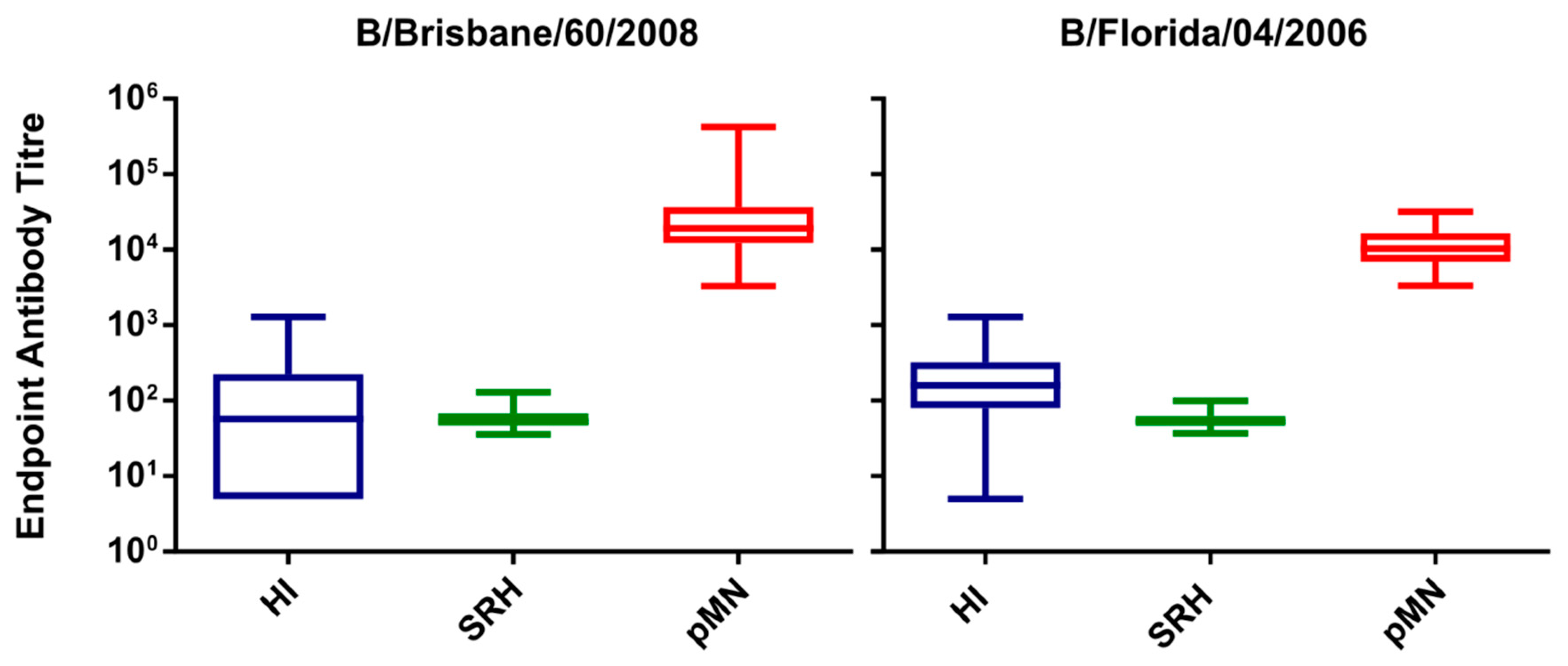
3.2. Correlation of SRH, HI and pMN
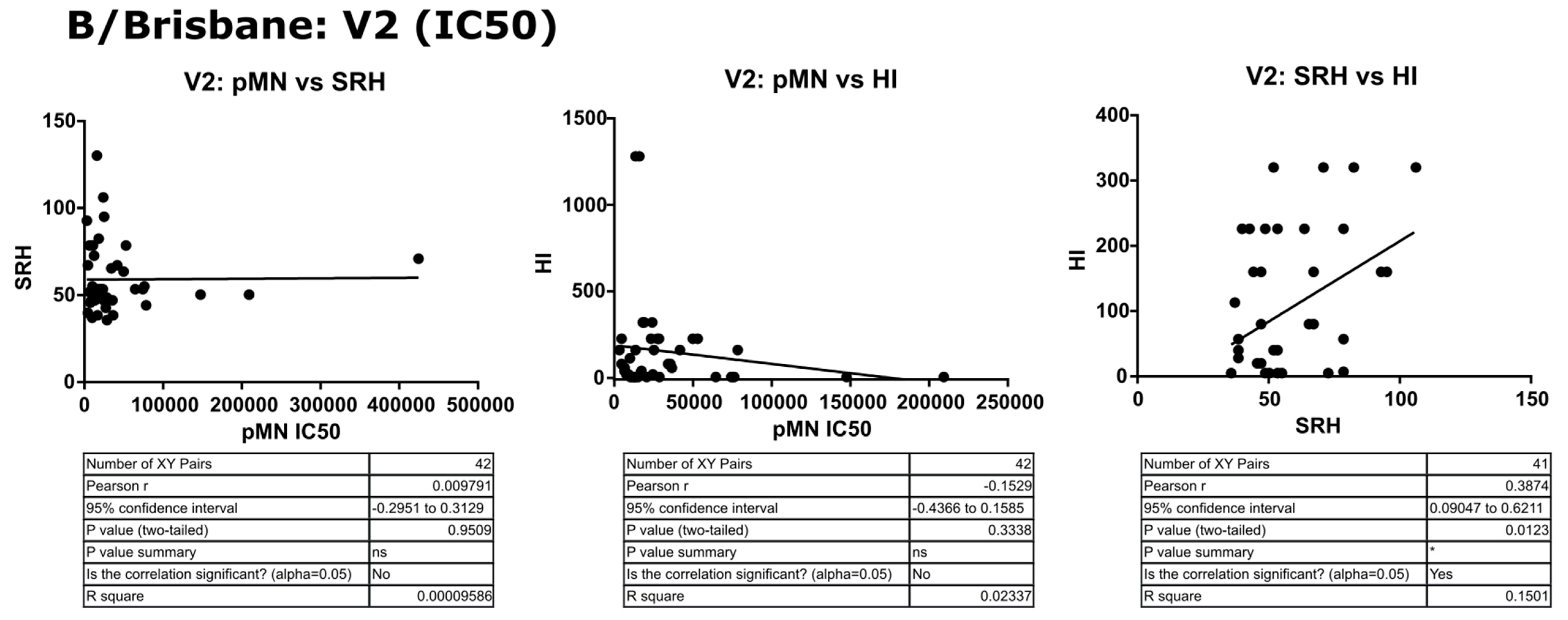
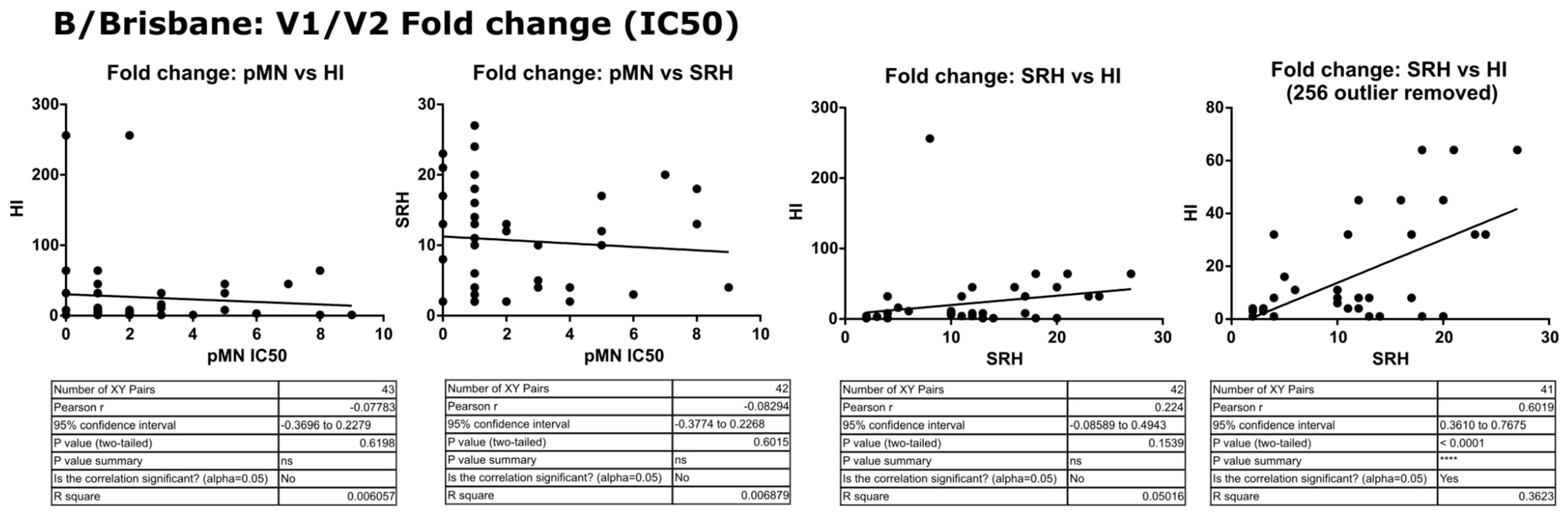
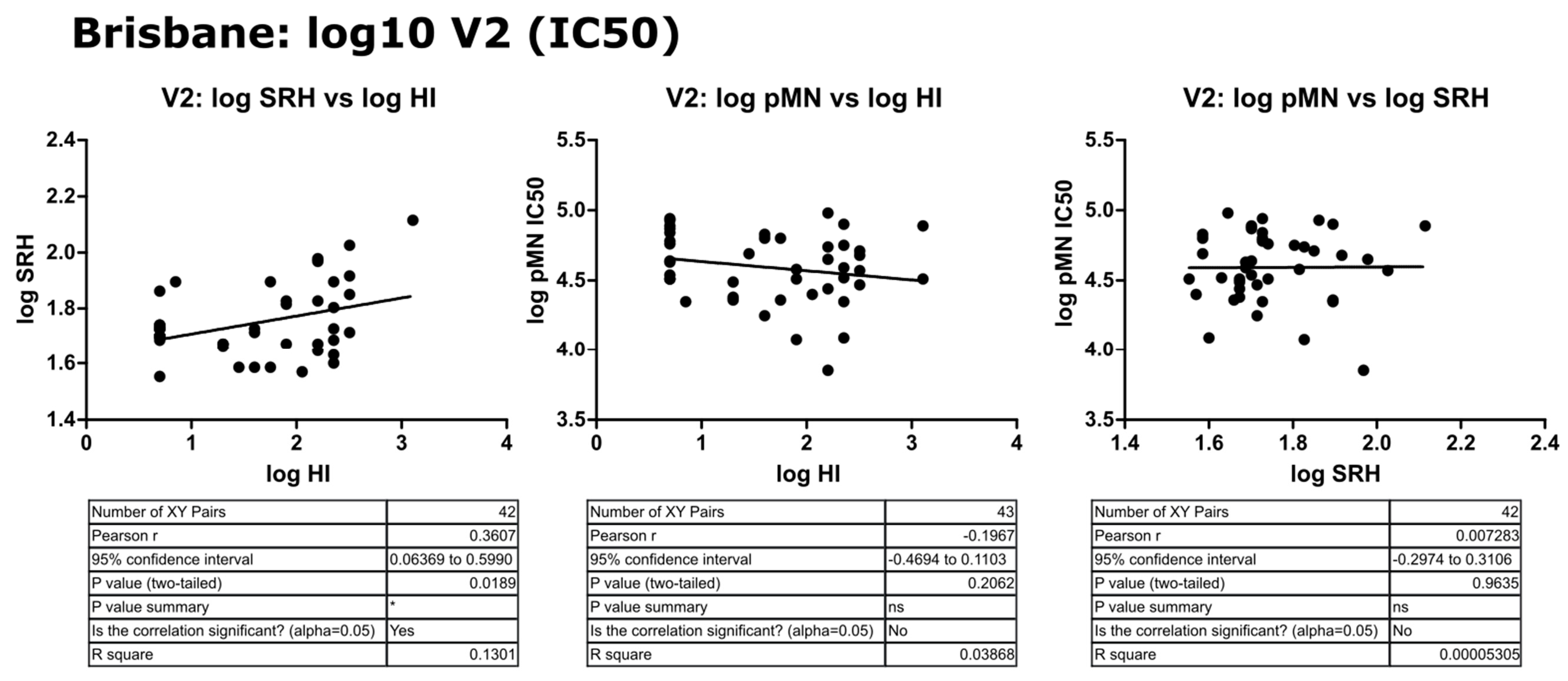
3.3. Correlation of Data: B/Brisbane/60/2008 IC50
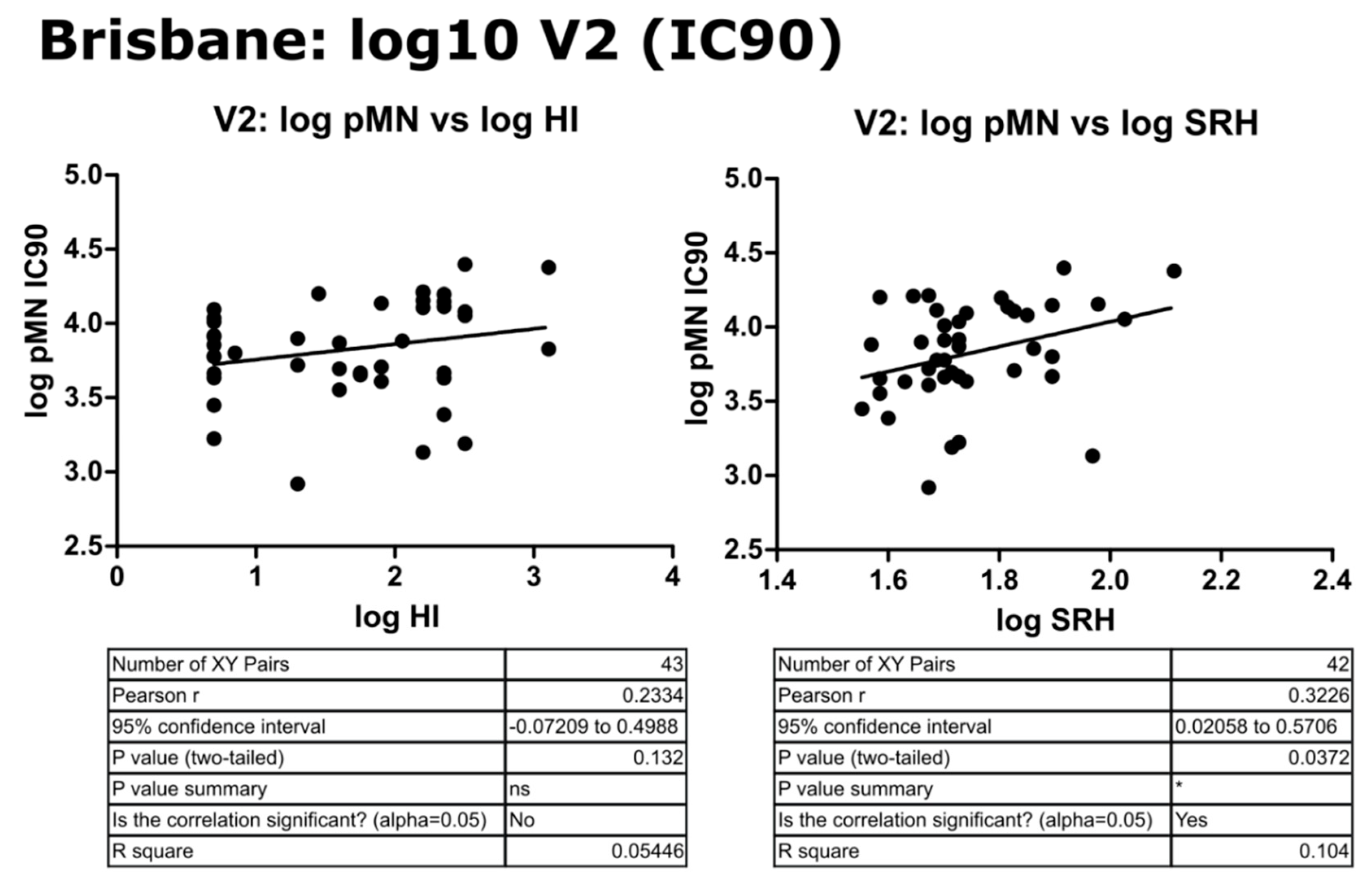
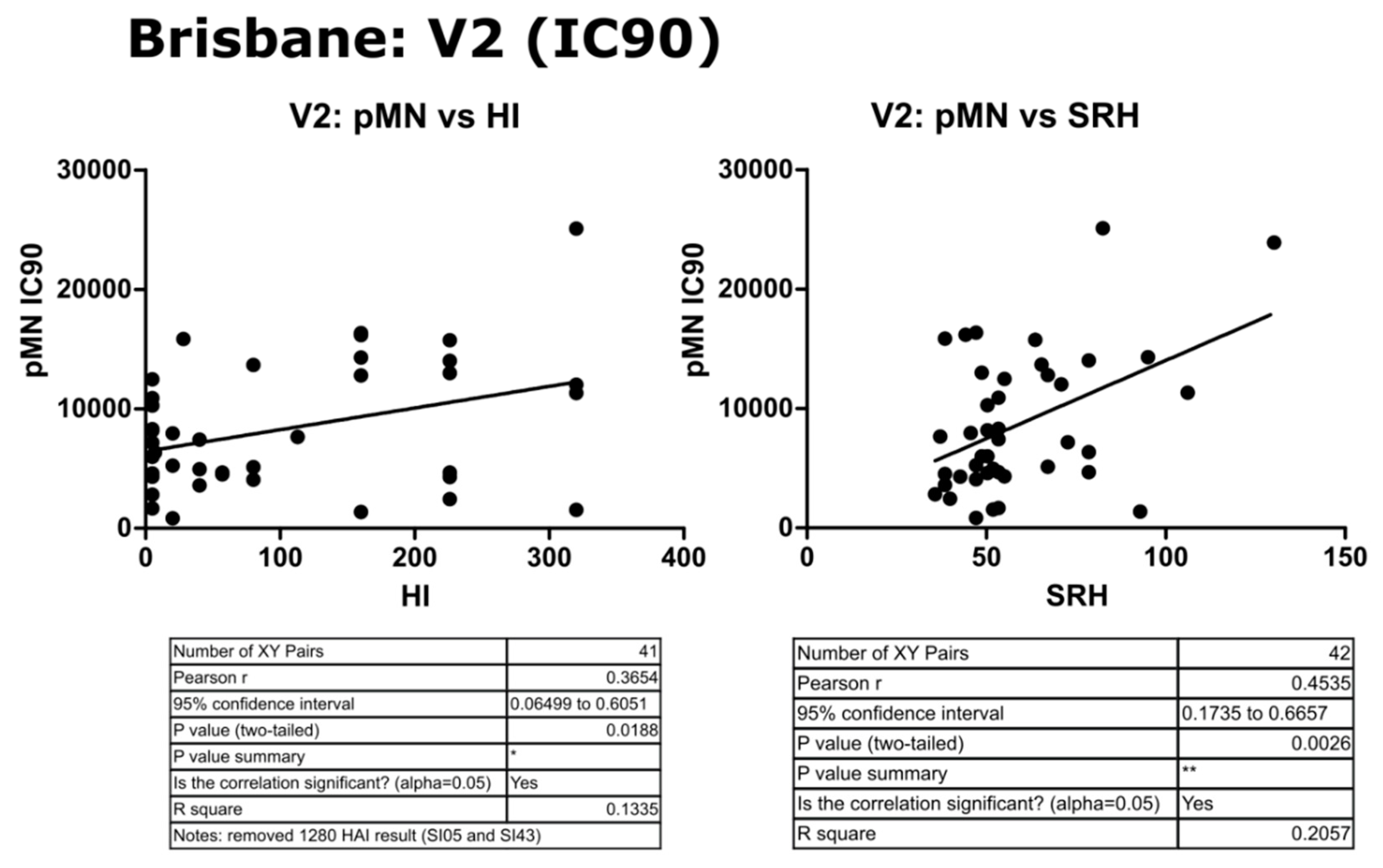
3.4. Correlation of Data: B/Brisbane IC90 and Transformed Data
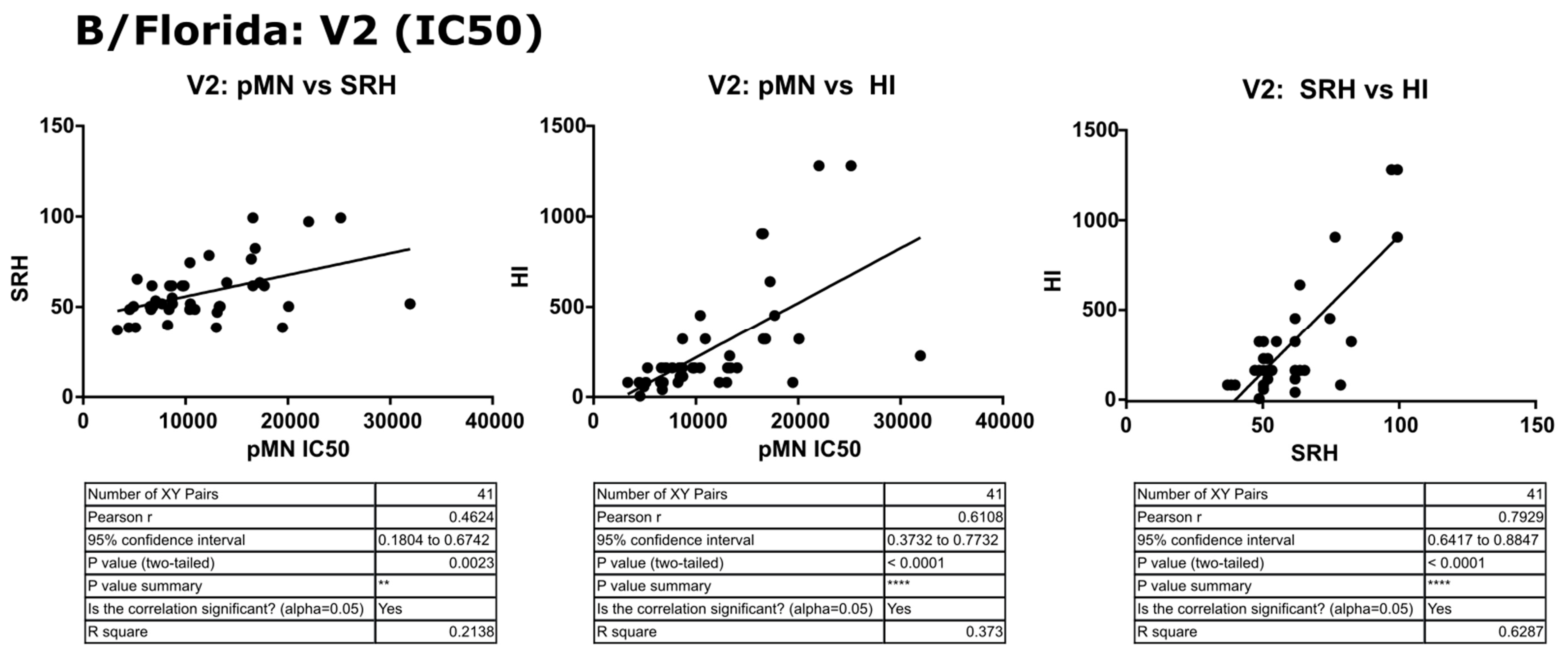
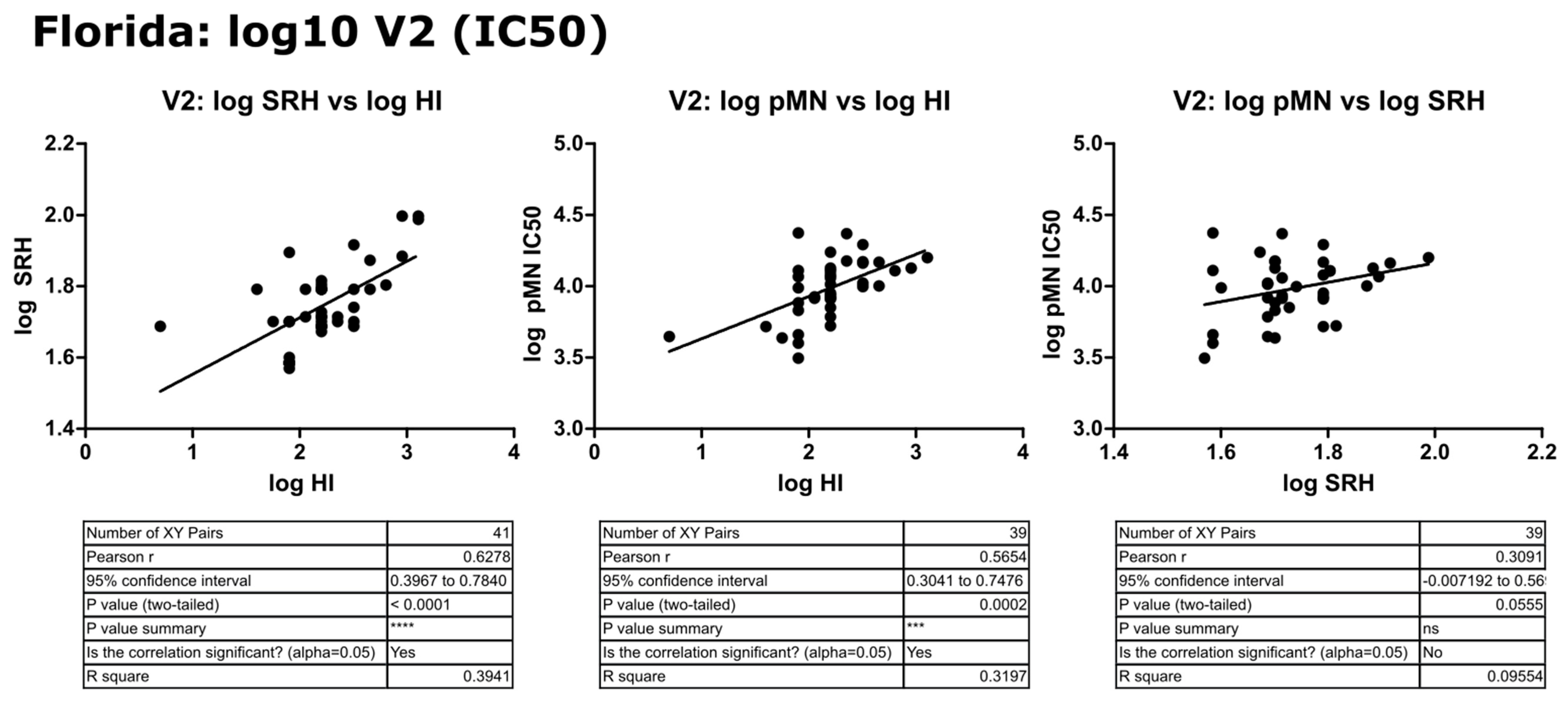
3.5. Correlation of Data: B/Florida/4/2006 IC50
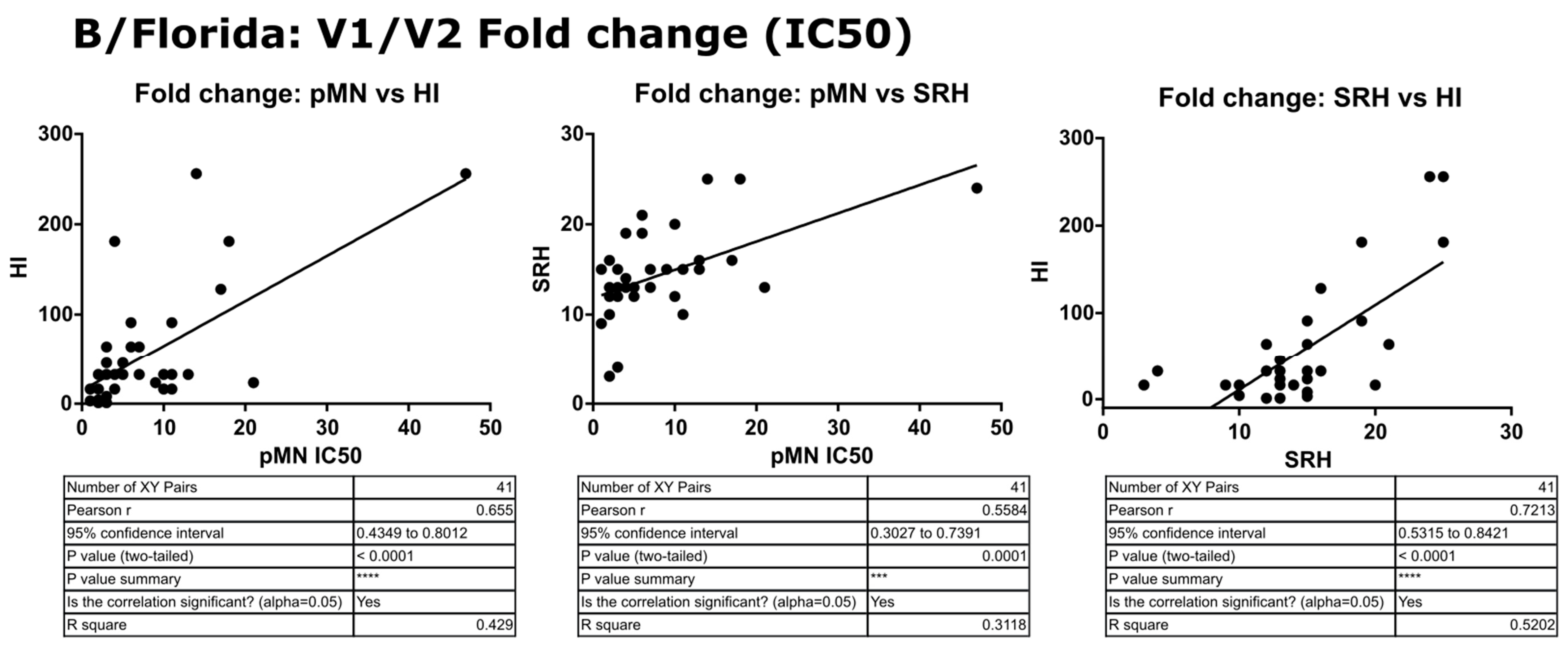
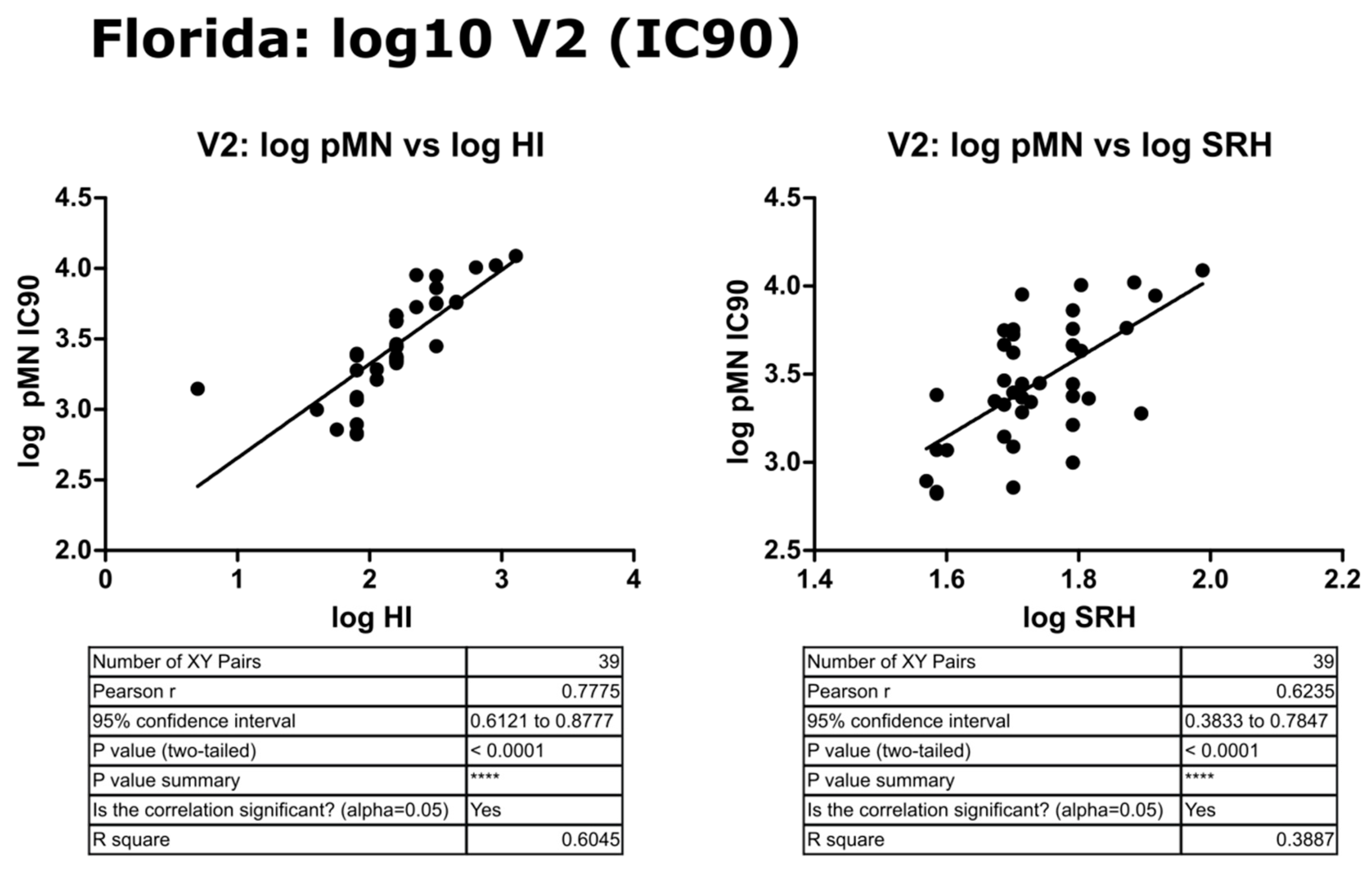
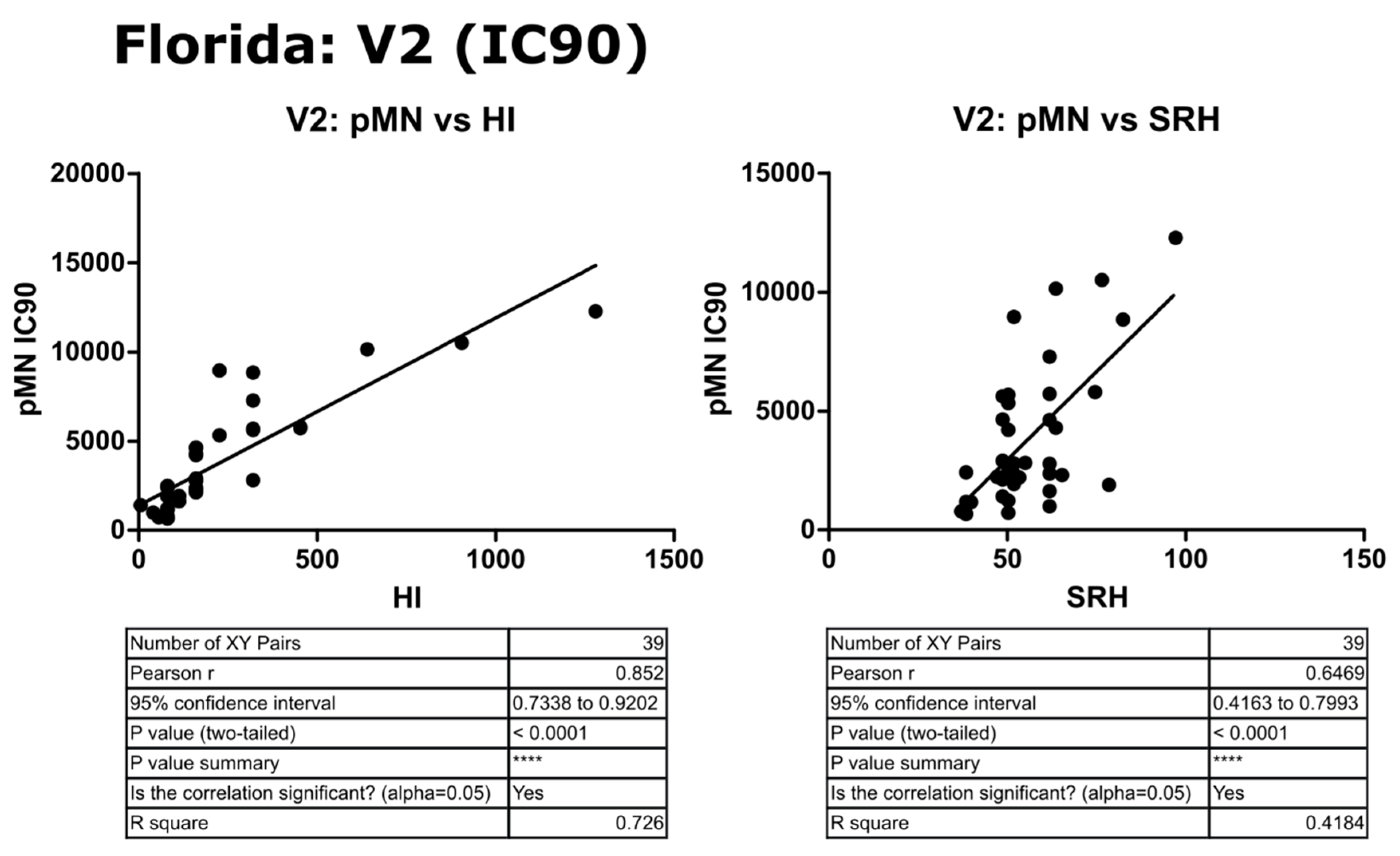
3.6. Correlation of Data: B/Florida/4/2006 IC90 and Transformed Data
4. Discussion
4.1. Neutralisation by Influenza B mAbs
4.2. Correlation between SRH and HI
4.3. Correlation between pMN and SRH
4.4. Correlation between pMN and HI
4.5. Limitations of This Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hause, B.M.; Collin, E.A.; Liu, R.; Huang, B.; Sheng, Z.; Lu, W.; Wang, D.; Nelson, E.A.; Li, F. Characterization of a novel influenza virus in cattle and swine: Proposal for a new genus in the Orthomyxoviridae family. MBio 2014, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rota, P.A.; Wallis, T.R.; Harmon, M.W.; Rota, J.S.; Kendal, A.P.; Nerome, K. Cocirculation of two distinct evolutionary lineages of influenza type B virus since 1983. Virology 1990, 175, 59–68. [Google Scholar] [CrossRef]
- Ambrose, C.S.; Levin, M.J. The rationale for quadrivalent influenza vaccines. Hum. Vaccines Immunother. 2012, 8, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Glezen, W.P.; Schmier, J.K.; Kuehn, C.M.; Ryan, K.J.; Oxford, J.S. The burden of influenza B: A structured literature review. Am. J. Public Health 2013, 103, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Mccullers, J.A.; Saito, T.; Iverson, A.R. Multiple Genotypes of Influenza B Virus Circulated between 1979 and 2003. J. Virol. 2004, 78, 12817–12828. [Google Scholar] [CrossRef] [Green Version]
- Belshe, R.B. The need for quadrivalent vaccine against seasonal influenza. Vaccine 2010, 28, D45–D53. [Google Scholar] [CrossRef] [PubMed]
