
Immunization with SP_1992 (DiiA) Protein of Streptococcus pneumoniae Reduces Nasopharyngeal Colonization and Protects against Invasive Disease in Mice



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Epitope/Immunogenicity Prediction
2.2. Immunization Experiments in Mice for Antibody Response Determination
2.3. Enzyme Linked Immunosorbent Assays
2.4. Western Blot
2.5. Bacterial Strains and Growth Conditions
2.6. Vaccination Experiments in Mice for Protection and Bacterial Clearance
3. Results
3.1. DiiA Is a Potential Vaccine Candidate
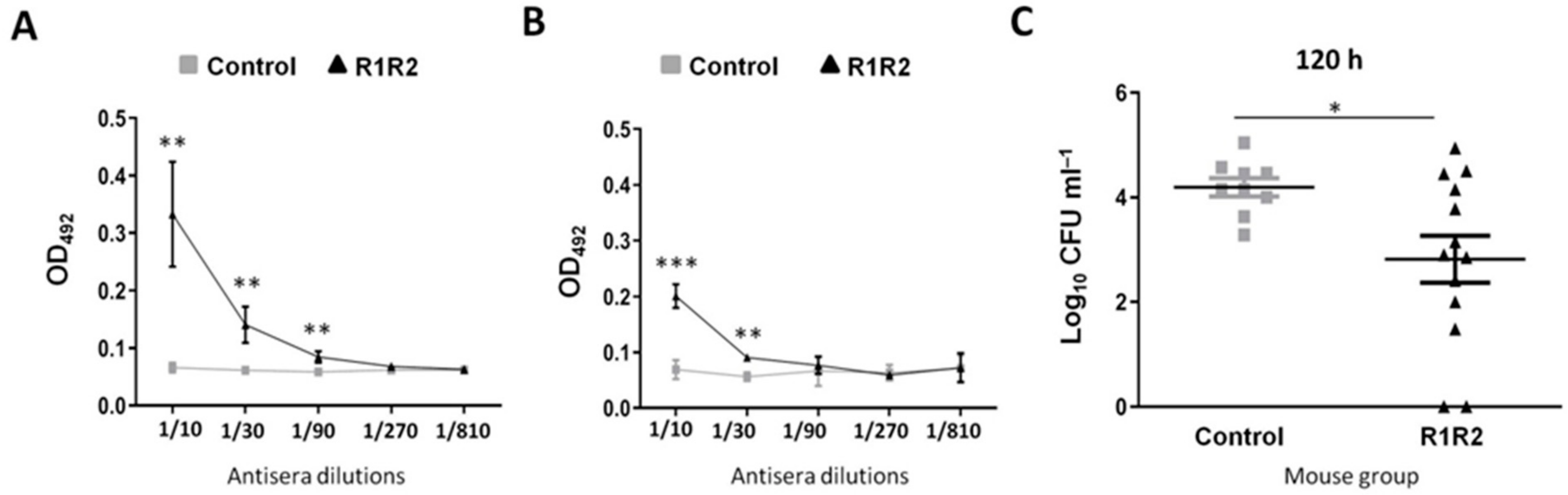
3.2. DiiA Protein Elicits a Strong and Diverse Immunogenic Response after Parenteral Immunization
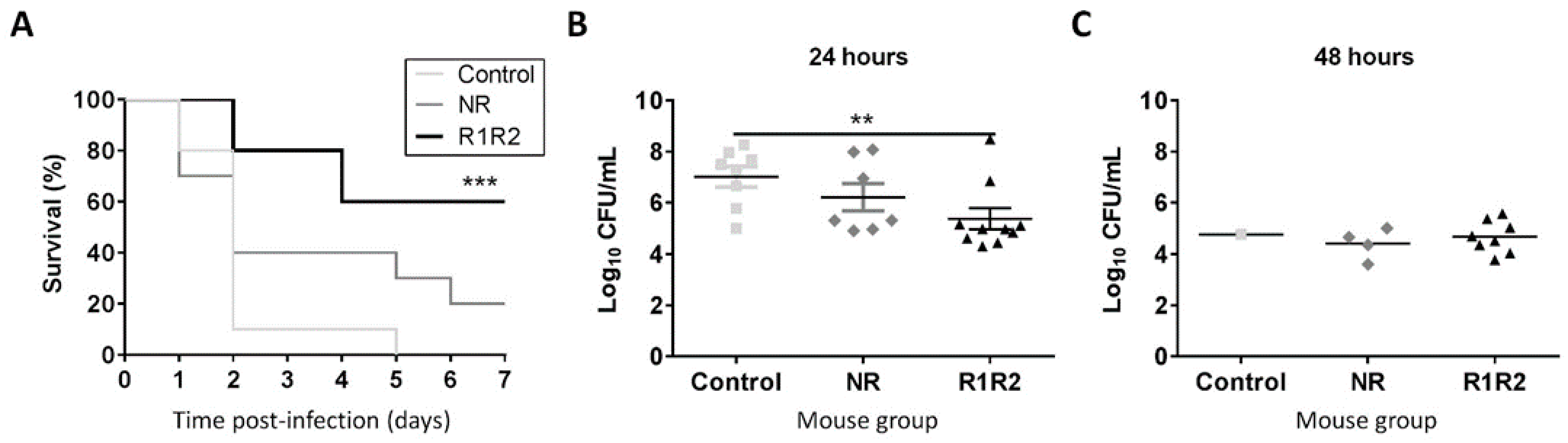
3.3. DiiA−R1R2 Protects Against Sepsis
3.4. DiiA−R1R2 Reduces Colonization in the Upper Respiratory Tract
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, L.; Oza, S.; Hogan, D.; Perin, J.; Rudan, I.; Lawn, J.E.; Cousens, S.; Mathers, C.; Black, R.E. Global, regional, and national causes of child mortality in 2000-13, with projections to inform post-2015 priorities: An updated systematic analysis. Lancet 2015, 385, 430–440. [Google Scholar] [CrossRef]
- Collaborators, G.L. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of lower respiratory tract infections in 195 countries: A systematic analysis for the Global Burden of Disease Study 2015. Lancet Infect. Dis. 2017, 17, 1133–1161. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, K.L.; Levine, O.S. Effectiveness of pneumococcal conjugate vaccine. Lancet 2006, 368, 1469–1470. [Google Scholar] [CrossRef]
- Ganaie, F.; Saad, J.S.; McGee, L.; van Tonder, A.J.; Bentley, S.D.; Lo, S.W.; Gladstone, R.A.; Turner, P.; Keenan, J.D.; Breiman, R.F.; et al. A New Pneumococcal Capsule Type, 10D, is the 100th Serotype and Has a Large cps Fragment from an Oral Streptococcus. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Aguinagalde, L.; Corsini, B.; Domenech, A.; Domenech, M.; Camara, J.; Ardanuy, C.; Garcia, E.; Linares, J.; Fenoll, A.; Yuste, J. Emergence of Amoxicillin-Resistant Variants of Spain9V-ST156 Pneumococci Expressing Serotype 11A Correlates with Their Ability to Evade the Host Immune Response. PLoS ONE 2015, 10, e0137565. [Google Scholar] [CrossRef] [PubMed]
- Andam, C.P.; Mitchell, P.K.; Callendrello, A.; Chang, Q.; Corander, J.; Chaguza, C.; McGee, L.; Beall, B.W.; Hanage, W.P. Genomic Epidemiology of Penicillin-Nonsusceptible Pneumococci with Nonvaccine Serotypes Causing Invasive Disease in the United States. J. Clin. Microbiol. 2017, 55, 1104–1115. [Google Scholar] [CrossRef] [Green Version]
- Ladhani, S.N.; Collins, S.; Djennad, A.; Sheppard, C.L.; Borrow, R.; Fry, N.K.; Andrews, N.J.; Miller, E.; Ramsay, M.E. Rapid increase in non-vaccine serotypes causing invasive pneumococcal disease in England and Wales, 2000-17: A prospective national observational cohort study. Lancet Infect. Dis. 2018, 18, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Jefferies, J.M.; Clarke, S.C.; Webb, J.S.; Kraaijeveld, A.R. Risk of red queen dynamics in pneumococcal vaccine strategy. Trends Microbiol. 2011, 19, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Feldman, C.; Anderson, R. Review: Current and new generation pneumococcal vaccines. J. Infect. 2014, 69, 309–325. [Google Scholar] [CrossRef] [Green Version]
