
Vaccination with LytA, LytC, or Pce of Streptococcus pneumoniae Protects against Sepsis by Inducing IgGs That Activate the Complement System


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Immunization Experiments in Mice with LytA, LytC, or Pce
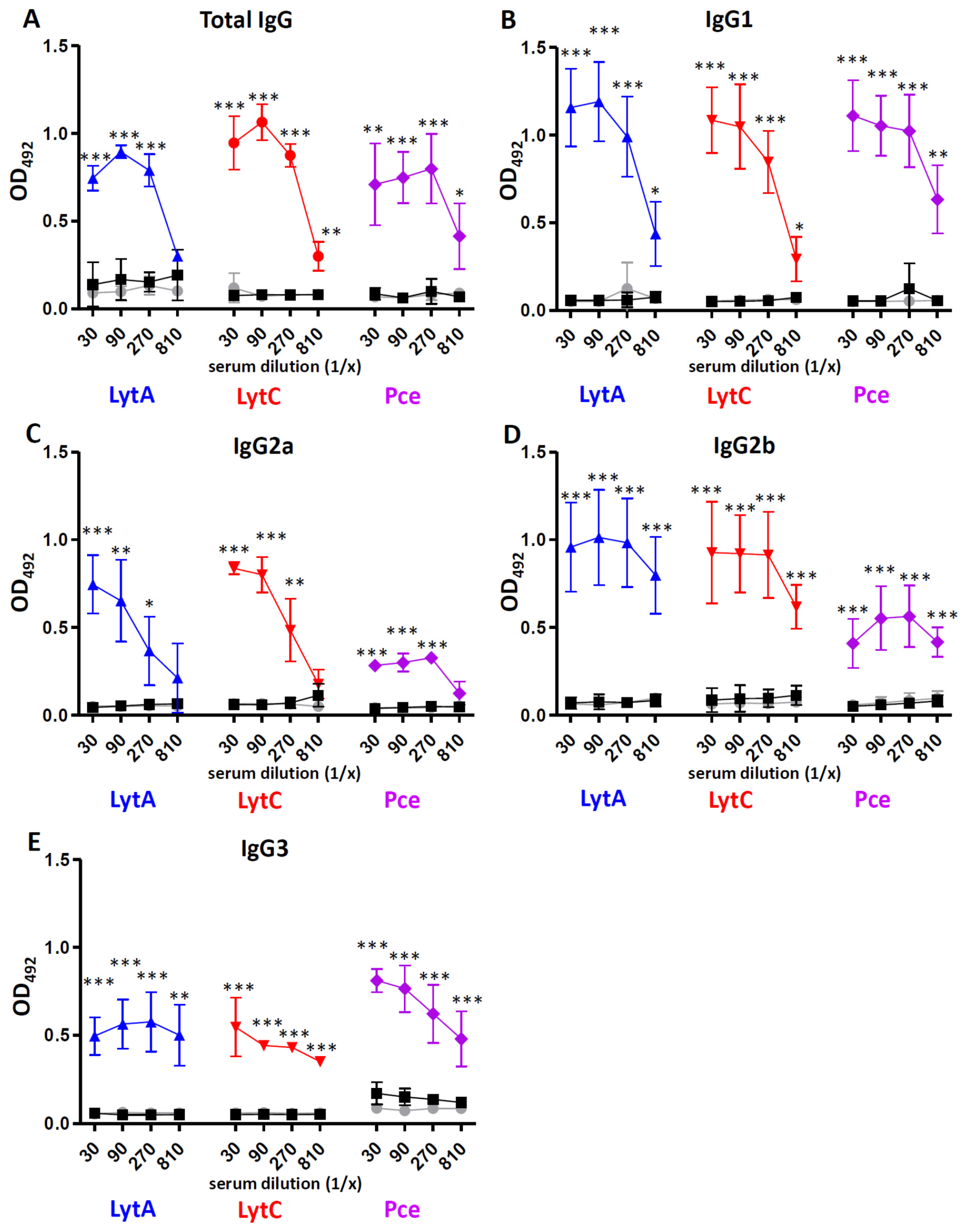
2.3. Enzyme-Linked Immunosorbent Assays to Detect Ig Subclasses
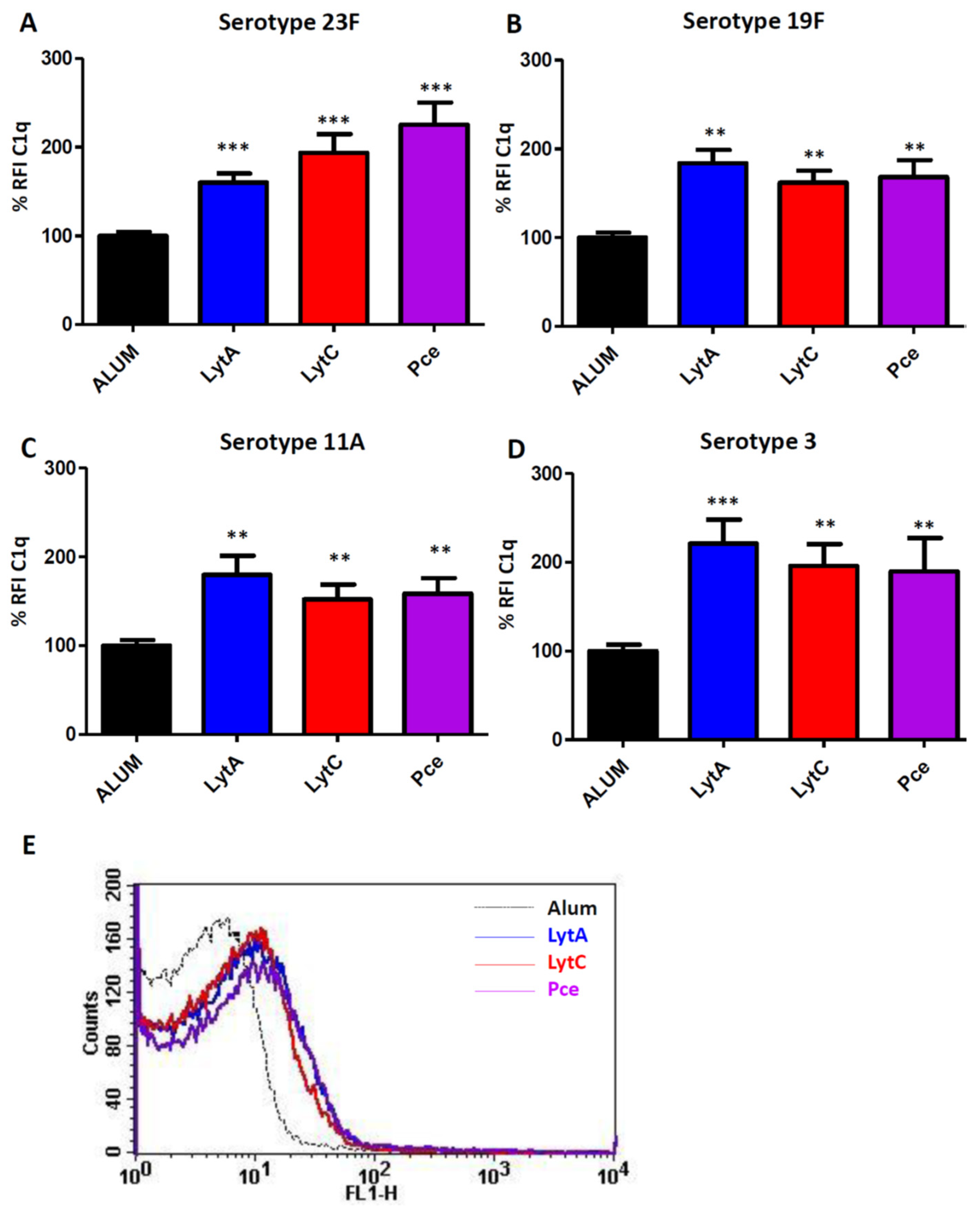
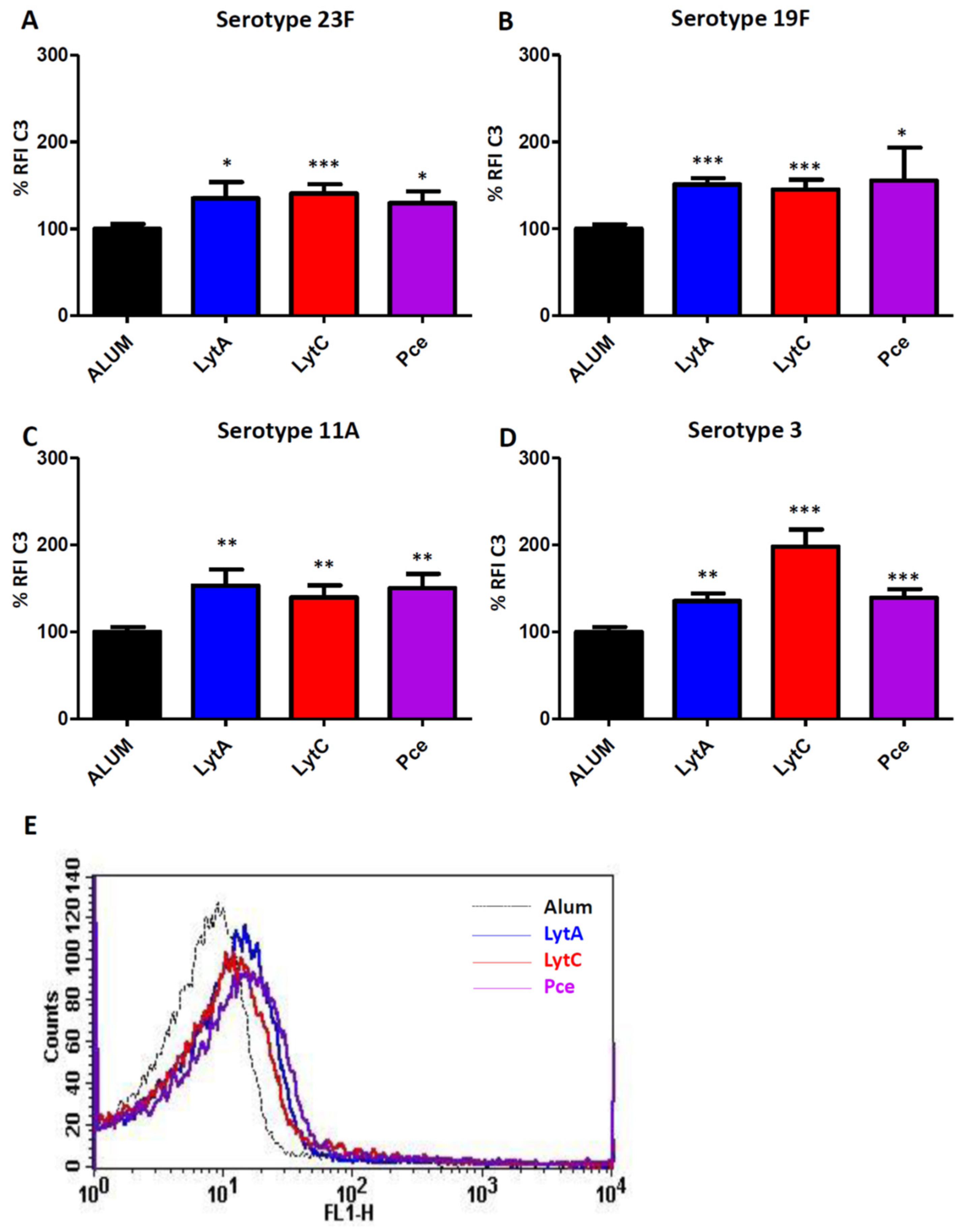
2.4. Activation of Complement-Mediated Immunity by Specific IgGs
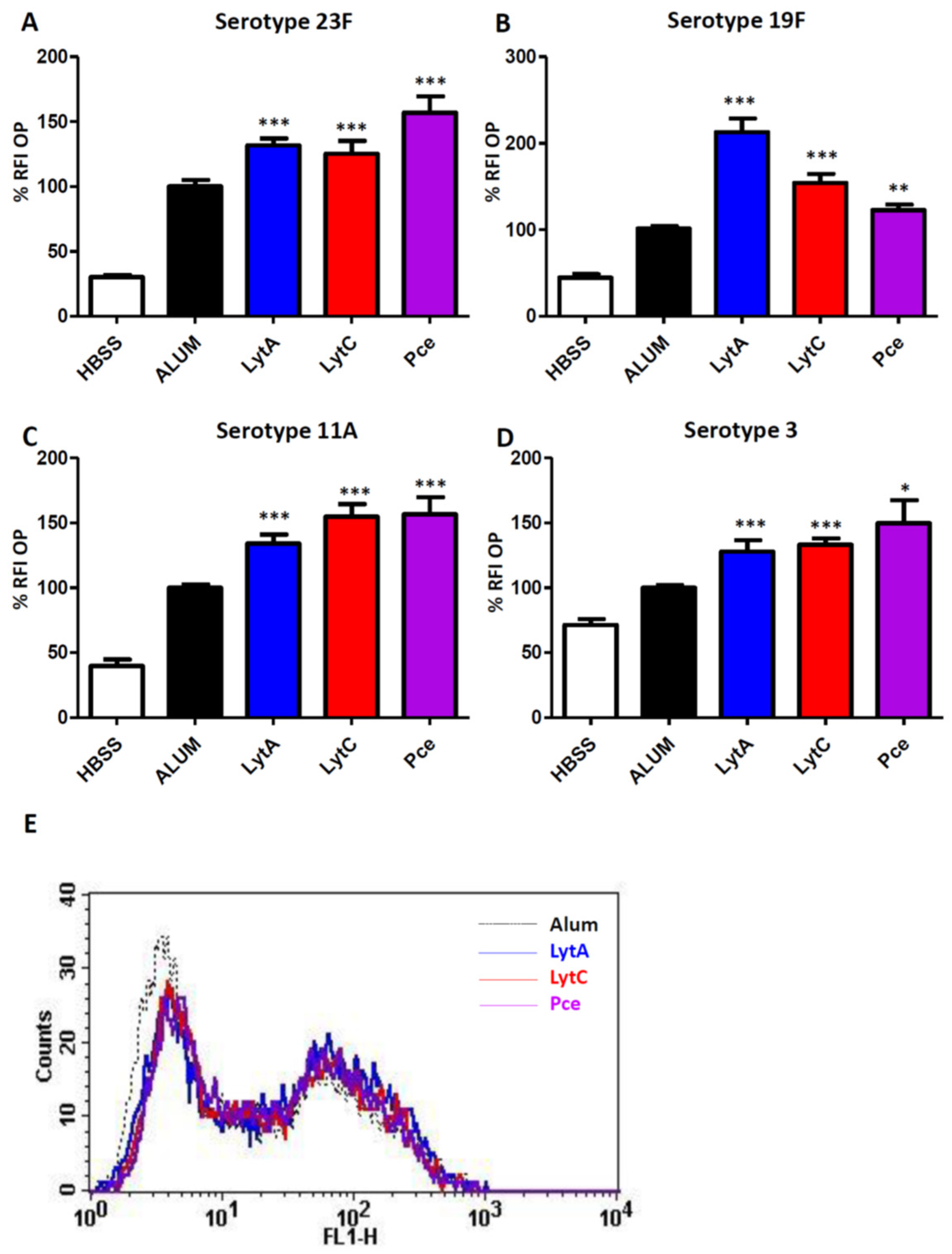
2.5. Opsonophagocytosis (OP) Using HL-60 Cells
2.6. Statistical Analysis
3. Results
3.1. Antibody Response to Cell Wall Hydrolases LytA, LytC, and Pce
3.2. Complement Activation Mediated by Antibodies to LytA, LytC, or Pce
3.3. Immunization with LytA, LytC, or Pce Induces Activation of the Opsonophagocytosis Process of S. pneumoniae Regardless of the Serotype
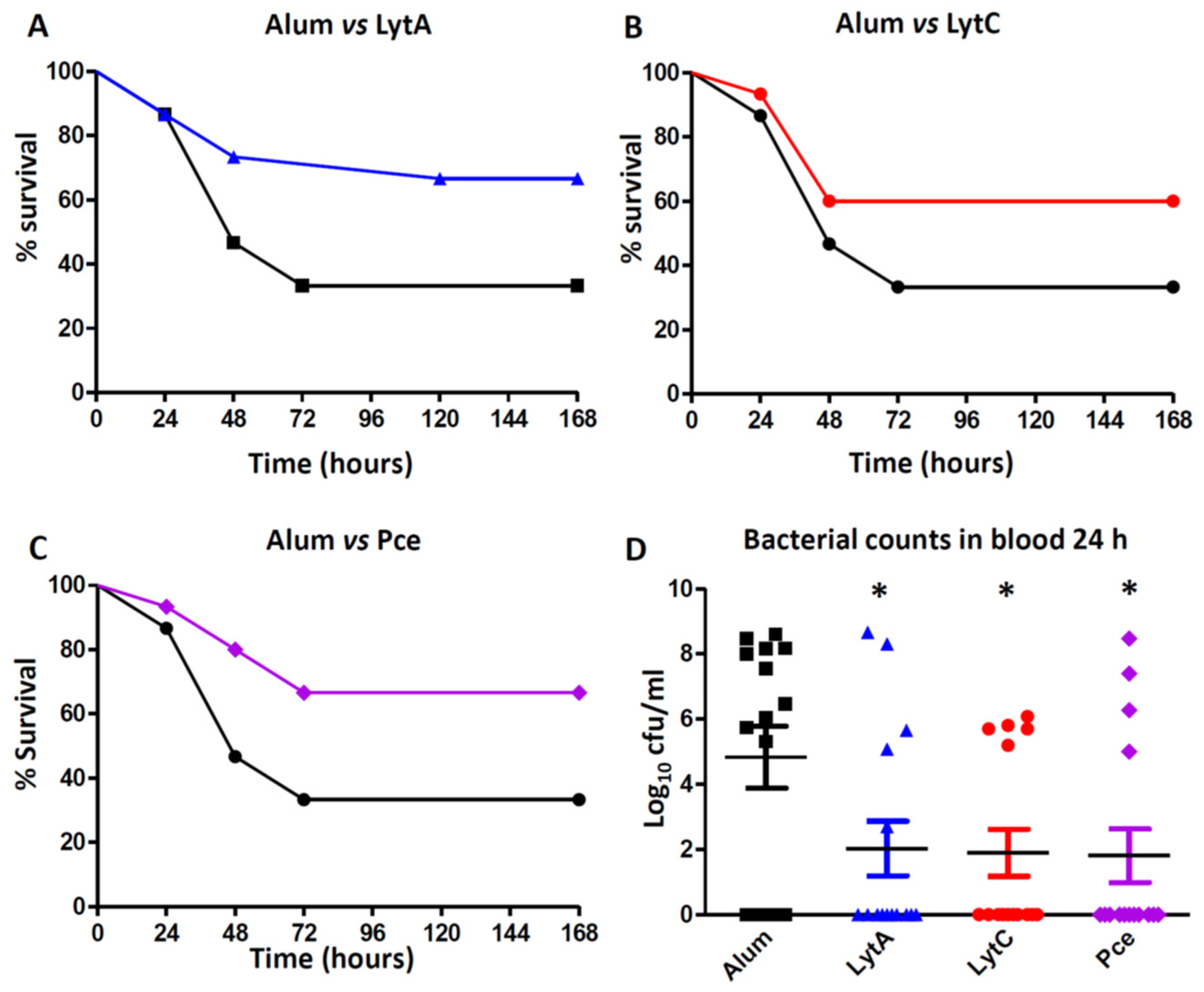
3.4. Vaccination with LytA, LytC, or Pce Protects Mice against Systemic Infection by S. Pneumoniae
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2016 Lower Respiratory Infections Collaborators. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of lower respiratory infections in 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1191–1210. [Google Scholar] [CrossRef] [Green Version]
- Kyaw, M.H.; Lynfield, R.; Schaffner, W.; Craig, A.S.; Hadler, J.; Reingold, A.; Thomas, A.R.; Harrison, L.H.; Bennett, N.M.; Farley, M.M.; et al. Effect of introduction of the pneumococcal conjugate vaccine on drug-resistant Streptococcus pneumoniae. N. Engl. J. Med. 2006, 354, 1455–1463. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, J.M.; Jiang, Q.; Isturiz, R.E.; Sings, H.L.; Swerdlow, D.L.; Gessner, B.D.; Carrico, R.M.; Peyrani, P.; Wiemken, T.L.; Mattingly, W.A.; et al. Effectiveness of 13-Valent Pneumococcal Conjugate Vaccine Against Hospitalization for Community-Acquired Pneumonia in Older US Adults: A Test-Negative Design. Clin. Infect. Dis. 2018, 67, 1498–1506. [Google Scholar] [CrossRef] [PubMed]
- Whitney, C.G.; Farley, M.M.; Hadler, J.; Harrison, L.H.; Bennett, N.M.; Lynfield, R.; Reingold, A.; Cieslak, P.R.; Pilishvili, T.; Jackson, D.; et al. Decline in invasive pneumococcal disease after the introduction of protein-polysaccharide conjugate vaccine. N. Engl. J. Med. 2003, 348, 1737–1746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regev-Yochay, G.; Katzir, M.; Strahilevitz, J.; Rahav, G.; Finn, T.; Miron, D.; Maor, Y.; Chazan, B.; Schindler, Y.; Dagan, R.; et al. The herd effects of infant PCV7/PCV13 sequential implementation on adult invasive pneumococcal disease, six years post implementation; a nationwide study in Israel. Vaccine 2017, 35, 2449–2456. [Google Scholar] [CrossRef] [PubMed]
- Jayasinghe, S.; Menzies, R.; Chiu, C.; Toms, C.; Blyth, C.C.; Krause, V.; McIntyre, P. Long-term impact of a “3 + 0” schedule for 7- and 13-valent pneumococcal conjugate vaccines on invasive pneumococcal disease in Australia, 2002–2014. Clin. Infect. Dis. 2017, 64, 175–183. [Google Scholar] [CrossRef] [Green Version]