- Heikkinen, T.; Ikonen, N.; Ziegler, T. Impact of influenza B lineage-level mismatch between trivalent seasonal influenza vaccines and circulating viruses, 1999–2012. Clin. Infect. Dis. 2014, 59, 1519–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO Weekly epidemiological record: 2016, 88, 73–81. Available online: https://www.who.int/wer/2016/wer9107.pdf?ua=1 (accessed on 10 November 2020).
- Weir, J.P.; Gruber, M.F. An overview of the regulation of influenza vaccines in the United States. Influenza Other Respir. Viruses 2016, 10, 354–360. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.M.; Schild, G.C.; Newman, R.W.; Seagroatt, V. An improved single-radial-immunodiffusion technique for the assay of influenza haemagglutinin antigen: Application for potency determinations of inactivated whole virus and subunit vaccines. J. Biol. Stand. 1977, 5, 237–247. [Google Scholar] [CrossRef]
- Cate, T.R.; Couch, R.B.; Parker, D.; Baxter, B. Reactogenicity, immunogenicity, and antibody persistence in adults given inactivated influenza virus vaccines—1978. Rev. Infect. Dis. 1983, 5, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Hobson, D.; Curry, R.L.; Beare, A.S.; Ward-Gardner, A.; Refinery, E. The role of serum haemagglutination-inhibiting antibody in protection against challenge infection with influenza A2 and B viruses. J. Hyg. (Lond) 1972, 70, 767–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Montagne, J.R.; Noble, G.R.; Quinnan, G.V.; Curlin, G.T.; Blackwelder, W.C.; Smith, J.I.; Ennis, F.A.; Bozeman, F.M. Summary of clinical trials of inactivated influenza vaccine-1978. Rev. Infect. Dis. 1983, 5, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.S. Single-radial-immunodiffusion as an in vitro potency assay for human inactivated viral vaccines. Vet. Microbiol. 1993, 37, 253–262. [Google Scholar] [CrossRef]
- Williams, M.S.; Mayner, R.E.; Daniel, N.J.; Phelan, M.A.; Rastogi, S.C.; Bozeman, F.M.; Ennis, F.A. New developments in the measurement of the hemagglutinin content of influenza virus vaccines by single-radial-immunodiffusion. J. Biol. Stand. 1980, 8, 289–296. [Google Scholar] [CrossRef]
- Wise, T.G.; Dolin, R.; Mazur, M.H.; Top, F.H.; Edelman, R.; Ennis, F.A. Serologic responses and systemic reactions in adults after vaccination with bivalent A/victoria/75-A/new jersey/76 and monovalent B/hong kong/72 influenza vaccines. J. Infect. Dis. 1977, 136, S507–S517. [Google Scholar] [CrossRef]
