
Cell Culture-Derived Tilapia Lake Virus-Inactivated Vaccine Containing Montanide Adjuvant Provides High Protection against Viral Challenge for Tilapia

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Fish, Cell Lines, and Virus
2.2. Virus Cultivation
2.3. Virus Inactivation
2.4. Preparation of Inactivated Vaccine and Tilapia Immunization
2.5. Sample Collection
2.6. Measurement of Antibody Response by ELISA
2.7. Serum Neutralization Test (SNT)
2.8. Determination of Immune-Related Genes Expression by RT-qPCR
2.9. Protection against Lethal Challenge and Determination of Viral Load
2.10. Statistical Analysis
3. Results
3.1. Virus Inactivation
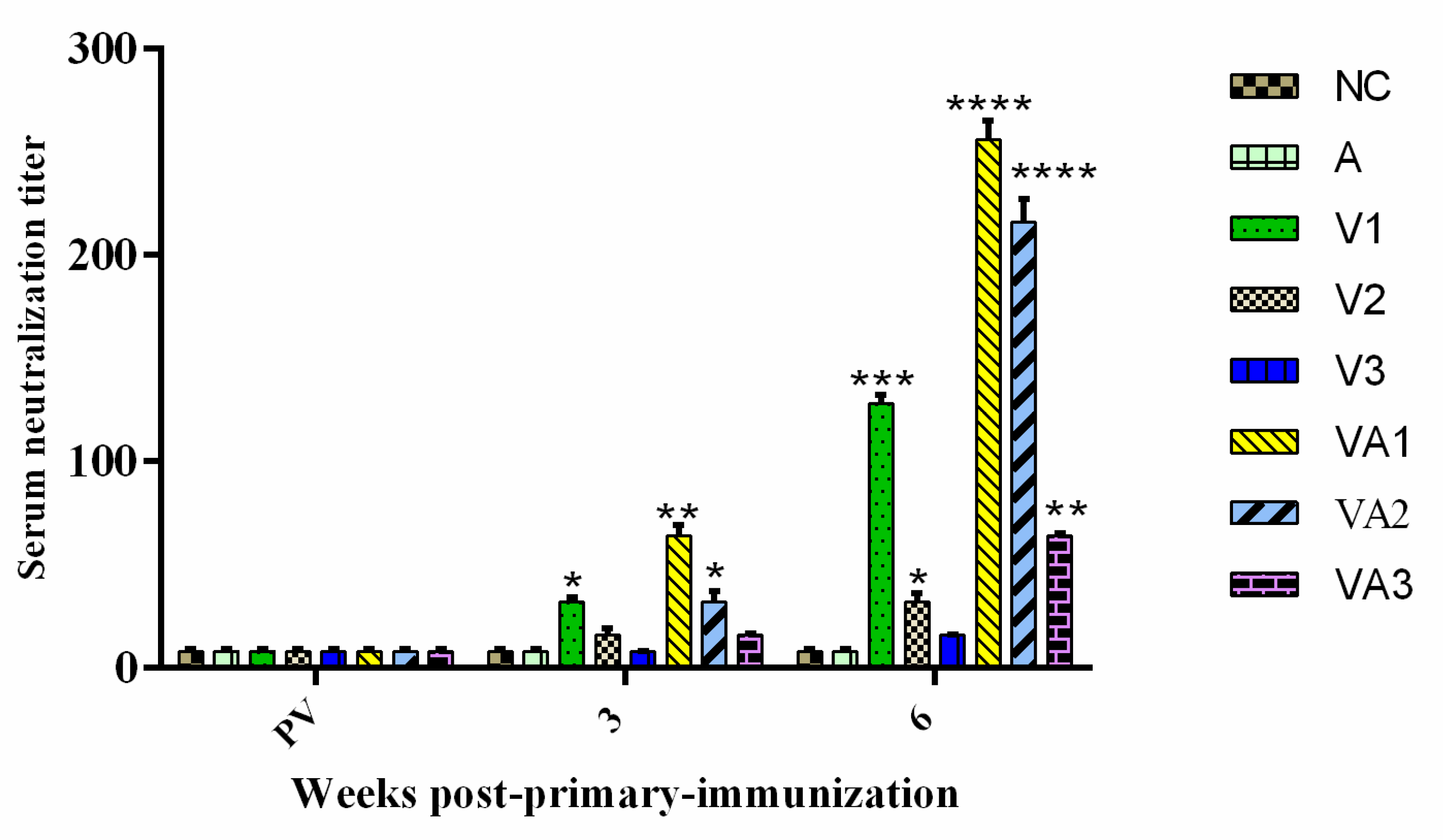
3.2. Detection of Serum IgM and Neutralizing Antibody
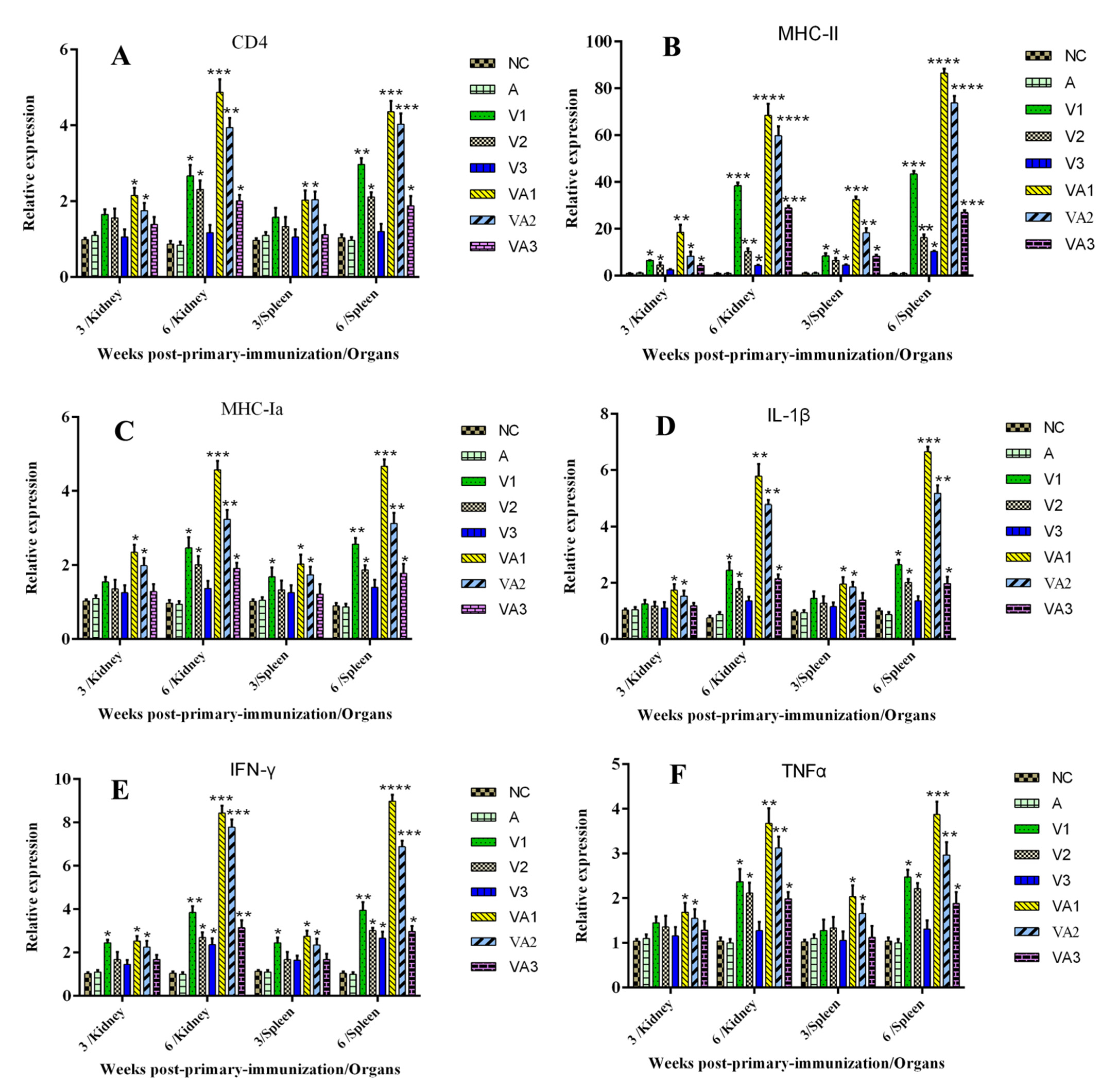
3.3. Immune-Related Gene Expression
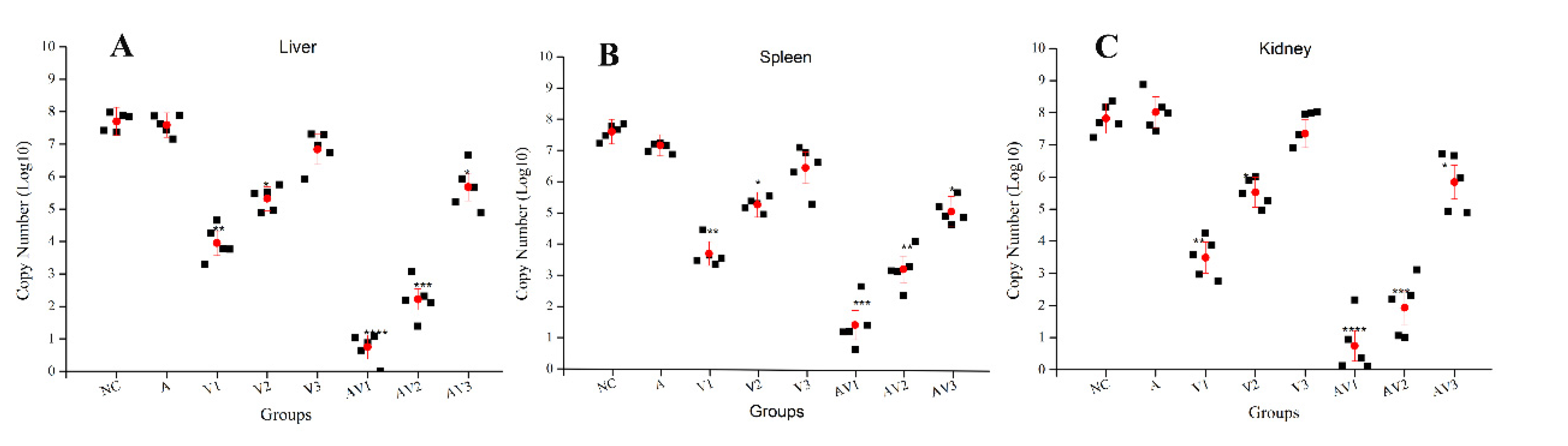
3.4. Determination of Viral Load in Different Tissues after Challenge with TiLV
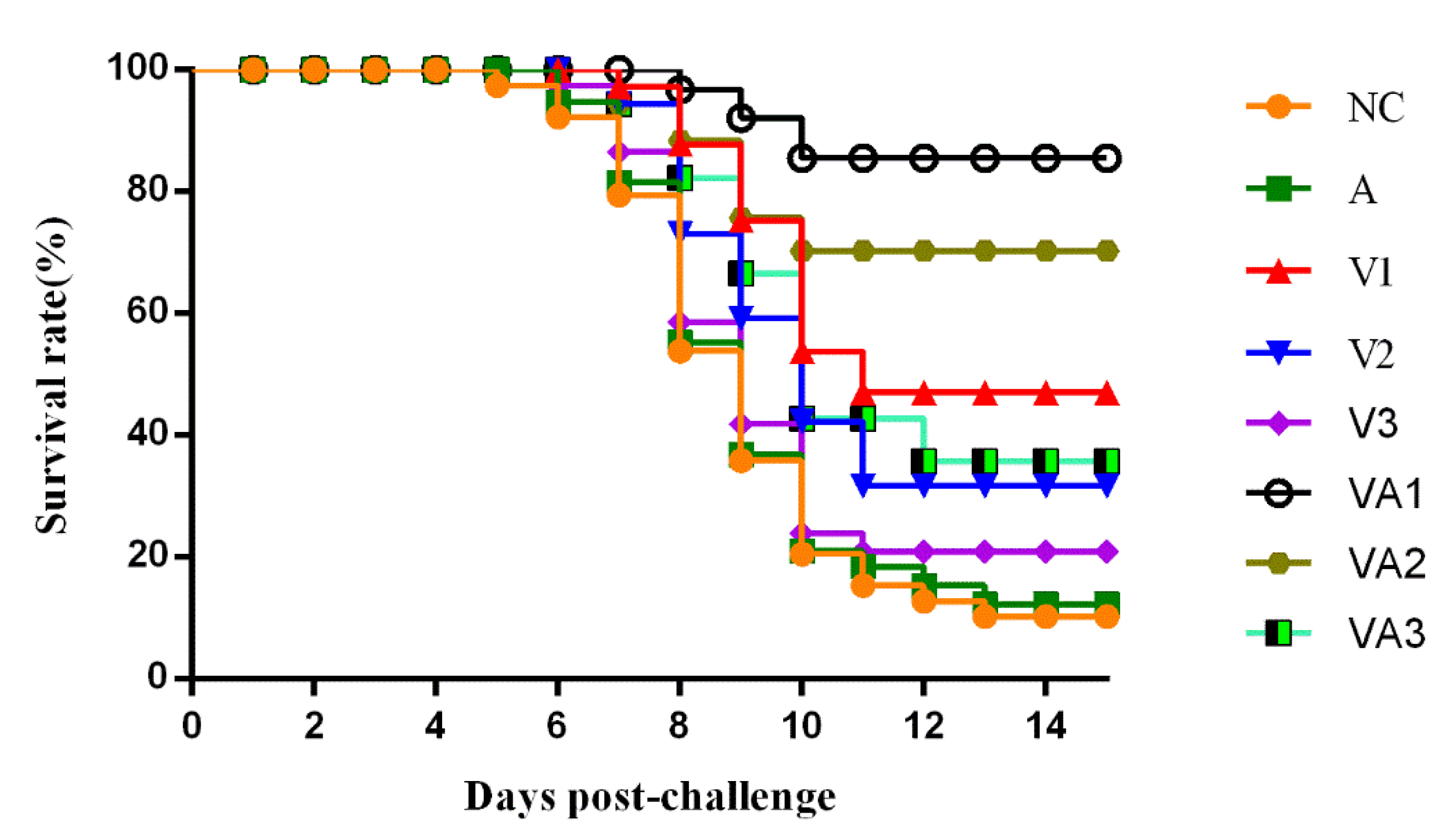
3.5. Protective Effectiveness of the Inactivated Vaccine against TiLV Challenge
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture, 2016; Contributing to Food Security and Nutrition for All; Food and Agriculture Organization of the United Nations Publishing: Rome, Italy, 2016. [Google Scholar]
- FAO. Report of the FAO-INERA Workshop on Good Agricultural Practices; Food and Agriculture Organization of the United Nations Publishing: Burkina FasoBobo-Dioulasso, Burkina Faso, 2004. [Google Scholar]
- Dinesh, R.; George, M.R.; KRJohn, K.R.; Abraham, S. TiLV-a worldwide menace to tilapiine aquaculture. J. Entomol. Zool. Stud. 2017, 5, 605–607. [Google Scholar]
- Ngo, V.H. Research findings from the use of probiotics in tilapia aquaculture: A review. Fish Shellfish Immunol. 2015, 45, 592–597. [Google Scholar]