- Fenoll, A.; Granizo, J.J.; Gimenez, M.J.; Yuste, J.; Aguilar, L. Secular trends (1990–2013) in serotypes and associated non-susceptibility of S. pneumoniae isolates causing invasive disease in the pre-/post-era of pneumococcal conjugate vaccines in Spanish regions without universal paediatric pneumococcal vaccination. Vaccine 2015, 33, 5691–5699. [Google Scholar] [CrossRef] [PubMed]
- Falkenhorst, G.; Remschmidt, C.; Harder, T.; Hummers-Pradier, E.; Wichmann, O.; Bogdan, C. Effectiveness of the 23-valent pneumococcal polysaccharide vaccine (PPV23) against pneumococcal disease in the elderly: Systematic review and meta-analysis. PLoS ONE 2017, 12, e0169368. [Google Scholar] [CrossRef]
- Lawrence, H.; Pick, H.; Baskaran, V.; Daniel, P.; Rodrigo, C.; Ashton, D.; Edwards-Pritchard, R.C.; Sheppard, C.; Eletu, S.D.; Litt, D.; et al. Effectiveness of the 23-valent pneumococcal polysaccharide vaccine against vaccine serotype pneumococcal pneumonia in adults: A case-control test-negative design study. PLoS Med. 2020, 17, e1003326. [Google Scholar] [CrossRef] [PubMed]
- Bonten, M.J.; Huijts, S.M.; Bolkenbaas, M.; Webber, C.; Patterson, S.; Gault, S.; van Werkhoven, C.H.; van Deursen, A.M.; Sanders, E.A.; Verheij, T.J.; et al. Polysaccharide conjugate vaccine against pneumococcal pneumonia in adults. N. Engl. J. Med. 2015, 372, 1114–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaughlin, J.M.; Jiang, Q.; Gessner, B.D.; Swerdlow, D.L.; Sings, H.L.; Isturiz, R.E.; Jodar, L. Pneumococcal conjugate vaccine against serotype 3 pneumococcal pneumonia in adults: A systematic review and pooled analysis. Vaccine 2019, 37, 6310–6316. [Google Scholar] [CrossRef]
- de Miguel, S.; Domenech, M.; González-Camacho, F.; Sempere, J.; Vicioso, D.; Sanz, J.C.; García Comas, L.; Ardanuy, C.; Fenoll, A.; Yuste, J. Nationwide trends of invasive pneumococcal disease in Spain (2009–2019) in children and adults during the pneumococcal conjugate vaccine era. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Ganaie, F.; Saad, J.S.; McGee, L.; van Tonder, A.J.; Bentley, S.D.; Lo, S.W.; Gladstone, R.A.; Turner, P.; Keenan, J.D.; Breiman, R.F.; et al. A New Pneumococcal capsule type, 10D, is the 100th serotype and has a large cps fragment from an oral Streptococcus. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Ladhani, S.N.; Collins, S.; Djennad, A.; Sheppard, C.L.; Borrow, R.; Fry, N.K.; Andrews, N.J.; Miller, E.; Ramsay, M.E. Rapid increase in non-vaccine serotypes causing invasive pneumococcal disease in England and Wales, 2000–2017: A prospective national observational cohort study. Lancet Infect. Dis. 2018, 18, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Cassiolato, A.P.; Almeida, S.C.G.; Andrade, A.L.; Minamisava, R.; Brandileone, M.C.C. Expansion of the multidrug-resistant clonal complex 320 among invasive Streptococcus pneumoniae serotype 19A after the introduction of a ten-valent pneumococcal conjugate vaccine in Brazil. PLoS ONE 2018, 13, e0208211. [Google Scholar] [CrossRef] [Green Version]
