
BEI Inactivated Vaccine Induces Innate and Adaptive Responses and Elicits Partial Protection upon Reassortant Betanodavirus Infection in Senegalese Sole



Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
2.2. Vaccine Inactivation
2.3. Animals and Sampling Procedure
2.4. Fish Vaccination
2.5. NNV Challenge
2.6. Specific and Neutralizing Antibody Levels
2.7. Gene Expression by Real-Time Polymerase Chain Reaction
2.8. Viral Quantification
2.9. Calculations and Statistics
3. Results
3.1. Formalin and BEI Effectively Inactivated Ss160
3.2. BEI-iSs160 Vaccine Induces the Production of Specific NNV-IgM
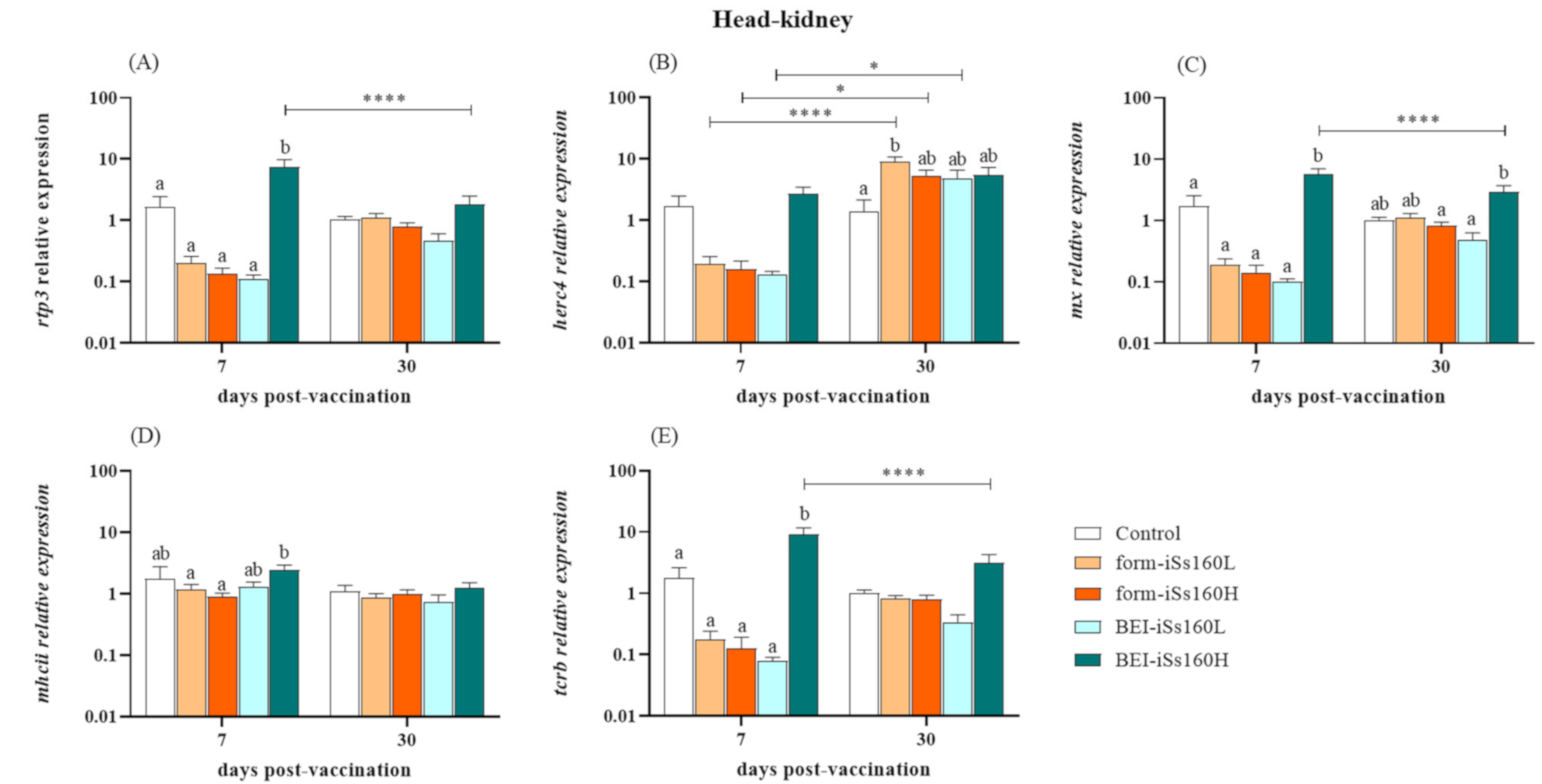
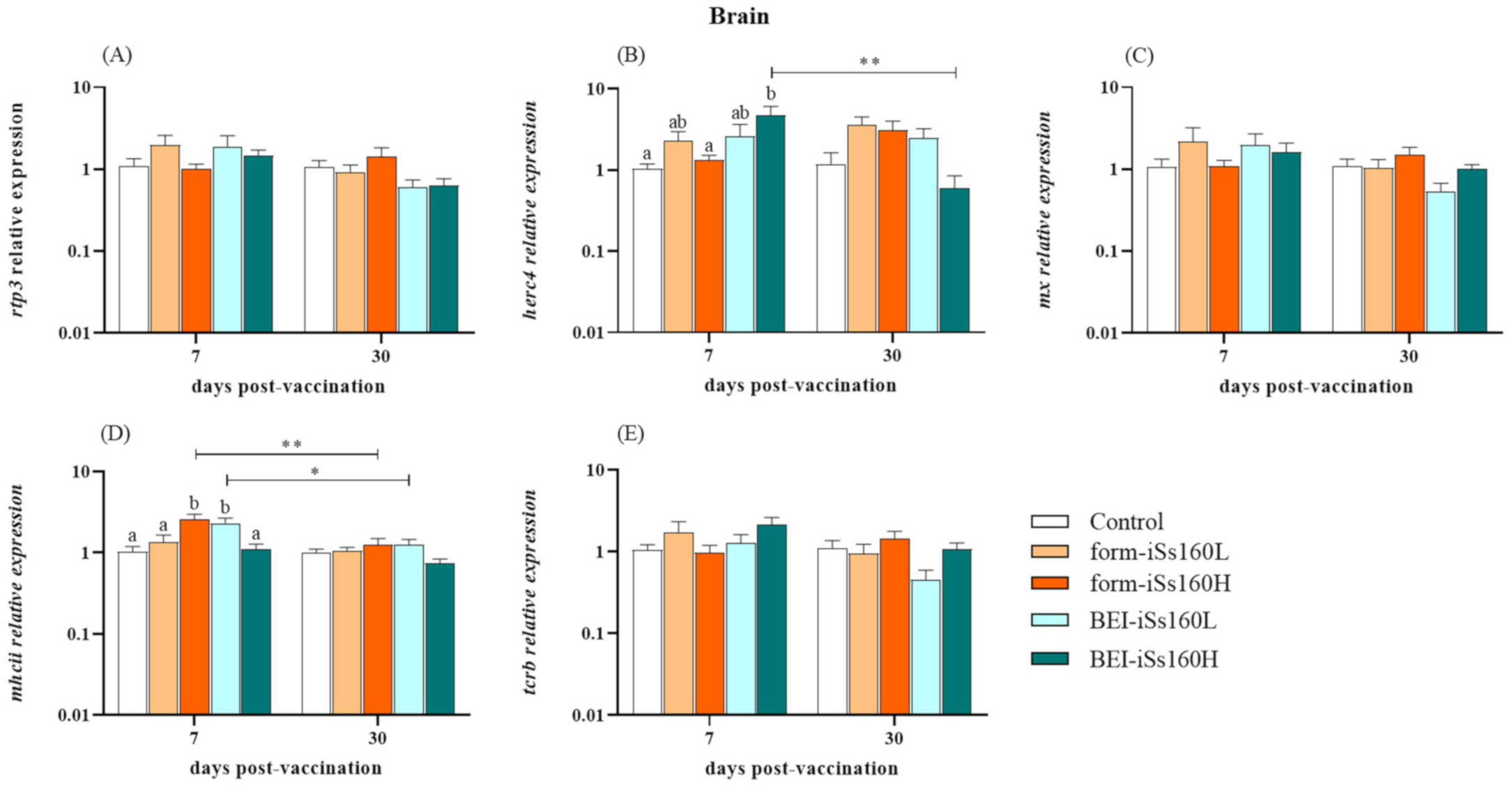
3.3. Immune-Related Gene Transcription in Vaccinated Fish
3.4. BEI-iSs160 Vaccine Improves Survival and Reduces Viral Load in Sole Brain after Challenge
3.5. NNV Infection Induces the Production of Specific NNV-IgM Similarly in All Fish
3.6. Immune-Related Genes in Head-Kidney after NNV Challenge
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture. Opportunities and Challenges; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012. [Google Scholar]
- Bandín, I.; Souto, S. Betanodavirus and VER disease: A 30-year research review. Pathogens 2020, 9, 106. [Google Scholar] [CrossRef] [Green Version]
- Olveira, J.G.; Souto, S.; Dopazo, C.P.; Thiéry, R.; Barja, J.L.; Bandín, I. Comparative analysis of both genomic segments of betanodaviruses isolated from epizootic outbreaks in farmed fish species provides evidence for genetic reassortment. J. Gen. Virol. 2009, 90, 2940–2951. [Google Scholar] [CrossRef] [PubMed]
- Volpe, E.; Gustinelli, A.; Caffara, M.; Errani, F.; Quaglio, F.; Fioravanti, M.L.; Ciulli, S. Viral nervous necrosis outbreaks caused by the RGNNV/SJNNV reassortant betanodavirus in gilthead sea bream (Sparus aurata) and European sea bass (Dicentrarchus labrax). Aquaculture 2020, 523, 735155. [Google Scholar] [CrossRef]
- Panzarin, V.; Fusaro, A.; Monne, I.; Cappellozza, E.; Patarnello, P.; Bovo, G.; Capua, I.; Holmes, E.C.; Cattoli, G. Molecular epidemiology and evolutionary dynamics of betanodavirus in southern Europe. Infect. Genet. Evol. 2012, 12, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Toffan, A.; Pascoli, F.; Pretto, T.; Panzarin, V.; Abbadi, M.; Buratin, A.; Quartesan, R.; Gijón, D.; Padros, F. Viral nervous necrosis in gilthead sea bream (Sparus aurata) caused by reassortant betanodavirus RGNNV/SJNNV: An emerging threat for Mediterranean aquaculture. Sci. Rep. 2017, 2, 46755. [Google Scholar] [CrossRef] [PubMed]