- Wang, W.; Ding, J.L.; Fang, J.C. The current situation and analysis of the standardization of tilapia industry. J. Shanghai Ocean Univ. 2012, 21, 976–981. [Google Scholar]
- Woo, P.T.K.; Cipriano, R.C. Fish Viruses and Bacteria: Pathobiology and Protection; CABI Publishing: London, UK, 2017. [Google Scholar]
- Mian, G.; Godoy, D.; Leal, C.; Yuhara, T.; Costa, G.; Figueiredo, H.C.P. Aspects of the natural history and virulence of S. agalactiae infection in Nile tilapia. Vet. Microbiol. 2009, 136, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Assis, G.B.N.; Pereira, F.L.; Figueiredo, H.C.P.; Leal, C.A.G.; Tavares, G.C. Natural coinfection by Streptococcus agalactiae and Francisella noatunensis subsp. orientalis in farmed Nile tilapia (Oreochromis niloticus L.). J. Fish Dis. 2016, 40, 51–63. [Google Scholar] [CrossRef]
- Soto-Rodriguez, S.A.; Cabanillas-Ramos, J.; Alcaraz, U.; Gomez-Gil, B.; Romalde, J.L. Identification and virulence of Aeromonas dhakensis, Pseudomonas mosselii andMicrobacterium paraoxydans isolated from Nile tilapia, Oreochromis niloticus, cultivatedin Mexico. J. Appl. Microbiol. 2013, 115, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.A.; Noureldin, E.; Mahmoud, M.A.; Fita, N.A. Molecular identification and epizootiology of Aeromonas veronii infection among farmed Oreochromis niloticus in Eastern Province, KSA. Egypt. J. Aquat. Res. 2017, 43, 161–167. [Google Scholar] [CrossRef]
- Figueiredo, H.C.; Klesius, P.H.; Arias, C.R.; Evans, J.; Shoemaker, C.A.; Pereira, D.J., Jr.; Peixoto, M.T.D. Isolation and characterization of strains of Flavobacterium columnare from Brazil. J. Fish Dis. 2005, 28, 199–204. [Google Scholar] [CrossRef]
- Eyngor, M.; Zamostiano, R.; Tsofack, J.E.K.; Berkowitz, A.; Bercovier, H.; Tinman, S.; Lev, M.; Hurvitz, A.; Galeotti, M.; Bacharach, E.; et al. Identification of a Novel RNA Virus Lethal to Tilapia. J. Clin. Microbiol. 2014, 52, 4137–4146. [Google Scholar] [CrossRef] [Green Version]