- Savulescu, C.; Krizova, P.; Lepoutre, A.; Mereckiene, J.; Vestrheim, D.F.; Ciruela, P.; Ordobas, M.; Guevara, M.; McDonald, E.; Morfeldt, E.; et al. Effect of high-valency pneumococcal conjugate vaccines on invasive pneumococcal disease in children in SpIDnet countries: An observational multicentre study. Lancet Respir. Med. 2017, 5, 648–656. [Google Scholar] [CrossRef] [Green Version]
- Ouldali, N.; Varon, E.; Levy, C.; Angoulvant, F.; Georges, S.; Ploy, M.C.; Kempf, M.; Cremniter, J.; Cohen, R.; Bruhl, D.L.; et al. Invasive pneumococcal disease incidence in children and adults in France during the pneumococcal conjugate vaccine era: An interrupted time-series analysis of data from a 17-year national prospective surveillance study. Lancet Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Sempere, J.; de Miguel, S.; González-Camacho, F.; Yuste, J.; Domenech, M. Clinical relevance and molecular pathogenesis of the emerging serotypes 22F and 33F of Streptococcus pneumoniae in Spain. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Feldman, C.; Anderson, R. Review: Current and new generation pneumococcal vaccines. J. Infect. 2014, 69, 309–325. [Google Scholar] [CrossRef] [Green Version]
- Giefing, C.; Meinke, A.L.; Hanner, M.; Henics, T.; Bui, M.D.; Gelbmann, D.; Lundberg, U.; Senn, B.M.; Schunn, M.; Habel, A.; et al. Discovery of a novel class of highly conserved vaccine antigens using genomic scale antigenic fingerprinting of pneumococcus with human antibodies. J. Exp. Med. 2008, 205, 117–131. [Google Scholar] [CrossRef]
- Chan, W.Y.; Entwisle, C.; Ercoli, G.; Ramos-Sevillano, E.; McIlgorm, A.; Cecchini, P.; Bailey, C.; Lam, O.; Whiting, G.; Green, N.; et al. A Novel, Multiple-Antigen Pneumococcal Vaccine Protects against Lethal Streptococcus pneumoniae Challenge. Infect. Immun. 2019, 87. [Google Scholar] [CrossRef] [Green Version]
- Converso, T.R.; Goulart, C.; Darrieux, M.; Leite, L.C.C. A protein chimera including PspA in fusion with PotD is protective against invasive pneumococcal infection and reduces nasopharyngeal colonization in mice. Vaccine 2017, 35, 5140–5147. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.S.; Ogunniyi, A.D.; Woodrow, M.C.; Holden, D.W.; Paton, J.C. Immunization with components of two iron uptake ABC transporters protects mice against systemic Streptococcus pneumoniae infection. Infect. Immun. 2001, 69, 6702–6706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogunniyi, A.D.; Folland, R.L.; Briles, D.E.; Hollingshead, S.K.; Paton, J.C. Immunization of mice with combinations of pneumococcal virulence proteins elicits enhanced protection against challenge with Streptococcus pneumoniae. Infect. Immun. 2000, 68, 3028–3033. [Google Scholar] [CrossRef] [Green Version]
- Ogunniyi, A.D.; Woodrow, M.C.; Poolman, J.T.; Paton, J.C. Protection against Streptococcus pneumoniae elicited by immunization with pneumolysin and CbpA. Infect. Immun. 2001, 69, 5997–6003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masomian, M.; Ahmad, Z.; Gew, L.T.; Poh, C.L. Development of next generation Streptococcus pneumoniae vaccines conferring broad protection. Vaccines 2020, 8, 132. [Google Scholar] [CrossRef] [Green Version]