- Wright, P.F.; Cherry, J.D.; Foy, H.M.; Glezen, W.P.; Hall, C.B.; McIntosh, K.; Monto, A.S.; Parrott, R.H.; Portnoy, B.; Taber, L.H. Antigenicity and reactogenicity of influenza A/USSR/77 virus vaccine in children--a multicentered evaluation of dosage and safety. Rev Infect Dis 1983, 5, 758–764. [Google Scholar] [CrossRef]
- Hirst, G.K. The agglutination of red cells by allantoic fluid of chick embryos infected with influenza virus. Science (80-) 1941, 94, 22–23. [Google Scholar] [CrossRef]
- Lambré, C.R.; Kasturi, K.N. A microplate immunoenzyme assay for anti-influenza antibodies. J. Immunol. Methods 1979, 26, 61–67. [Google Scholar] [CrossRef]
- Schild, G.C.; Pereira, M.S.; Chakraverty, P. Single-radial-hemolysis: A new method for the assay of antibody to influenza haemagglutinin. Applications for diagnosis and seroepidemiologic surveillance of influenza. Bull. World Health Organ. 1975, 52, 43–50. [Google Scholar]
- Bodle, J.; Verity, E.E.; Ong, C.; Vandenberg, K.; Shaw, R.; Barr, I.G.; Rockman, S. Development of an enzyme-linked immunoassay for the quantitation of influenza haemagglutinin: An alternative method to single radial immunodiffusion. Influenza Other Respir. Viruses 2013, 7, 191–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estmer Nilsson, C.; Abbas, S.; Bennemo, M.; Larsson, A.; Hämäläinen, M.D.; Frostell-Karlsson, Å. A novel assay for influenza virus quantification using surface plasmon resonance. Vaccine 2010, 28, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.M.; Gravel, C.; Farnsworth, A.; Zou, W.; Lemieux, M.; Xu, K.; Li, C.; Wang, J.; Goneau, M.F.; Merziotis, M.; et al. A Novel Synthetic Receptor-Based Immunoassay for Influenza Vaccine Quantification. PLoS ONE 2013, 8, e55428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khurana, S.; King, L.R.; Manischewitz, J.; Coyle, E.M.; Golding, H. Novel antibody-independent receptor-binding SPR-based assay for rapid measurement of influenza vaccine potency. Vaccine 2014, 32, 2188–2197. [Google Scholar] [CrossRef] [PubMed]