- Kai, Y.-H.; Wu, Y.-C.; Chi, S.-C. Immune gene expressions in grouper larvae (Epinephelus coioides) induced by bath and oral vaccinations with inactivated betanodavirus. Fish Shellfish Immunol. 2014, 40, 563–569. [Google Scholar] [CrossRef]
- Valero, Y.; Mokrani, D.; Chaves-Pozo, E.; Arizcun, M.; Oumouna, M.; Meseguer, J.; Esteban, M.Á.; Cuesta, A. Vaccination with UV-inactivated nodavirus partly protects European sea bass against infection, while inducing few changes in immunity. Dev. Comp. Immunol. 2018, 86, 171–179. [Google Scholar] [CrossRef]
- Chien, M.-H.; Wu, S.-Y.; Lin, C.-H. Oral immunization with cell-free self-assembly virus-like particles against orange-spotted grouper nervous necrosis virus in grouper larvae, Epinephelus coioides. Vet. Immunol. Immunopathol. 2018, 197, 69–75. [Google Scholar] [CrossRef]
- Coeurdacier, J.-L.; Laporte, F.; Pepin, J.-F. Preliminary approach to find synthetic peptides from nodavirus capsid potentially protective against sea bass viral encephalopathy and retinopathy. Fish Shellfish Immunol. 2003, 14, 435–447. [Google Scholar] [CrossRef]
- Kim, J.-O.; Kim, W.-S.; Oh, M.-J. Development of a Recombinant Protein Vaccine Based on Cell-Free Protein Synthesis for Sevenband Grouper Epinephelus septemfasciatus Against Viral Nervous Necrosis. J. Microbiol. Biotechnol. 2015, 25, 1761–1767. [Google Scholar] [CrossRef]
- González-Silvera, D.; Guardiola, F.A.; Espinosa, C.; Chaves-Pozo, E.; Esteban, M.Á.; Cuesta, A. Recombinant nodavirus vaccine produced in bacteria and administered without purification elicits humoral immunity and protects European sea bass against infection. Fish Shellfish Immunol. 2019, 88, 458–463. [Google Scholar] [CrossRef]
- Lin, C.-F.; Jiang, H.-K.; Chen, N.-C.; Wang, T.-Y.; Chen, T.-Y. Novel subunit vaccine with linear array epitope protect giant grouper against nervous necrosis virus infection. Fish Shellfish Immunol. 2018, 74, 551–558. [Google Scholar] [CrossRef]