- Giefing, C.; Meinke, A.L.; Hanner, M.; Henics, T.; Bui, M.D.; Gelbmann, D.; Lundberg, U.; Senn, B.M.; Schunn, M.; Habel, A.; et al. Discovery of a novel class of highly conserved vaccine antigens using genomic scale antigenic fingerprinting of pneumococcus with human antibodies. J. Exp. Med. 2008, 205, 117–131. [Google Scholar] [CrossRef]
- Rolo, D.; Ardanuy, C.; Fleites, A.; Martin, R.; Linares, J. Diversity of pneumococcal surface protein A (PspA) among prevalent clones in Spain. BMC Microbiol. 2009, 9, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannelli, F.; Oggioni, M.R.; Pozzi, G. Allelic variation in the highly polymorphic locus pspC of Streptococcus pneumoniae. Gene 2002, 284, 63–71. [Google Scholar] [CrossRef]
- Barocchi, M.A.; Ries, J.; Zogaj, X.; Hemsley, C.; Albiger, B.; Kanth, A.; Dahlberg, S.; Fernebro, J.; Moschioni, M.; Masignani, V.; et al. A pneumococcal pilus influences virulence and host inflammatory responses. Proc. Natl. Acad. Sci. USA 2006, 103, 2857–2862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamou, J.E.; Heinrichs, J.H.; Erwin, A.L.; Walsh, W.; Gayle, T.; Dormitzer, M.; Dagan, R.; Brewah, Y.A.; Barren, P.; Lathigra, R.; et al. Identification and characterization of a novel family of pneumococcal proteins that are protective against sepsis. Infect. Immun. 2001, 69, 949–958. [Google Scholar] [CrossRef] [Green Version]
- Chiang, M.H.; Sung, W.C.; Lien, S.P.; Chen, Y.Z.; Lo, A.F.; Huang, J.H.; Kuo, S.C.; Chong, P. Identification of novel vaccine candidates against Acinetobacter baumannii using reverse vaccinology. Hum. Vaccines Immunother. 2015, 11, 1065–1073. [Google Scholar] [CrossRef] [Green Version]
- Monterrubio-Lopez, G.P.; Gonzalez, Y.M.J.A.; Ribas-Aparicio, R.M. Identification of Novel Potential Vaccine Candidates against Tuberculosis Based on Reverse Vaccinology. BioMed Res. Int. 2015, 2015, 483150. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Naz, A.; Soares, S.C.; Bakhtiar, M.; Tiwari, S.; Hassan, S.S.; Hanan, F.; Ramos, R.; Pereira, U.; Barh, D.; et al. Pan-genome analysis of human gastric pathogen H. pylori: Comparative genomics and pathogenomics approaches to identify regions associated with pathogenicity and prediction of potential core therapeutic targets. BioMed Res. Int. 2015, 2015, 139580. [Google Scholar] [CrossRef] [Green Version]
- Kelly, D.F.; Rappuoli, R. Reverse vaccinology and vaccines for serogroup B Neisseria meningitidis. Adv. Exp. Med. Biol. 2005, 568, 217–223. [Google Scholar] [CrossRef]