- Kuck, L.R.; Sorensen, M.; Matthews, E.; Srivastava, I.; Cox, M.M.J.; Rowlen, K.L. Titer on chip: New analytical tool for influenza vaccine potency determination. PLoS ONE 2014, 9, e109616. [Google Scholar] [CrossRef] [PubMed]
- Pierce, C.L.; Williams, T.L.; Moura, H.; Pirkle, J.L.; Cox, N.J.; Stevens, J.; Donis, R.O.; Barr, J.R. Quantification of immunoreactive viral influenza proteins by immunoaffinity capture and isotope-dilution liquid chromatography-tandem mass spectrometry. Anal. Chem. 2011, 83, 4729–4737. [Google Scholar] [CrossRef]
- Schmeisser, F.; Vasudevan, A.; Soto, J.; Kumar, A.; Williams, O.; Weir, J.P. A monoclonal antibody-based immunoassay for measuring the potency of 2009 pandemic influenza H1N1 vaccines. Influenza Other Respir. Viruses 2014, 8, 587–595. [Google Scholar] [CrossRef]
- Temperton, N.J.; Hoschler, K.; Major, D.; Nicolson, C.; Manvell, R.; Hien, V.M.; Ha, D.Q.; de Jong, M.D.; Zambon, M.C.; Takeuchi, Y.; et al. A sensitive retroviral pseudotype assay for influenza H5N1-neutralizing antibodies. Influenza Other Respir. Viruses 2007, 1, 105–112. [Google Scholar] [CrossRef]
- Trombetta, C.M.; Perini, D.; Mather, S.T.; Temperton, N.J.; Montomoli, E. Overview of Serological Techniques for Influenza Vaccine Evaluation: Past, Present and Future. Vaccines 2014, 2, 707–734. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.; Han, L.; Palladino, G.; Ferrari, A.; Xie, Y.; Carfi, A.; Dormitzer, P.R.; Settembre, E.C. Conformationally selective biophysical assay for influenza vaccine potency determination. Vaccine 2015, 33, 5342–5349. [Google Scholar] [CrossRef]
- Kendal, A.P.; Cate, T.R. Increased sensitivity and reduced specificity of hemagglutination inhibition tests with ether-treated influenza B/Singapore/222/79. J. Clin. Microbiol. 1983, 18, 930–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monto, A.S.; Maassab, H.F. Ether treatment of type B influenza virus antigen for the hemagglutination inhibition test. J. Clin. Microbiol. 1981, 13, 54–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyhälä, R.; Kleemola, M.; Visakorpi, R. The HI test modified by ether treatment in the sero-epidemiological surveillance of influenza B. J. Hyg. (Lond) 1985, 94, 341–348. [Google Scholar] [CrossRef] [Green Version]