- FAO. The State of World Fisheries and Aquaculture, 2018: Meeting the Sustainable Development Goals; Food and Agriculture Organization of the United Nations Publishing: Rome, Italy, 2018. [Google Scholar]
- Bacharach, E.; Mishra, N.; Briese, T.; Zody, M.C.; Tsofack, J.E.K.; Zamostiano, R.; Berkowitz, A.; Ng, J.; Nitido, A.; Corvelo, A.; et al. Characterization of a Novel Orthomyxo-like Virus Causing Mass Die-Offs of Tilapia. mBio 2016, 7, e00431-16. [Google Scholar] [CrossRef] [Green Version]
- Surachetpong, W.; Janetanakit, T.; Nonthabenjawan, N.; Tattiyapong, P.; Sirikanchana, K.; Amonsin, A. Outbreaks of Tilapia Lake Virus Infection, Thailand, 2015–2016. Emerg. Infect. Dis. 2017, 23, 1031–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acharya, V.; Chakraborty, H.J.; Rout, A.K.; Balabantaray, S.; Behera, B.K.; Das, B.K. Structural Characterization of Open Reading Frame-Encoded Functional Genes from Tilapia Lake Virus (TiLV). Mol. Biotechnol. 2019, 61, 945–957. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, H.W.; Kabuusu, R.M.; Beltran, S.; Reyes, E.; Lince, J.A.; Delpozo, J. Syncytial hepatitis of farmed tilapia, Oreochromis niloticus (L.): A case report. J. Fish Dis. 2014, 37, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Del-Pozo, J.; Mishra, N.; Kabuusu, R.; Cheetham, S.; Eldar, A.; Bacharach, E.; Lipkin, W.I.; Ferguson, H.W. Syncytial Hepatitis of Tilapia (Oreochromis niloticus L.) is Associated With Orthomyxovirus-Like Virions in Hepatocytes. Vet. Pathol. 2016, 54, 164–170. [Google Scholar] [CrossRef] [Green Version]
- ICTV, International Committee on Taxonomy of Viruses. Available online: https://talk.ictvonline.org/ (accessed on 9 July 2020).
- Jansen, M.D.; Mohan, C.V. Tilapia lake virus (TiLV): Literature review. In Working Paper: FISH-04; CGIAR Research Program on Fish Agri-Food Systems: Penang, Malaysia, 2017. [Google Scholar]