- Valero, Y.; Awad, E.; Buonocore, F.; Arizcun, M.; Esteban, M.Á.; Meseguer, J.; Chaves-Pozo, E.; Cuesta, A. An oral chitosan DNA vaccine against nodavirus improves transcription of cell-mediated cytotoxicity and interferon genes in the European sea bass juveniles gut and survival upon infection. Dev. Comp. Immunol. 2016, 65, 64–72. [Google Scholar] [CrossRef]
- Lama, R.; Pereiro, P.; Novoa, B.; Coll, J. Sea bass immunization to downsize the betanodavirus protein displayed in the surface of inactivated repair-less bacteria. Vaccines 2019, 7, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, L.J.; Müench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Gye, H.J.; Nishizawa, T. Reducing background optical density in enzyme-linked immunosorbent assay for detecting nervous necrosis virus (NNV)-specific IgM by immobilizing fish sera. Aquaculture 2018, 485, 93–100. [Google Scholar] [CrossRef]
- Souto, S.; Olveira, J.G.; Bandín, I. Influence of temperature on Betanodavirus infection in Senegalese sole (Solea senegalensis). Vet. Microbiol. 2015, 179, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Gémez-Mata, J.; Labella, A.M.; Bandín, I.; Borrego, J.J.; García-Rosado, E. Immunogene expression analysis in betanodavirus infected-Senegalese sole using an OpenArray® platform. Gene 2021, 774, 145430. [Google Scholar] [CrossRef]
- Souto, S.; López-Jimena, B.; Alonso, M.C.; García-Rosado, E.; Bandín, I. Experimental susceptibility of European sea bass and Senegalese sole to different betanodavirus isolates. Vet. Microbiol. 2015, 177, 53–61. [Google Scholar] [CrossRef]
- Thwaite, R.; Berbel, C.; Aparicio, M.; Torrealba, D.; Pesarrodona, M.; Villaverde, A.; Borrego, J.J.; Manchado, M.; Roher, N. Nanostructured recombinant protein particles raise specific antibodies against the nodavirus NNV coat protein in sole. Fish Shellfish Immunol. 2020, 99, 578–586. [Google Scholar] [CrossRef]
- Delrue, I.; Verzele, D.; Madder, A.; Nauwynck, H.J. Inactivated virus vaccines from chemistry to prophylaxis: Merits, risks and challenges. Expert Rev. Vaccines 2014, 11, 695–719. [Google Scholar] [CrossRef] [Green Version]
- Arimoto, M.; Sato, J.; Maruyama, K.; Mimura, G.; Furusawa, I. Effect of chemical and physical treatments on the inactivation of striped jack nervous necrosis virus (SJNNV). Aquaculture 1996, 143, 15–22. [Google Scholar] [CrossRef]