- Corsini, B.; Aguinagalde, L.; Ruiz, S.; Domenech, M.; Antequera, M.L.; Fenoll, A.; García, P.; García, E.; Yuste, J. Immunization with LytB protein of Streptococcus pneumoniae activates complement-mediated phagocytosis and induces protection against pneumonia and sepsis. Vaccine 2016, 34, 6148–6157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lock, R.A.; Hansman, D.; Paton, J.C. Comparative efficacy of autolysin and pneumolysin as immunogens protecting mice against infection by Streptococcus pneumoniae. Microb. Pathog. 1992, 12, 137–143. [Google Scholar] [CrossRef]
- Canvin, J.R.; Marvin, A.P.; Sivakumaran, M.; Paton, J.C.; Boulnois, G.J.; Andrew, P.W.; Mitchell, T.J. The role of pneumolysin and autolysin in the pathology of pneumonia and septicemia in mice infected with a type 2 pneumococcus. J. Infect. Dis. 1995, 172, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Sevillano, E.; Urzainqui, A.; Campuzano, S.; Moscoso, M.; González-Camacho, F.; Domenech, M.; Rodríguez de Córdoba, S.; Saánchez-Madrid, F.; Brown, J.S.; García, E.; et al. Pleiotropic effects of cell wall amidase LytA on Streptococcus pneumoniae sensitivity to the host immune response. Infect. Immun. 2015, 83, 591–603. [Google Scholar] [CrossRef] [Green Version]
- Eldholm, V.; Johnsborg, O.; Haugen, K.; Ohnstad, H.S.; Håvarstein, L.S. Fratricide in Streptococcus pneumoniae: Contributions and role of the cell wall hydrolases CbpD, LytA and LytC. Microbiology 2009, 155, 2223–2234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moscoso, M.; García, E.; López, R. Biofilm formation by Streptococcus pneumoniae: Role of choline, extracellular DNA, and capsular polysaccharide in microbial accretion. J. Bacteriol. 2006, 188, 7785–7795. [Google Scholar] [CrossRef] [Green Version]
- Gosink, K.K.; Mann, E.R.; Guglielmo, C.; Tuomanen, E.I.; Masure, H.R. Role of novel choline binding proteins in virulence of Streptococcus pneumoniae. Infect. Immun. 2000, 68, 5690–5695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Sevillano, E.; Moscoso, M.; García, P.; García, E.; Yuste, J. Nasopharyngeal colonization and invasive disease are enhanced by the cell wall hydrolases LytB and LytC of Streptococcus pneumoniae. PLoS ONE 2011, 6, e23626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermoso, J.A.; Lagartera, L.; González, A.; Stelter, M.; García, P.; Martínez-Ripoll, M.; García, J.L.; Menéndez, M. Insights into pneumococcal pathogenesis from the crystal structure of the modular teichoic acid phosphorylcholine esterase Pce. Nat. Struct. Mol. Biol. 2005, 12, 533–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, J.L.; García, E.; López, R. Overproduction and rapid purification of the amidase of Streptococcus pneumoniae. Arch. Microbiol. 1987, 149, 52–56. [Google Scholar] [CrossRef]
- García, P.; Paz González, M.; García, E.; García, J.L.; López, R. The molecular characterization of the first autolytic lysozyme of Streptococcus pneumoniae reveals evolutionary mobile domains. Mol. Microbiol. 1999, 33, 128–138. [Google Scholar] [CrossRef]