- Dong, H.; Siriroob, S.; Meemetta, W.; Santimanawong, W.; Gangnonngiw, W.; Pirarat, N.; Khunrae, P.; Rattanarojpong, T.; Vanichviriyakit, R.; Senapin, S. Emergence of tilapia lake virus in Thailand and an alternative semi-nested RT-PCR for detection. Aquaculture 2017, 476, 111–118. [Google Scholar] [CrossRef]
- Thammatorn, W.; Rawiwan, P.; Surachetpong, W. Minimal risk of tilapia lake virus transmission via frozen tilapia fillets. J. Fish Dis. 2018, 42, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Tattiyapong, P.; Dachavichitlead, W.; Surachetpong, W. Experimental infection of Tilapia Lake Virus (TiLV) in Nile tilapia (Oreochromis niloticus) and red tilapia (Oreochromis spp.). Vet. Microbiol. 2017, 207, 170–177. [Google Scholar] [CrossRef]
- Koesharyani, I.; Gardenia, L.; Widowati, Z.; Khumaira, K.; Rustianti, D. Studi Kasus Infeksi Tilapia Lake Virus (Tilv) Pada Ikan Nila (Oreochromis niloticus). J. Ris. Akuakultur 2018, 13, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Mugimba, K.K.; Chengula, A.A.; Wamala, S.; Mwega, E.D.; Kasanga, C.J.; Byarugaba, D.K.; Mdegela, R.H.; Tal, S.; Bornstein, B.; Dishon, A.; et al. Detection of tilapia lake virus (TiLV) infection by PCR in farmed and wild Nile tilapia (Oreochromis niloticus) from Lake Victoria. J. Fish Dis. 2018, 41, 1181–1189. [Google Scholar] [CrossRef]
- Nicholson, P.; Fathi, M.A.; Fischer, A.; Mohan, C.; Schieck, E.; Mishra, N.; Heinimann, A.; Frey, J.; Wieland, B.; Jores, J. Detection of Tilapia Lake Virus in Egyptian fish farms experiencing high mortalities in 2015. J. Fish Dis. 2017, 40, 1925–1928. [Google Scholar] [CrossRef]
- OIE. Tilapia Lake Virus Disease (TiLV), United States of America. Immediate Notification; OIE: Paris, France, 2019; Available online: http://www.oie.int/wahis_2/public/wahid.php/Reviewreport/Review?page_refer=MapFullEventReport&reportid=30412 (accessed on 8 March 2020).