- Hoskins, J.; Alborn, W.E., Jr.; Arnold, J.; Blaszczak, L.C.; Burgett, S.; DeHoff, B.S.; Estrem, S.T.; Fritz, L.; Fu, D.J.; Fuller, W.; et al. Genome of the bacterium Streptococcus pneumoniae strain R6. J. Bacteriol. 2001, 183, 5709–5717. [Google Scholar] [CrossRef] [Green Version]
- Tettelin, H.; Nelson, K.E.; Paulsen, I.T.; Eisen, J.A.; Read, T.D.; Peterson, S.; Heidelberg, J.; DeBoy, R.T.; Haft, D.H.; Dodson, R.J.; et al. Complete genome sequence of a virulent isolate of Streptococcus pneumoniae. Science 2001, 293, 498–506. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, S.; Hammerschmidt, S. Versatility of pneumococcal surface proteins. Microbiology 2006, 152, 295–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohler, S.; Voss, F.; Gomez Mejia, A.; Brown, J.S.; Hammerschmidt, S. Pneumococcal lipoproteins involved in bacterial fitness, virulence, and immune evasion. FEBS Lett. 2016, 590, 3820–3839. [Google Scholar] [CrossRef] [PubMed]
- Escolano-Martinez, M.S.; Domenech, A.; Yuste, J.; Cercenado, M.I.; Ardanuy, C.; Linares, J.; de la Campa, A.G.; Martin-Galiano, A.J. DiiA is a novel dimorphic cell wall protein of Streptococcus pneumoniae involved in invasive disease. J. Infect. 2016, 73, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Rosch, J.W.; Gao, G.; Ridout, G.; Wang, Y.D.; Tuomanen, E.I. Role of the manganese efflux system mntE for signalling and pathogenesis in Streptococcus pneumoniae. Mol. Microbiol. 2009, 72, 12–25. [Google Scholar] [CrossRef] [Green Version]
- Heath, C.J.; del Mar Cendra, M.; Watson, A.; Auger, J.P.; Pandey, A.; Tighe, P.; Christodoulides, M. Co-Transcriptomes of Initial Interactions In Vitro between Streptococcus Pneumoniae and Human Pleural Mesothelial Cells. PLoS ONE 2015, 10, e0142773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davydov Ia, I.; Tonevitskii, A.G. Linear B-cell epitope prediction. Mol. Biol. 2009, 43, 166–174. [Google Scholar] [CrossRef]
- Saha, S.; Raghava, G.P. Prediction of continuous B-cell epitopes in an antigen using recurrent neural network. Proteins 2006, 65, 40–48. [Google Scholar] [CrossRef]
- Larsen, J.E.; Lund, O.; Nielsen, M. Improved method for predicting linear B-cell epitopes. Immunome Res. 2006, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Kolaskar, A.S.; Tongaonkar, P.C. A semi-empirical method for prediction of antigenic determinants on protein antigens. FEBS Lett. 1990, 276, 172–174. [Google Scholar] [CrossRef] [Green Version]
- Saravanan, V.; Gautham, N. Harnessing Computational Biology for Exact Linear B-Cell Epitope Prediction: A Novel Amino Acid Composition-Based Feature Descriptor. OMICS 2015, 19, 648–658. [Google Scholar] [CrossRef]
- Yao, B.; Zhang, L.; Liang, S.; Zhang, C. SVMTriP: A method to predict antigenic epitopes using support vector machine to integrate tri-peptide similarity and propensity. PLoS ONE 2012, 7, e45152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynisson, B.; Alvarez, B.; Paul, S.; Peters, B.; Nielsen, M. NetMHCpan-4.1 and NetMHCIIpan-4.0: Improved predictions of MHC antigen presentation by concurrent motif deconvolution and integration of MS MHC eluted ligand data. Nucleic Acids Res. 2020, 48, W449–W454. [Google Scholar] [CrossRef]