- Sánchez-Puelles, J.M.; Sanz, J.M.; García, J.L.; García, E. Cloning and expression of gene fragments encoding the choline-binding domain of pneumococcal murein hydrolases. Gene 1990, 89, 69–75. [Google Scholar] [CrossRef]
- Lagartera, L.; González, A.; Stelter, M.; García, P.; Kahn, R.; Menéndez, M.; Hermoso, J.A. Crystallization and preliminary X-ray diffraction studies of the pneumococcal teichoic acid phosphorylcholine esterase Pce. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2005, 61, 221–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jomaa, M.; Yuste, J.; Paton, J.C.; Jones, C.; Dougan, G.; Brown, J.S. Antibodies to the iron uptake ABC transporter lipoproteins PiaA and PiuA promote opsonophagocytosis of Streptococcus pneumoniae. Infect. Immun. 2005, 73, 6852–6859. [Google Scholar] [CrossRef] [Green Version]
- Cafini, F.; Yuste, J.; Gimenez, M.J.; Sevillano, D.; Aguilar, L.; Alou, L.; Ramos-Sevillano, E.; Torrico, M.; Gonzalez, N.; Garcia, E.; et al. Enhanced in vivo activity of cefditoren in pre-immunized mice against penicillin-resistant S. pneumoniae (serotypes 6B, 19F and 23F) in a sepsis model. PLoS ONE 2010, 5, e12041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuste, J.; Sen, A.; Truedsson, L.; Jönsson, G.; Tay, L.S.; Hyams, C.; Baxendale, H.E.; Goldblatt, F.; Botto, M.; Brown, J.S. Impaired opsonization with C3b and phagocytosis of Streptococcus pneumoniae in sera from subjects with defects in the classical complement pathway. Infect. Immun. 2008, 76, 3761–3770. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Sevillano, E.; Rodríguez-Sosa, C.; Cafini, F.; Giménez, M.J.; Navarro, A.; Sevillano, D.; Alou, L.; García, E.; Aguilar, L.; Yuste, J. Cefditoren and ceftriaxone enhance complement-mediated immunity in the presence of specific antibodies against antibiotic-resistant pneumococcal strains. PLoS ONE 2012, 7, e44135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Sevillano, E.; Rodríguez-Sosa, C.; Díez-Martínez, R.; Giménez, M.J.; Olmedillas, E.; García, P.; García, E.; Aguilar, L.; Yuste, J. Macrolides and beta-lactam antibiotics enhance C3b deposition on the surface of multidrug-resistant Streptococcus pneumoniae strains by a LytA autolysin-dependent mechanism. Antimicrob. Agents Chemother. 2012, 56, 5534–5540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Exley, R.M.; Shaw, J.; Mowe, E.; Sun, Y.H.; West, N.P.; Williamson, M.; Botto, M.; Smith, H.; Tang, C.M. Available carbon source influences the resistance of Neisseria meningitidis against complement. J. Exp. Med. 2005, 201, 1637–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Steiner, S.; Libutti, D.; Pais, L.B.; Dykes, J.; Anderson, P.; Whitin, J.C.; Keyserling, H.L.; Carlone, G.M. Standardization of an opsonophagocytic assay for the measurement of functional antibody activity against Streptococcus pneumoniae using differentiated HL-60 cells. Clin. Diagn. Lab. Immunol. 1997, 4, 415–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Sevillano, E.; Urzainqui, A.; de Andrés, B.; González-Tajuelo, R.; Domenech, M.; González-Camacho, F.; Sánchez-Madrid, F.; Brown, J.S.; García, E.; Yuste, J. PSGL-1 on leukocytes is a critical component of the host immune response against invasive pneumococcal disease. PLoS Pathog. 2016, 12, e1005500. [Google Scholar] [CrossRef] [PubMed]