- Jansen, M.D.; Dong, H.T.; Mohan, C.V. Tilapia lake virus: A threat to the global tilapia industry? Rev. Aquac. 2019, 11, 725–739. [Google Scholar] [CrossRef]
- World Organisation for Animal Health (OIE). Tilapia Lake Virus, United States of America- Immediate Notification; OIE: Paris, France, 2019. [Google Scholar]
- Dong, H.T.; Ataguba, G.A.; Khunrae, P.; Rattanarojpong, T.; Senapin, S. Evidence of TiLV infection in tilapia hatcheries in Thailand from 2012 to 2017 reveals probable global spread of the disease. Aquaculture 2017, 479, 579–583. [Google Scholar] [CrossRef]
- Fathi, M.; Dickson, C.; Dickson, M.; Leschen, W.; Baily, J.; Muir, F.; Ulrich, K.; Weidmann, M. Identification of tilapia lake virus in Egypt in Nile tilapia affected by ‘summer mortality’ syndrome. Aquaculture 2017, 473, 430–432. [Google Scholar] [CrossRef]
- Pulido, L.L.H.; Mora, C.M.; Hung, A.L.; Dong, H.T.; Senapin, S. Tilapia lake virus (TiLV) from Peru is genetically close to the Israeli isolates. Aquaculture 2019, 510, 61–65. [Google Scholar] [CrossRef]
- Nanthini, R.; Majeed, S.A.; Vimal, S.; Taju, G.; Sivakumar, S.; Kumar, S.S.; Pillai, D.; Sneha, K.G.; Rakesh, C.G.; Hameed, A.S.S. In vitro propagation of tilapia lake virus in cell lines developed from Oreochromis mossambicus. J. Fish Dis. 2019, 42, 1–10. [Google Scholar] [CrossRef]
- Azmai, M.N.A.; Koh, C.; Nurliyana, M.; Suhaiba, M.; Nor-Amalina, Z.; Santha, S.; Diyana-Nadhirah, K.; Yusof, M.; Ina-Salwany, M.; Saad, M.Z. A case of natural co-infection of Tilapia Lake Virus and Aeromonas veronii in a Malaysian red hybrid tilapia (Oreochromis niloticus × O. mossambicus) farm experiencing high mortality. Aquaculture 2018, 485, 12–16. [Google Scholar]
- Senapin, S.; Shyam, K.; Meemetta, W.; Rattanarojpong, T.; Dong, H.T. Inapparent infection cases of tilapia lake virus (TiLV) in farmed tilapia. Aquaculture 2018, 487, 51–55. [Google Scholar] [CrossRef]
- Liamnimitr, P.; Thammatorn, W.; U-thoomporn, S.; Tattiyapong, P.; Surachetpong, W. Non-lethal sampling for Tilapia Lake Virus detection by RT-qPCR and cell culture. Aquaculture 2018, 486, 75–80. [Google Scholar] [CrossRef]
- Yamkasem, J.; Tattiyapong, P.; Kamlangdee, A.; Surachetpong, W. Evidence of potential vertical transmission of tilapia lake virus. J. Fish Dis. 2019, 42, 1293–1300. [Google Scholar] [CrossRef]
- Pamela, N.; Natthawadee, M.; Phitchaya, J.; Puntanat, T.; Win, S. Coinfection of tilapia lake virus and Aeromonas hydrophila synergisticallyincreased mortality and worsened the disease severity in tilapia (Oreochromis spp.). Aquaculture 2020, 520, 734746. [Google Scholar]
- FAO. Global Aquaculture Production; FAO: Rome, Italy, 2017. [Google Scholar]
- OIE. Tilapia Lake Virus (TiLV)-A Novel Orthomyxo-Like Virus; OIE: Paris, France, 2017. [Google Scholar]
- NACA. Urgent Update on Possible Worldwide Spread of Tilapia Lake Virus (TiLV) [R]; NACA: Bangkok, Thailand, 2017. [Google Scholar]
- Zeng, W.; Wang, Q.; Wang, Y.; Zhao, C.; Li, Y.; Shi, C.; Wu, S.; Song, X.; Huang, Q.; Li, S. Immunogenicity of a cell culture-derived inactivated vaccine against a common virulent isolate of grass carp reovirus. Fish Shellfish Immunol. 2016, 54, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Hjeltnes, B.; Bornø, G.; Jansen, M.D.; Haukaas, A.; Walde, C. The Health Situation in Norwegian Aquaculture 2016; Norwegian Veterinary Institute Report Series; Norwegian Veterinary Institute: Oslo, Norway, 2017. [Google Scholar]
- Jee, Y.H.; Mun-Gyeong, K.; Yu, J.K.; Sung, H.J.; Myoung, A.P.; Maeng, H.S. Montanide IMS 1312 VG adjuvant enhances the efficacy of immersion vaccine of inactivated viral hemorrhagic septicemia virus (VHSV) in olive flounder, Paralichthys olivaceus. Fish Shellfish Immunol. 2017, 60, 420–425. [Google Scholar]
- Matsuura, Y.; Terashima, S.; Takano, T.; Matsuyama, T. Current status of fish vaccines in Japan. Fish Shellfish Immunol. 2019, 95, 236–247. [Google Scholar] [CrossRef]