- Frerichs, G.N.; Tweedie, A.; Starkey, W.G.; Richards, R.H. Temperature, pH and electrolyte sensitivity, and heat, UV and disinfectant inactivation of sea bass (Dicentrarchus labrax) neuropathy nodavirus. Aquaculture 2000, 185, 13–24. [Google Scholar] [CrossRef]
- Kai, Y.H.; Chi, S.C. Efficacies of inactivated vaccines against Betanodavirus in grouper larvae (Epinephelus coioides) by bath immunization. Vaccine 2008, 26, 1450–1457. [Google Scholar] [CrossRef]
- Yamashita, H.; Fujita, Y.; Kawakami, H.; Nakai, T. The efficacy of inactivated virus vaccine against viral nervous necrosis (VNN). Fish Pathol. 2005, 40, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.K.; Wu, Y.C.; Chi, S.C. Humoral and cytokine responses in giant groupers after vaccination and challenge with betanodavirus. Dev. Comp. Immunol. 2017, 67, 385–394. [Google Scholar] [CrossRef]
- Kai, Y.H.; Su, H.M.; Tai, K.T.; Chi, S.C. Vaccination of grouper broodfish (Epinephelus tukula) reduces the risk of vertical transmission by nervous necrosis virus. Vaccine 2010, 28, 996–1001. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.M.; Cheng, J.H.; Tu, C.; Chen, T.N.; Lin, C.T.; Chang, S.K. A bivalent inactivated vaccine of viral nervous necrosis virus and grouper iridovirus applied to grouper broodfish (Epinephelus coioides) reduces the risk of vertical transmission. Taiwan Vet. J. 2017, 43, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Núñez-Ortiz, N.; Pascoli, F.; Picchietti, S.; Buonocore, F.; Bernini, C.; Toson, M.; Scapigliati, G.; Toffan, A. A formalin-inactivated immunogen against viral encephalopathy and retinopathy (VER) disease in European sea bass (Dicentrarchus labrax): Immunological and protection effects. Vet. Res. 2016, 47, 89. [Google Scholar] [CrossRef] [Green Version]
- Pakingking, R.; Bautista, N.B.; de Jesus-Ayson, E.G.; Reyes, O. Protective immunity against viral nervous necrosis (VNN) in brown-marbled grouper (Epinephelus fuscogutattus) following vaccination with inactivated betanodavirus. Fish Shellfish Immunol. 2010, 28, 525–533. [Google Scholar] [CrossRef]