- López, R.; García, E. Recent trends on the molecular biology of pneumococcal capsules, lytic enzymes, and bacteriophage. FEMS Microbiol. Rev. 2004, 28, 553–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, M.; García, P.; de la Campa, A.G.; Linares, J.; Ardanuy, C.; García, E. Evidence of localized prophage-host recombination in the lytA gene, encoding the major pneumococcal autolysin. J. Bacteriol. 2010, 192, 2624–2632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, F.; Talekar, S.J.; Lanata, C.F.; Grijalva, C.G.; Klugman, K.P.; Vidal, J.E.; Group, R.P.; Investigators, G. Expression of Streptococcus pneumoniae virulence-related genes in the nasopharynx of healthy children. PLoS ONE 2013, 8, e67147. [Google Scholar] [CrossRef] [Green Version]
- Desa, M.N.; Sekaran, S.D.; Vadivelu, J.; Parasakthi, N. Distribution of CBP genes in Streptococcus pneumoniae isolates in relation to vaccine types, penicillin susceptibility and clinical site. Epidemiol. Infect. 2008, 136, 940–942. [Google Scholar] [CrossRef]
- Bologa, M.; Kamtchoua, T.; Hopfer, R.; Sheng, X.; Hicks, B.; Bixler, G.; Hou, V.; Pehlic, V.; Yuan, T.; Gurunathan, S. Safety and immunogenicity of pneumococcal protein vaccine candidates: Monovalent choline-binding protein A (PcpA) vaccine and bivalent PcpA-pneumococcal histidine triad protein D vaccine. Vaccine 2012, 30, 7461–7468. [Google Scholar] [CrossRef]
- Kamtchoua, T.; Bologa, M.; Hopfer, R.; Neveu, D.; Hu, B.; Sheng, X.; Corde, N.; Pouzet, C.; Zimmermann, G.; Gurunathan, S. Safety and immunogenicity of the pneumococcal pneumolysin derivative PlyD1 in a single-antigen protein vaccine candidate in adults. Vaccine 2013, 31, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Uddin, S.; Borrow, R.; Haeney, M.R.; Moran, A.; Warrington, R.; Balmer, P.; Arkwright, P.D. Total and serotype-specific pneumococcal antibody titres in children with normal and abnormal humoral immunity. Vaccine 2006, 24, 5637–5644. [Google Scholar] [CrossRef] [PubMed]
- Lottenbach, K.R.; Mink, C.M.; Barenkamp, S.J.; Anderson, E.L.; Homan, S.M.; Powers, D.C. Age-associated differences in immunoglobulin G1 (IgG1) and IgG2 subclass antibodies to pneumococcal polysaccharides following vaccination. Infect. Immun. 1999, 67, 4935–4938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelly, M.A.; Jacoby, H.; Riley, G.J.; Graves, B.T.; Pichichero, M.; Treanor, J.J. Comparison of pneumococcal polysaccharide and CRM197-conjugated pneumococcal oligosaccharide vaccines in young and elderly adults. Infect. Immun. 1997, 65, 242–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuorimaa, T.; Kayhty, H.; Leroy, O.; Eskola, J. Tolerability and immunogenicity of an 11-valent pneumococcal conjugate vaccine in adults. Vaccine 2001, 19, 1863–1869. [Google Scholar] [CrossRef]