- Wang, Q.C.; Ji, W.; Xu, Z. Current use and development of fish vaccines in China. Fish Shellfish Immunol. 2020, 96, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Evensen, Ø.; Leong, J.A. DNA vaccines against viral diseases of farmed fish. Fish Shellfish Immunol. 2013, 35, 1751–1758. [Google Scholar] [CrossRef] [PubMed]
- Salonius, K.; Simard, N.; Harland, R.; Ulmer, J.B. The road to licensure of a DNA vaccine. Curr. Opin. Investig. Drugs 2007, 8, 635–641. [Google Scholar]
- Sommerset, I.; Krossøy, B.; Biering, E.; Frost, P. Vaccines for fish in aquaculture. Expert Rev. Vaccines 2005, 4, 89–101. [Google Scholar] [CrossRef]
- Brudeseth, B.E.; Wiulsrod, R.; Fredriksen, B.N.; Lindmo, K.; Lokling, K.E.; Bordevik, M. Status and future perspectives of vaccines for industrialised fin-fish farming. Fish Shellfish Immunol. 2013, 121, 55–73. [Google Scholar] [CrossRef]
- Monath, T.P.; Fowler, E.; Johnson, C.T.; Balser, J.; Morin, M.J.; Sisti, M.; Trent, D.W. An Inactivated Cell-Culture Vaccine against Yellow Fever. New Engl. J. Med. 2011, 364, 1326–1333. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Zeng, W.; Yin, J.; Li, Y.; Ren, Y.; Shi, C.; Bergmann, S.M.; Zhu, X. Establishment and characterization of a cell line from tilapia brain for detection of tilapia lake virus. J. Fish Dis. 2018, 41, 1803–1809. [Google Scholar] [CrossRef]
- Hu, H.; Zeng, W.; Wang, Y.; Wang, Q.; Bergmann, S.M.; Yin, J.; Li, Y.; Chen, X.; Gao, C.; Zhang, D.; et al. Development and application of a recombinant protein-based indirect ELISA for detection of anti-tilapia lake virus IgM in sera from tilapia. Aquaculture 2020, 520, 734756. [Google Scholar] [CrossRef]
- Minetaro, A.; Masae, I.; Takaji, W.; Hiroyuki, S. Development of a Poliovirus Neutralization Test with Poliovirus Pseudovirus for Measurement of Neutralizing Antibody Titer in Human Serum. Clin. Vaccine Immunol. 2011, 18, 1889–1894. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2011, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Gallage, S.; Katagiri, T.; Endo, M.; Futami, K.; Endo, M.; Maita, M. Influence of moderate hypoxia on vaccine efficacy against Vibrio anguillarum in Oreochromis niloticus (Nile tilapia). Fish Shellfish Immunol. 2016, 51, 271–281. [Google Scholar] [CrossRef]
- Yao, Y.Y.; Chen, D.D.; Cui, Z.W.; Zhang, X.Y.; Zhou, Y.Y.; Guo, X.; Li, A.H.; Zhang, Y.A. Oral vaccination of tilapi against Streptococcus agalactiae using Bacillus subtilis spores expressing Sip. Fish Shellfish Immunol. 2019, 86, 999–1008. [Google Scholar] [CrossRef]
- Mo, X.B.; Wang, J.; Guo, S.; Li, A.X. Potential of naturally attenuated Streptococcus agalactiae as a live vaccine in Nile tilapia (Oreochromis niloticus). Aquaculture 2020, 518, 734774. [Google Scholar] [CrossRef]
- Amend, D.F. Potency testing of fish vaccines. Dev. Biol. Stand. 1981, 49, 447–454. [Google Scholar]
- Ellis, A.E. Fish Vaccination; Academic Press: London, UK, 1988; pp. 2–20. [Google Scholar]
- Huang, H.-Y.; Chen, Y.-C.; Wang, P.-C.; Tsai, M.-A.; Yeh, S.-C.; Liang, H.-J.; Chen, S.-C. Efficacy of a formalin-inactivated vaccine against Streptococcus iniae infection in the farmed grouper Epinephelus coioides by intraperitoneal immunization. Vaccine 2014, 32, 7014–7020. [Google Scholar] [CrossRef]
- Nayak, S.K.; Shibasaki, Y.; Nakanishi, T. Immune responses to live and inactivated Nocardia seriolae and protective effect of recombinant interferon gamma (rIFNγ) against nocardiosis in ginbuna crucian carp. Fish Shellfish Immunol. 2014, 39, 354–364. [Google Scholar] [CrossRef]
- Tauffer, F.; El-Bacha, T.; Da Poian, A.T. Advances in the development of inactivated virus vaccines. Recent Patents Anti-Infect. Drug Discov. 2006, 1, 291–296. [Google Scholar] [CrossRef]
- Frazatti-Gallina, N.M.; Mourão-Fuches, R.M.; Paoli, R.L.; Silva, M.L.; Miyaki, C.; Valentini, E.J.; Raw, I.; Higashi, H.G. Vero-cell rabies vaccine produced using serum-free medium. Vaccine 2004, 3, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Bahnemann, H.G. Inactivation of viral antigens for vaccine preparation with particular reference to the application of binary ethylenimine. Vaccine 1990, 8, 299–303. [Google Scholar] [CrossRef]