- Sidney, J.; Steen, A.; Moore, C.; Ngo, S.; Chung, J.; Peters, B.; Sette, A. Five HLA-DP molecules frequently expressed in the worldwide human population share a common HLA supertypic binding specificity. J. Immunol. 2010, 184, 2492–2503. [Google Scholar] [CrossRef]
- Pan, X.; Zuallaert, J.; Wang, X.; Shen, H.B.; Campos, E.P.; Marushchak, D.O.; De Neve, W. ToxDL: Deep learning using primary structure and domain embeddings for assessing protein toxicity. Bioinformatics 2020, 36, 5159–5168. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, I.; Bangov, I.; Flower, D.R.; Doytchinova, I. AllerTOP v.2—A server for in silico prediction of allergens. J. Mol. Model. 2014, 20, 2278. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Raghava, G.P. AlgPred: Prediction of allergenic proteins and mapping of IgE epitopes. Nucleic Acids Res. 2006, 34, W202–W209. [Google Scholar] [CrossRef]
- Jomaa, M.; Yuste, J.; Paton, J.C.; Jones, C.; Dougan, G.; Brown, J.S. Antibodies to the iron uptake ABC transporter lipoproteins PiaA and PiuA promote opsonophagocytosis of Streptococcus pneumoniae. Infect. Immun. 2005, 73, 6852–6859. [Google Scholar] [CrossRef] [Green Version]
- Corsini, B.; Aguinagalde, L.; Ruiz, S.; Domenech, M.; Antequera, M.L.; Fenoll, A.; Garcia, P.; Garcia, E.; Yuste, J. Immunization with LytB protein of Streptococcus pneumoniae activates complement-mediated phagocytosis and induces protection against pneumonia and sepsis. Vaccine 2016, 34, 6148–6157. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Sevillano, E.; Moscoso, M.; Garcia, P.; Garcia, E.; Yuste, J. Nasopharyngeal colonization and invasive disease are enhanced by the cell wall hydrolases LytB and LytC of Streptococcus pneumoniae. PLoS ONE 2011, 6, e23626. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Sevillano, E.; Urzainqui, A.; Campuzano, S.; Moscoso, M.; Gonzalez-Camacho, F.; Domenech, M.; Rodriguez de Cordoba, S.; Sanchez-Madrid, F.; Brown, J.S.; Garcia, E.; et al. Pleiotropic effects of cell wall amidase LytA on Streptococcus pneumoniae sensitivity to the host immune response. Infect. Immun. 2015, 83, 591–603. [Google Scholar] [CrossRef] [Green Version]
- Cafini, F.; Yuste, J.; Gimenez, M.J.; Sevillano, D.; Aguilar, L.; Alou, L.; Ramos-Sevillano, E.; Torrico, M.; Gonzalez, N.; Garcia, E.; et al. Enhanced in vivo activity of cefditoren in pre-immunized mice against penicillin-resistant S. pneumoniae (serotypes 6B, 19F and 23F) in a sepsis model. PLoS ONE 2010, 5, e12041. [Google Scholar] [CrossRef] [Green Version]