- Chakraverty, P. Comparison of haemagglutination-inhibition and single-radial-haemolysis techniques for detection of antibodies to influenza B virus. Arch. Virol. 1980, 63, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Julkunen, I.; Pyhälä, R.; Hovi, T. Enzyme immunoassay, complement fixation and hemagglutination inhibition tests in the diagnosis of influenza A and B virus infections. Purified hemagglutinin in subtype-specific diagnosis. J. Virol. Methods 1985, 10, 75–84. [Google Scholar] [CrossRef]
- Oxford, J.S.; Yetts, R.; Schild, G.C. Quantitation and analysis of the specificity of post-immunization antibodies to influenza B viruses using single radial haemolysis. J. Hyg. 1982, 88, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Turner, R.; Lathey, J.L.; Van Voris, L.P.; Belshe, R.B. Serological diagnosis of influenza B virus infection: Comparison of an enzyme-linked immunosorbent assay and the hemagglutination inhibition test. J. Clin. Microbiol 1982, 15, 824–829. [Google Scholar] [CrossRef] [Green Version]
- Prevail, T.; Writing, I.I.; Ii, M.P.; Team, S.; Richard, T.; Davey, J.; Dodd, L.; Proschan, M.A.; Neaton, J.; Nordwall, J.N.; et al. A Randomized, Controlled Trial of ZMapp for Ebola Virus Infection. N. Engl. J. Med. 2016, 375, 1448–1456. [Google Scholar] [CrossRef]
- Qiu, X.; Wong, G.; Audet, J.; Bello, A.; Fernando, L.; Alimonti, J.B.; Fausther-Bovendo, H.; Wei, H.; Aviles, J.; Hiatt, E.; et al. Reversion of advanced Ebola virus disease in nonhuman primates with ZMapp. Nature 2014, 514, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Soto, J.; Vasudevan, A.; Schmeisser, F.; Alvarado-Facundo, E.; Wang, W.; Weiss, C.D.; Weir, J.P. Determination of influenza B identity and potency in quadrivalent inactivated influenza vaccines using lineage-specific monoclonal antibodies. PLoS ONE 2017, 12, e0175733. [Google Scholar] [CrossRef]
- Hai, R.; Krammer, F.; Tan, G.S.; Pica, N.; Eggink, D.; Maamary, J.; Margine, I.; Albrecht, R.A.; Palese, P. Influenza Viruses Expressing Chimeric Hemagglutinins: Globular Head and Stalk Domains Derived from Different Subtypes. J. Virol. 2012, 86, 5774–5781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Khayatt, R.; Jennings, R.; Potter, C.W. Interpretation of responses and protective levels of antibody against attenuated influenza A viruses using single radial haemolysis. J. Hyg. (Lond) 1984, 93, 301–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, R.J. Correlates of protection to influenza virus, where do we go from here? Hum. Vaccin. Immunother. 