- Luo, S.; Yan, L.; Zhang, X.; Yuan, L.; Fang, Q.; Zhang, Y.A. Yeast surface display of capsid protein VP7 of grass carp reovirus: Fundamental investigation for the developmentof vaccine against hemorrhagic disease. J. Microbiol. Biotechnol. 2015, 25, 2135–2145. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.C.; Silva, A.N.; Souza, M.C.; Silva, M.V.; Neves, P.P.; Silva, A.A. An inactivated yellow fever 17DD vaccine cultivated in Vero cell cultures. Vaccine 2015, 33, 4261–4268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delrue, I.; Verzele, D.; Madder, A.; Nauwynck, H.J. Inactivated virus vaccines from chemistry to prophylaxis: Merits, risks and challenges. Expert Rev. Vaccines 2012, 11, 695–719. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Bian, Q.; Zeng, W.; Ren, P.; Sun, H.; Lin, Z.; Tang, Z.; Zhou, X.; Wang, Q.; Wang, Y.; et al. Oral delivery of Bacillus subtilis spores expressing grass carp reovirus VP4 protein producesprotection against grass carp reovirus infection. Fish Shellfish Immunol. 2019, 84, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Secombes, C.J. The function of fish cytokines. Biology 2016, 5, 23. [Google Scholar] [CrossRef]
- Poli, G.; Vicenzi, E.; Ghezzi, S.; Lazzarin, A. Cytokines in the acquired immunodeficiency syndrome and other infectious diseases. Int. J. Clin. Lab. Res. 1995, 25, 128–134. [Google Scholar] [CrossRef]
- Øvergård, A.-C.; Nepstad, I.; Nerland, A.H.; Patel, S. Characterisation and expression analysis of the Atlantic halibut (Hippoglossus hippoglossus L.) cytokines: IL-1b, IL-6, IL-11, IL-12b and IFNg. Mol. Biol. Rep. 2012, 39, 2201–2213. [Google Scholar] [CrossRef] [Green Version]
- Rao, Y.; Su, J. Insights into the antiviral immunity against grass carp (Ctenopharyngodon idella) reovirus (GCRV) in grass carp. J. Immunol. Res. 2015, 2015, 670437. [Google Scholar] [CrossRef] [Green Version]
- Somamoto, T.; Yoshiura, Y.; Nakanishi, T.; Ototake, M. Molecular cloning and characterization of two types of CD8? From ginbuna crucian carp. Dev. Comp. Immunol. 2005, 29, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Wang, Y.; Wang, X.; Chen, C.; Zhou, H. Characterization of grass carp (Ctenopharyngodon idellus) Foxp1a/1b/2: Evidence for their involvement in the activation of peripheral blood lymphocyte subpopulations. Fish Shellfish Immunol. 2010, 28, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.-C.; Gao, F.-Y.; Lu, M.; Ye, X.; Zhu, H.-P.; Ke, X.-L. Major histocompatibility complex class IIA and IIB genes of Nile tilapia Oreochromis niloticus: Genomic structure, molecular polymorphism and expression patterns. Fish Shellfish Immunol. 2013, 34, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Du, H.; Liu, L.; You, X.; Wu, M.; Liao, Z. MHC class II alpha, beta and MHC class II-associated invariant chains from Chinese sturgeon (Acipenser sinensis) and their response to immune stimulation. Fish Shellfish Immunol. 2017, 70, 1–12. [Google Scholar] [CrossRef]
- O’Hagan, D.T.; Singh, M. Microparticles as vaccine adjuvant and delivery systems. Expert Rev. Vaccines 2003, 2, 269–283. [Google Scholar] [CrossRef]
- Aucouturier, J.; Ganne, V.; Laval, A. Efficacy and safety of new adjuvants. Ann. N. Y. Acad. Sci. 2009, 916, 600–604. [Google Scholar] [CrossRef]
- Khimmakthong, U.; Deshmukh, S.; Chettri, J.K.; Kania, W.K.; Dalsgaard, I.; Buchmann, K. Tissue specific uptake of inactivated and live Yersinia ruckeri in rainbow trout (Oncorhynchus mykiss): Visualization by immunohistochemistry and in situ hybridization. Microb. Pathog. 2013, 59–60, 33–41. [Google Scholar] [CrossRef]
- Soltani, M.; Shafiei, S.; Yosefi, P.; Mosavi, S.; Mokhtari, A. Effect of Montanide™ IMS 1312 VG adjuvant on efficacy of Yersinia ruckeri vaccine in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2014, 37, 60–65. [Google Scholar] [CrossRef]
- Sá, E.; Silva, M.; Ellis, A.; Karaca, K.; Minke, J.; Nordgren, R.; Wu, S.; Swayne, D.E. Domestic goose as a model for West Nile virus vaccine efficacy. Vaccine 2013, 31, 1045–1050. [Google Scholar] [CrossRef] [PubMed]