- Pakingking, R.; de Jesus-Ayson, E.G.; Reyes, O.; Brian Bautista, N. Immunization regimen in Asian sea bass (Lates calcarifer) broodfish: A practical strategy to control vertical transmission of nervous necrosis virus during seed production. Vaccine 2018, 36, 5002–5009. [Google Scholar] [CrossRef]
- Pakingking, R.; Seron, R.; dela Peña, L.; Mori, K.; Yamashita, H.; Nakai, T. Immune responses of Asian sea bass, Lates calcarifer Bloch, against an inactivated betanodavirus vaccine. J. Fish Dis. 2009, 32, 457–463. [Google Scholar] [CrossRef]
- Pascoli, F.; Guazzo, A.; Buratin, A.; Toson, M.; Buonocore, F.; Scapigliati, G.; Toffan, A. Lack of in vivo cross-protection of two different betanodavirus species RGNNV and SJNNV in European sea bass Dicentrachus Labrax. Fish Shellfish Immunol. 2019, 85, 85–89. [Google Scholar] [CrossRef]
- Yamashita, H.; Mori, K.; Kuroda, A.; Nakai, T. Neutralizing antibody levels for protection against betanodavirus infection in sevenband grouper, Epinephelus septemfasciatus (Thunberg), immunized with an inactivated virus vaccine. J. Fish Dis. 2009, 32, 767–775. [Google Scholar] [CrossRef]
- Krasnov, A.; Timmerhaus, G.; Schiøtz, B.L.; Torgersen, J.; Afanasyev, S.; Iliev, D.; Jørgensen, J.; Takle, H.; Jørgensen, S.M. Genomic survey of early responses to viruses in Atlantic salmon, Salmo salar L. Mol. Immunol. 2011, 49, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Wang, L.; Ye, B.Q.; Huang, S.; Wong, S.-M.; Yue, G.H. Characterization of a novel disease resistance gene rtp3 and its association with VNN disease resistance in Asian seabass. Fish Shellfish Immunol. 2017, 61, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Wang, L.; Kwang, J.; Yue, G.H.; Wong, S.-M. Transcriptome analysis of genes responding to NNV infection in Asian seabass epithelial cells. Fish Shellfish Immunol. 2016, 54, 342–352. [Google Scholar] [CrossRef]
- Valero, Y.; Morcillo, P.; Meseguer, J.; Buonocore, F.; Esteban, M.Á.; Chaves-Pozo, E.; Cuesta, A. Characterization of the interferon pathway in the teleost fish gonad against the vertically transmitted viral nervous necrosis virus. J. Gen. Virol. 2015, 96, 2176–2187. [Google Scholar] [CrossRef]