2013, 9, 405–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, C.W.; Oxford, J.S. Determinants of immunity to influenza infection in man. Br. Med. Bull. 1979, 35, 69–75. [Google Scholar] [CrossRef]
- Wood, J.M.; Gaines-Das, R.E.; Taylor, J.; Chakraverty, P. Comparison of influenza serological techniques by international collaborative study. Vaccine 1994, 12, 167–174. [Google Scholar] [CrossRef]
- Trombetta, C.M.; Remarque, E.J.; Mortier, D.; Montomoli, E. Comparison of hemagglutination inhibition, single radial hemolysis, virus neutralization assays, and ELISA to detect antibody levels against seasonal influenza viruses. Influenza Other Respir. Viruses 2018, 12, 675–686. [Google Scholar] [CrossRef]
- Demaison, C.; Parsley, K.; Brouns, G.; Scherr, M.; Battmer, K.; Kinnon, C.; Grez, M.; Thrasher, A.J. High-level transduction and gene expression in hematopoietic repopulating cells using a human immunodeficiency [correction of imunodeficiency] virus type 1-based lentiviral vector containing an internal spleen focus forming virus promoter. Hum. Gene Ther. 2002, 13, 803–813. [Google Scholar] [CrossRef]
- Zufferey, R.; Dull, T.; Mandel, R.J.; Bukovsky, A.; Quiroz, D.; Naldini, L.; Trono, D. Self-inactivating lentivirus vector for safe and efficient in vivo gene delivery. J. Virol. 1998, 72, 9873–9880. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.; Lee, K.P.; Park, S.J.; Park, J.H.; Jang, Y.S.; Choi, S.Y.; Jung, J.G.; Jo, K.; Park, D.Y.; Yoon, J.H.; et al. TMPRSS4 promotes invasion, migration and metastasis of human tumor cells by facilitating an epithelial-mesenchymal transition. Oncogene 2008, 27, 2635–2647. [Google Scholar] [CrossRef] [Green Version]
- Böttcher-Friebertshäuser, E.; Klenk, H.-D.; Garten, W. Activation of influenza viruses by proteases from host cells and bacteria in the human airway epithelium. Pathog. Dis. 2013, 69, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Böttcher-Friebertshäuser, E.; Matrosovich, T.Y.; Beyerle, M.; Klenk, H.