- Angenvoort, J.; Fischer, D.; Fast, C.; Ziegler, U.; Eiden, M.; De La Fuente, J.G.; Lierz, M.; Groschup, M.H. Limited efficacy of West Nile virus vaccines in large falcons (Falco spp.). Vet. Res. 2014, 45, 41. [Google Scholar] [CrossRef] [Green Version]
- Tsofack, J.E.K.; Zamostiano, R.; Watted, S.; Berkowitz, A.; Rosenbluth, E.; Mishra, N.; Briese, T.; Lipkin, W.I.; Kabuusu, R.M.; Ferguson, H.; et al. Detection of Tilapia Lake Virus in Clinical Samples by Culturing and Nested Reverse Transcription-PCR. J. Clin. Microbiol. 2016, 55, 759–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inactivators | Groups | Inactivation Temperature (°C) | Concentration | Inactivation Time (h) | Infectivity | Protection Rate (%) |
|---|---|---|---|---|---|---|
| Formaldehyde | 1 | 37 | 1:2000 | 12 | + | / |
| 24 | + | / | ||||
| 36 | + | / | ||||
| 48 | + | / | ||||
| 60 | − * | 15 | ||||
| 2 | 1:1000 | 12 | + | / | ||
| 24 | + | / | ||||
| 36 | − * | 30 | ||||
| 48 | − | 20 | ||||
| 60 | − | 15 | ||||
| 3 | 1:500 | 12 | + | / | ||
| 24 | − * | 30 | ||||
| 36 | − | 20 | ||||
| 48 | − | 20 | ||||
| 60 | − | 10 | ||||
| BPL | 4 | 4 | 1:3000 | 12 | + | / |
| 24 | + | / | ||||
| 36 | + | / | ||||
| 48 | + | 35 | ||||
| 60 | − * | 25 | ||||
| 5 | 1:2000 | 12 | + | / | ||
| 24 | + | / | ||||
| 36 | − * | 45 | ||||
| 48 | − | 35 | ||||
| 60 | − | 25 | ||||
| 6 | 1:1000 | 12 | + | / | ||
| 24 | − * | 35 | ||||
| 36 | − | 30 | ||||
| 48 | − | 25 | ||||
| 60 | − | 20 |
| Genes | Forward (5′-3′) | Reverse (5′-3′) | Product Size | GenBank ID | References |
|---|---|---|---|---|---|
| β-actin | GCGGAATCCACGAAACCACC | CTGTCAGCGATGCCAGGGTA | 183 bp | AB270897.1 | [56] |
| MHC-Ia | TTCTCACCAACAATGACGGG | AGGGATGATCAGGGAGAAGG | 132 bp | CP003810.1 | [57] |
| MHC-II | AGTGTGGGGAAGTTTGTTGGAT | ATGGTGACTGGAGAGAGGCG | 156 bp | JN967618.1 | [57] |
| TNFα | CTCAGAGTCTATGGGAAGCAG | GCAAACACGCCAAAGAAGGT | 216 bp | NM_001279533 | [57] |
| IFN-γ | CCAACAACTCAGGCTCGCTA | TGCTCATGGTAGCGGTGTTT | 100 bp | KF294754.1 | [56] |
| CD4 | AAGAAACAGATGCGGGAGAGT | AGCAGAGGGAACGACAGAGAC | 100 bp | XM_005455473.3 | [58] |
| IL-1β | AACACTGACAGAACAACTGCGAACA | TCGCAGTTGTTCTGTCAGTGTTGTT | 124 bp | XM_019365844 | [57] |
| Group | Fish No. | Mortality Rate (%) | Survival Rate (%) | RPS (%) |
|---|---|---|---|---|
| NC | 30 | 93.3 | 6.7 | / |
| A | 30 | 90.0 | 10.0 | 3.5 |
| V1 | 30 | 53.3 | 46.7 | 42.9 b |
| V2 | 30 | 66.7 | 33.3 | 28.5 a |
| V3 | 30 | 80.0 | 20.0 | 14.3 |
| AV1 | 30 | 13.3 | 86.7 | 85.7 d |
| AV2 | 30 | 33.3 | 66.7 | 64.3 c |
| AV3 | 30 | 63.3 | 36.7 | 32.1 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, W.; Wang, Y.; Hu, H.; Wang, Q.; Bergmann, S.M.; Wang, Y.; Li, B.; Lv, Y.; Li, H.; Yin, J.; et al. Cell Culture-Derived Tilapia Lake Virus-Inactivated Vaccine Containing Montanide Adjuvant Provides High Protection against Viral Challenge for Tilapia. Vaccines 2021, 9, 86. https://doi.org/10.3390/vaccines9020086
Zeng W, Wang Y, Hu H, Wang Q, Bergmann SM, Wang Y, Li B, Lv Y, Li H, Yin J, et al. Cell Culture-Derived Tilapia Lake Virus-Inactivated Vaccine Containing Montanide Adjuvant Provides High Protection against Viral Challenge for Tilapia. Vaccines. 2021; 9(2):86. https://doi.org/10.3390/vaccines9020086
Chicago/Turabian StyleZeng, Weiwei, Yingying Wang, Huzi Hu, Qing Wang, Sven M. Bergmann, Yahui Wang, Bo Li, Yuefeng Lv, Hua Li, Jiyuan Yin, and et al. 2021. "Cell Culture-Derived Tilapia Lake Virus-Inactivated Vaccine Containing Montanide Adjuvant Provides High Protection against Viral Challenge for Tilapia" Vaccines 9, no. 2: 86. https://doi.org/10.3390/vaccines9020086
APA StyleZeng, W., Wang, Y., Hu, H., Wang, Q., Bergmann, S. M., Wang, Y., Li, B., Lv, Y., Li, H., Yin, J., & Li, Y. (2021). Cell Culture-Derived Tilapia Lake Virus-Inactivated Vaccine Containing Montanide Adjuvant Provides High Protection against Viral Challenge for Tilapia. Vaccines, 9(2), 86. https://doi.org/10.3390/vaccines9020086