- Valero, Y.; Boughlala, B.; Arizcun, M.; Patel, S.; Fiksdal, I.U.; Esteban, M.Á.; De Juan, J.; Meseguer, J.; Chaves-Pozo, E.; Cuesta, A. Genes related to cell-mediated cytotoxicity and interferon response are induced in the retina of European sea bass upon intravitreal infection with nodavirus. Fish Shellfish Immunol. 2018, 74, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Carballo, C.; García-Rosado, E.; Borrego, J.J.; Alonso, M.C. SJNNV down-regulates RGNNV replication in European sea bass by the induction of the type I interferon system. Vet. Res. 2016, 47, 6. [Google Scholar] [CrossRef] [Green Version]
- Poisa-Beiro, L.; Dios, S.; Montes, A.; Aranguren, R.; Figueras, A.; Novoa, B. Nodavirus increases the expression of Mx and inflammatory cytokines in fish brain. Mol. Immunol. 2008, 45, 218–225. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-Torres, D.; Podadera, A.M.; Alonso, M.C.; Bandín, I.; Béjar, J.; García-Rosado, E. Molecular characterization and expression analyses of the Solea senegalensis interferon-stimulated gene 15 (isg15) following NNV infections. Fish Shellfish Immunol. 2017, 66, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Øvergård, A.C.; Nerland, A.H.; Fiksdal, I.U.; Patel, S. Atlantic halibut experimentally infected with nodavirus shows increased levels of T-cell marker and IFNgamma transcripts. Dev. Comp. Immunol. 2012, 37, 139–150. [Google Scholar] [CrossRef] [PubMed]
- González-Fernández, C.; Esteban, M.Á.; Cuesta, A. Molecular characterization of the T cell costimulatory receptors CD28 and CTLA4 in the European sea bass. Fish Shellfish Immunol. 2021, 109, 106–115. [Google Scholar] [CrossRef] [PubMed]
- López-Muñoz, A.; Sepulcre, M.P.; García-Moreno, D.; Fuentes, I.; Béjar, J.; Manchado, M.; Álvarez, M.C.; Meseguer, J.; Mulero, V. Viral nervous necrosis virus persistently replicates in the central nervous system of asymptomatic gilthead seabream and promotes a transient inflammatory response followed by the infiltration of IgM+ B lymphocytes. Dev. Comp. Immunol. 2012, 37, 429–437. [Google Scholar] [CrossRef]
- Øvergård, A.C.; Patel, S.; Nøstbakken, O.J.; Nerland, A.H. Atlantic halibut (Hippoglossus hippoglossus L.) T-cell and cytokine response after vaccination and challenge with nodavirus. Vaccine 2013, 31, 2395–2402. [Google Scholar] [CrossRef]