-D.; Garten, W.; Matrosovich, M.N. Proteolytic activation of influenza viruses by serine proteases TMPRSS2 and HAT from human airway epithelium. J. Virol. 2006, 80, 9896–9898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böttcher-Friebertshäuser, E.; Freuer, C.; Steinmetzer, T.; Klenk, H.-D.; Garten, W. MDCK cells that express proteases TMPRSS2 and HAT provide a cell system to propagate influenza viruses in the absence of trypsin and to study cleavage of HA and its inhibition. Vaccine 2009, 27, 6324–6329. [Google Scholar] [CrossRef] [PubMed]
- Trombetta, C.M.; Perini, D.; Vitale, L.; Cox, R.J.; Stanzani, V.; Piccirella, S.; Montomoli, E. Validation of Single Radial Haemolysis assay: A reliable method to measure antibodies against influenza viruses. J. Immunol. Methods 2015, 422, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, F.; Carnell, G.; Kinsley, R.; Böttcher-Friebertshäuser, E.; Pöhlmann, S.; Scott, S.; Fereidouni, S.; Corti, D.; Kellam, P.; Gilbert, S.C.; et al. Development and use of lentiviral vectors pseudotyped with influenza B haemagglutinins: Application to vaccine immunogenicity, mAb potency and sero-surveillance studies. BioRxiv 2018. [Google Scholar] [CrossRef]
- Robertson, J.S.; Naeve, C.W.; Webster, R.G.; Bootman, J.S.; Newman, R.W.; Schild, G.C. Alterations in the hemagglutinin associated with adaptation of influenza B virus to growth in eggs. Virology 1985, 143, 166–174. [Google Scholar] [CrossRef]
- Shaw, M.W.; Xu, X.; Li, Y.; Normand, S.; Ueki, R.T.; Kunimoto, G.Y.; Hall, H.; Klimov, A.; Cox, N.J.; Subbarao, K. Reappearance and Global Spread of Variants of Influenza B/Victoria/2/87 Lineage Viruses in the 2000–2001 and 2001–2002 Seasons. Virology 2002. [Google Scholar] [CrossRef] [Green Version]
- Saito, T.; Nakaya, Y.; Suzuki, T.; Ito, R.; Saito, T.; Saito, H.; Takao, S.; Sahara, K.; Odagiri, T.; Murata, T.; et al. Antigenic alteration of influenza B virus associated with loss of a glycosylation site due to host-cell adaptation. J. Med. Virol. 2004, 74, 336–343. [Google Scholar] [CrossRef]
- Ni, F.; Kondrashkina, E.; Wang, Q. Structural basis for the divergent evolution of influenza B virus hemagglutinin. Virology 2013, 446, 112–122. [Google Scholar] [CrossRef] [Green Version]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Accession | Lineage | Protease | ng per 6-Well |