- Mann, B.; Orihuela, C.; Antikainen, J.; Gao, G.; Sublett, J.; Korhonen, T.K.; Tuomanen, E. Multifunctional role of choline binding protein G in pneumococcal pathogenesis. Infect. Immun. 2006, 74, 821–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brueggemann, A.B.; Pai, R.; Crook, D.W.; Beall, B. Vaccine escape recombinants emerge after pneumococcal vaccination in the United States. PLoS Pathog. 2007, 3, e168. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Diaz, A.; Machado, M.P.; Camara, J.; Yuste, J.; Varon, E.; Domenech, M.; Del Grosso, M.; Marimon, J.M.; Cercenado, E.; Larrosa, N.; et al. Two multi-fragment recombination events resulted in the beta-lactam-resistant serotype 11A-ST6521 related to Spain9V-ST156 pneumococcal clone spreading in south-western Europe, 2008 to 2016. Eurosurveillance 2020, 25, 1900457. [Google Scholar] [CrossRef]
- McAllister, L.J.; Tseng, H.J.; Ogunniyi, A.D.; Jennings, M.P.; McEwan, A.G.; Paton, J.C. Molecular analysis of the psa permease complex of Streptococcus pneumoniae. Mol. Microbiol. 2004, 53, 889–901. [Google Scholar] [CrossRef]
- Kilian, M.; Tettelin, H. Identification of Virulence-Associated Properties by Comparative Genome Analysis of Streptococcus pneumoniae, S. pseudopneumoniae, S. mitis, Three S. oralis Subspecies, and S. infantis. mBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Salva-Serra, F.; Connolly, G.; Moore, E.R.B.; Gonzales-Siles, L. Detection of “Xisco” gene for identification of Streptococcus pneumoniae isolates. Diagn. Microbiol. Infect. Dis. 2018, 90, 248–250. [Google Scholar] [CrossRef] [PubMed]
- Bologa, M.; Kamtchoua, T.; Hopfer, R.; Sheng, X.; Hicks, B.; Bixler, G.; Hou, V.; Pehlic, V.; Yuan, T.; Gurunathan, S. Safety and immunogenicity of pneumococcal protein vaccine candidates: Monovalent choline-binding protein A (PcpA) vaccine and bivalent PcpA-pneumococcal histidine triad protein D vaccine. Vaccine 2012, 30, 7461–7468. [Google Scholar] [CrossRef]
- Kamtchoua, T.; Bologa, M.; Hopfer, R.; Neveu, D.; Hu, B.; Sheng, X.; Corde, N.; Pouzet, C.; Zimmermann, G.; Gurunathan, S. Safety and immunogenicity of the pneumococcal pneumolysin derivative PlyD1 in a single-antigen protein vaccine candidate in adults. Vaccine 2013, 31, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Bonam, S.R.; Partidos, C.D.; Halmuthur, S.K.M.; Muller, S. An Overview of Novel Adjuvants Designed for Improving Vaccine Efficacy. Trends Pharmacol. Sci. 2017, 38, 771–793. [Google Scholar] [CrossRef] [PubMed]
- Lottenbach, K.R.; Mink, C.M.; Barenkamp, S.J.; Anderson, E.L.; Homan, S.M.; Powers, D.C. Age-associated differences in immunoglobulin G1 (IgG1) and IgG2 subclass antibodies to pneumococcal polysaccharides following vaccination. Infect. Immun. 1999, 67, 4935–4938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uddin, S.; Borrow, R.; Haeney, M.R.; Moran, A.; Warrington, R.; Balmer, P.; Arkwright, P.D. Total and serotype-specific pneumococcal antibody titres in children with normal and abnormal humoral immunity. Vaccine 2006, 24, 5637–5644. [Google Scholar] [CrossRef] [PubMed]