- Soininen, A.; Seppala, I.; Nieminen, T.; Eskola, J.; Kayhty, H. IgG subclass distribution of antibodies after vaccination of adults with pneumococcal conjugate vaccines. Vaccine 1999, 17, 1889–1897. [Google Scholar] [CrossRef]
- Amir, J.; Scott, M.G.; Nahm, M.H.; Granoff, D.M. Bactericidal and opsonic activity of IgG1 and IgG2 anticapsular antibodies to Haemophilus influenzae type b. J. Infect. Dis. 1990, 162, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Bredius, R.G.; Driedijk, P.C.; Schouten, M.F.; Weening, R.S.; Out, T.A. Complement activation by polyclonal immunoglobulin G1 and G2 antibodies against Staphylococcus aureus, Haemophilus influenzae type b, and tetanus toxoid. Infect. Immun. 1992, 60, 4838–4847. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.S.; Hussell, T.; Gilliland, S.M.; Holden, D.W.; Paton, J.C.; Ehrenstein, M.R.; Walport, M.J.; Botto, M. The classical pathway is the dominant complement pathway required for innate immunity to Streptococcus pneumoniae infection in mice. Proc. Natl. Acad. Sci. USA 2002, 99, 16969–16974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, J.G.; Kim, H.W.; Choi, H.J.; Lee, J.H.; Kim, K.H. Functional immune responses to twelve serotypes after immunization with a 23-valent pneumococcal polysaccharide vaccine in older adults. Vaccine 2015, 33, 4770–4775. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Pichichero, M. Comparison of anti-capsular antibody quantity and functionality in children after different primary dose and booster schedules of 13 valent-pneumococcal conjugate vaccine. Vaccine 2020, 38, 4423–4431. [Google Scholar] [CrossRef] [PubMed]
- Standish, A.J.; Weiser, J.N. Human neutrophils kill Streptococcus pneumoniae via serine proteases. J. Immunol. 2009, 183, 2602–2609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, P.J.; Sternby, E.; Ursing, B. Septicemia in granulocytopenic patients: A shift in bacterial etiology. Scand. J. Infect. Dis. 1992, 24, 357–360. [Google Scholar] [CrossRef]
- Briles, D.E.; Claflin, J.L.; Schroer, K.; Forman, C. Mouse Igg3 antibodies are highly protective against infection with Streptococcus pneumoniae. Nature 1981, 294, 88–90. [Google Scholar] [CrossRef]
- Wizemann, T.M.; Heinrichs, J.H.; Adamou, J.E.; Erwin, A.L.; Kunsch, C.; Choi, G.H.; Barash, S.C.; Rosen, C.A.; Masure, H.R.; Tuomanen, E.; et al. Use of a whole genome approach to identify vaccine molecules affording protection against Streptococcus pneumoniae infection. Infect. Immun. 2001, 69, 1593–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.Q.; Lv, Z.Y.; Gan, H.Q.; Xian, M.; Zhang, K.X.; Mai, J.Y.; Yu, X.B.; Wu, Z.D. Intranasal immunization with autolysin (LytA) in mice model induced protection against five prevalent Streptococcus pneumoniae serotypes in China. Immunol. Res. 2011, 51, 108–115. [Google Scholar] [CrossRef]
- Lipsitch, M.; Siber, G.R. How Can Vaccines Contribute to Solving the Antimicrobial Resistance Problem? mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosini, R.; Nicchi, S.; Pizza, M.; Rappuoli, R. Vaccines Against Antimicrobial Resistance. Front. Immunol. 2020, 11, 1048. [Google Scholar] [CrossRef] [PubMed]