|---|---|---|---|---|
| B/Hong Kong/8/1973 | K00425 | Pre-split | HAT | 125 |
| B/Victoria/2/1987 | FJ766840 | Victoria | HAT | 125 |
| B/Brisbane/60/2008 | KX058884 | Victoria | TMPRSS4 | 125 |
| B/Yamagata/16/1988 | CY018765 | Yamagata | HAT | 125 |
| B/Florida/4/2006 | EU515992 | Yamagata | HAT | 125 |
| B/Bangladesh/3333/2007 | CY115255 | Yamagata | HAT | 250 |
| mAb | 2F11 | 3E8 | 1H4 | 8E12 | 5A1 |
|---|---|---|---|---|---|
| Target lineage | Cross | Yamagata | Yamagata | Victoria | Victoria |
| mAb | Target | B/Hong Kong/8/1973 | B/Florida/4/2006 | B/Bangladesh/3333/2007 | B/Victoria/2/1987 | B/Brisbane/60/2008 |
|---|---|---|---|---|---|---|
| 2F11 | Cross | 8 | 3 | 2 | 4 | - |
| 3E8 | Yamagata | - | 34 | 21 | 3 | - |
| 1H4 | Yamagata | - | 6 | 3 | 4 | - |
| 8E12 | Victoria | 2 | - | - | - | - |
| 5A1 | VIctoria | 4 | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carnell, G.W.; Trombetta, C.M.; Ferrara, F.; Montomoli, E.; Temperton, N.J. Correlation of Influenza B Haemagglutination Inhibiton, Single-Radial Haemolysis and Pseudotype-Based Microneutralisation Assays for Immunogenicity Testing of Seasonal Vaccines. Vaccines 2021, 9, 100. https://doi.org/10.3390/vaccines9020100
Carnell GW, Trombetta CM, Ferrara F, Montomoli E, Temperton NJ. Correlation of Influenza B Haemagglutination Inhibiton, Single-Radial Haemolysis and Pseudotype-Based Microneutralisation Assays for Immunogenicity Testing of Seasonal Vaccines. Vaccines. 2021; 9(2):100. https://doi.org/10.3390/vaccines9020100
Chicago/Turabian StyleCarnell, George W., Claudia M. Trombetta, Francesca Ferrara, Emanuele Montomoli, and Nigel J. Temperton. 2021. "Correlation of Influenza B Haemagglutination Inhibiton, Single-Radial Haemolysis and Pseudotype-Based Microneutralisation Assays for Immunogenicity Testing of Seasonal Vaccines" Vaccines 9, no. 2: 100. https://doi.org/10.3390/vaccines9020100
APA StyleCarnell, G. W., Trombetta, C. M., Ferrara, F., Montomoli, E., & Temperton, N. J. (2021). Correlation of Influenza B Haemagglutination Inhibiton, Single-Radial Haemolysis and Pseudotype-Based Microneutralisation Assays for Immunogenicity Testing of Seasonal Vaccines. Vaccines, 9(2), 100. https://doi.org/10.3390/vaccines9020100