- Shelly, M.A.; Jacoby, H.; Riley, G.J.; Graves, B.T.; Pichichero, M.; Treanor, J.J. Comparison of pneumococcal polysaccharide and CRM197-conjugated pneumococcal oligosaccharide vaccines in young and elderly adults. Infect. Immun. 1997, 65, 242–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuorimaa, T.; Kayhty, H.; Leroy, O.; Eskola, J. Tolerability and immunogenicity of an 11-valent pneumococcal conjugate vaccine in adults. Vaccine 2001, 19, 1863–1869. [Google Scholar] [CrossRef]
- Read, R.C.; Dull, P.; Bai, X.; Nolan, K.; Findlow, J.; Bazaz, R.; Kleinschmidt, A.; McCarthy, M.; Wang, H.; Toneatto, D.; et al. A phase III observer-blind randomized, controlled study to evaluate the immune response and the correlation with nasopharyngeal carriage after immunization of university students with a quadrivalent meningococcal ACWY glycoconjugate or serogroup B meningococcal vaccine. Vaccine 2017, 35, 427–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, H.S.; McMillan, M.; Koehler, A.P.; Lawrence, A.; Sullivan, T.R.; MacLennan, J.M.; Maiden, M.C.J.; Ladhani, S.N.; Ramsay, M.E.; Trotter, C.; et al. Meningococcal B Vaccine and Meningococcal Carriage in Adolescents in Australia. N. Engl. J. Med. 2020, 382, 318–327. [Google Scholar] [CrossRef]
- Bogaert, D.; De Groot, R.; Hermans, P.W. Streptococcus pneumoniae colonisation: The key to pneumococcal disease. Lancet Infect. Dis. 2004, 4, 144–154. [Google Scholar] [CrossRef]
- Ladhani, S.N.; Andrews, N.; Parikh, S.R.; Campbell, H.; White, J.; Edelstein, M.; Bai, X.; Lucidarme, J.; Borrow, R.; Ramsay, M.E. Vaccination of Infants with Meningococcal Group B Vaccine (4CMenB) in England. N. Engl. J. Med. 2020, 382, 309–317. [Google Scholar] [CrossRef]
- Martin-Galiano, A.J. The MiiA motif is a common marker present in polytopic surface proteins of oral and urinary tract invasive bacteria. Infect. Genet. Evol. 2017, 49, 283–292. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-Galiano, A.J.; Escolano-Martínez, M.S.; Corsini, B.; Campa, A.G.d.l.; Yuste, J. Immunization with SP_1992 (DiiA) Protein of Streptococcus pneumoniae Reduces Nasopharyngeal Colonization and Protects against Invasive Disease in Mice. Vaccines 2021, 9, 187. https://doi.org/10.3390/vaccines9030187
Martín-Galiano AJ, Escolano-Martínez MS, Corsini B, Campa AGdl, Yuste J. Immunization with SP_1992 (DiiA) Protein of Streptococcus pneumoniae Reduces Nasopharyngeal Colonization and Protects against Invasive Disease in Mice. Vaccines. 2021; 9(3):187. https://doi.org/10.3390/vaccines9030187
Chicago/Turabian StyleMartín-Galiano, Antonio J., María S. Escolano-Martínez, Bruno Corsini, Adela G. de la Campa, and José Yuste. 2021. "Immunization with SP_1992 (DiiA) Protein of Streptococcus pneumoniae Reduces Nasopharyngeal Colonization and Protects against Invasive Disease in Mice" Vaccines 9, no. 3: 187. https://doi.org/10.3390/vaccines9030187
APA StyleMartín-Galiano, A. J., Escolano-Martínez, M. S., Corsini, B., Campa, A. G. d. l., & Yuste, J. (2021). Immunization with SP_1992 (DiiA) Protein of Streptococcus pneumoniae Reduces Nasopharyngeal Colonization and Protects against Invasive Disease in Mice. Vaccines, 9(3), 187. https://doi.org/10.3390/vaccines9030187