- Domenech, M.; Sempere, J.; de Miguel, S.; Yuste, J. Combination of antibodies and antibiotics as a promising strategy against multidrug-resistant pathogens of the respiratory tract. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azoulay-Dupuis, E.; Rieux, V.; Muffat-Joly, M.; Bedos, J.P.; Vallee, E.; Rivier, C.; Isturiz, R.; Carbon, C.; Moine, P. Relationship between capsular type, penicillin susceptibility, and virulence of human Streptococcus pneumoniae isolates in mice. Antimicrob. Agents Chemother. 2000, 44, 1575–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rieux, V.; Carbon, C.; Azoulay-Dupuis, E. Complex relationship between acquisition of beta-lactam resistance and loss of virulence in Streptococcus pneumoniae. J. Infect. Dis. 2001, 184, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Read, R.C.; Dull, P.; Bai, X.; Nolan, K.; Findlow, J.; Bazaz, R.; Kleinschmidt, A.; McCarthy, M.; Wang, H.; Toneatto, D.; et al. A phase III observer-blind randomized, controlled study to evaluate the immune response and the correlation with nasopharyngeal carriage after immunization of university students with a quadrivalent meningococcal ACWY glycoconjugate or serogroup B meningococcal vaccine. Vaccine 2017, 35, 427–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, H.S.; McMillan, M.; Koehler, A.P.; Lawrence, A.; Sullivan, T.R.; MacLennan, J.M.; Maiden, M.C.J.; Ladhani, S.N.; Ramsay, M.E.; Trotter, C.; et al. Meningococcal B Vaccine and Meningococcal Carriage in Adolescents in Australia. N. Engl. J. Med. 2020, 382, 318–327. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain (Serotype) | PEN | AMX | CTX | TET | CHL | ERY | LVX |
|---|---|---|---|---|---|---|---|
| 48 (23F) | 8 | 16 | 8 | 64 | 4 | >128 | 2 |
| 69 (19F) | 2 | 2 | 2 | 4 | 4 | >128 | 1 |
| 450 (11A) | 2 | 4 | 0.5 | 0.25 | 4 | 0.12 | 1 |
| 957 (3) | 0.015 | 0.015 | 0.015 | 0.5 | 4 | 0.25 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corsini, B.; Aguinagalde, L.; Ruiz, S.; Domenech, M.; Yuste, J. Vaccination with LytA, LytC, or Pce of Streptococcus pneumoniae Protects against Sepsis by Inducing IgGs That Activate the Complement System. Vaccines 2021, 9, 186. https://doi.org/10.3390/vaccines9020186
Corsini B, Aguinagalde L, Ruiz S, Domenech M, Yuste J. Vaccination with LytA, LytC, or Pce of Streptococcus pneumoniae Protects against Sepsis by Inducing IgGs That Activate the Complement System. Vaccines. 2021; 9(2):186. https://doi.org/10.3390/vaccines9020186
Chicago/Turabian StyleCorsini, Bruno, Leire Aguinagalde, Susana Ruiz, Mirian Domenech, and Jose Yuste. 2021. "Vaccination with LytA, LytC, or Pce of Streptococcus pneumoniae Protects against Sepsis by Inducing IgGs That Activate the Complement System" Vaccines 9, no. 2: 186. https://doi.org/10.3390/vaccines9020186
APA StyleCorsini, B., Aguinagalde, L., Ruiz, S., Domenech, M., & Yuste, J. (2021). Vaccination with LytA, LytC, or Pce of Streptococcus pneumoniae Protects against Sepsis by Inducing IgGs That Activate the Complement System. Vaccines, 9(2), 186. https://doi.org/10.3390/vaccines9020186