- Rise, M.L.; Hall, J.R.; Rise, M.; Hori, T.S.; Browne, M.J.; Gamperl, A.K.; Hubert, S.; Kimball, J.; Bowman, S.; Johnson, S.C. Impact of asymptomatic nodavirus carrier state and intraperitoneal viral mimic injection on brain transcript expression in Atlantic cod (Gadus morhua). Physiol. Genom. 2010, 42, 266–280. [Google Scholar] [CrossRef]
- Eslamloo, K.; Xue, X.; Booman, M.; Smith, N.C.; Rise, M.L. Transcriptome profiling of the antiviral immune response in Atlantic cod macrophages. Dev. Comp. Immunol. 2016, 63, 187–205. [Google Scholar] [CrossRef]
- Tanaka, S.; Mori, K.; Arimoto, M.; Iwamoto, T.; Nakai, T. Protective immunity of sevenband grouper, Epinephelus septemfasciatus Thunberg, against experimental viral nervous necrosis. J. Fish Dis. 2001, 24, 15–22. [Google Scholar] [CrossRef]
- Qin, Y.; Wang, Y.; Liu, J.; Lu, Y.; Liu, X. Red-grouper nervous necrosis virus B2 protein negatively regulates fish interferon response by suppressing host transcription directed by RNA polymerase II. Aquaculture 2021, 536, 736488. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Gene Name | Accession Number, UniGen Name or Reference | Sequence (5′–3′) | ||
|---|---|---|---|---|---|
| Nodavirus | RNA-dependent RNA polymerase | RNA1 | FJ803911 | F | TCCAAAAGAAAGAAGCATAC |
| R | TGGCATGTACCACGGAAC | ||||
| Senegalese sole | Receptor transporter protein | rtp3 | [20] | F | GACGCCCCAATGGTGGAT |
| R | CCAGATTCTTCATGAGGATGGTGAT | ||||
| E3 ubiquitin-protein ligase | herc4 | [20] | F | GCCAAAACACTGGCATGGTT | |
| R | AACGCCAAACAGGAAGTACCT | ||||
| Interferon-induced GTP-binding protein Mx | mx | [20] | F | CCTCTCTCCTTCAGGATCCTCCTCCTGTGC | |
| R | CAAAACAAGAAACTATCTGCCTGGTGGTTC | ||||
| Major histocompatibility complex II | mhcii | [20] | F | CGCTGATGAAAATGATCCACCTTCT | |
| R | ACCAGTCACATGACAGATCAGAGT | ||||
| T-cell receptor beta chain | tcrb | solea_v4.1_unigene681812 * | F | CAGGAGGCACAGCTATGAAA | |
| R | TCTCCACCCAAATCTCCAAA | ||||
| Beta actin | actb | DQ485686 | F | GACGACATGGAGAAGATC | |
| R | GGTGTTGAAGGTCTCAAA | ||||
| Chemical Compound | Concentration | Time | CPE under Each Blind Passage | Titer after Virus Inactivation(Log10 TCID50/mL) | ||||
|---|---|---|---|---|---|---|---|---|
| % | mM | Days | Hours | 1 | 2 | 3 | ||
| Formalin | 0.2 | 7 | − | − | + | nt | ||
| 9 | − | − | − | <1.0 | ||||
| BEI | 0.1 | 1 | + | nt | nt | nt | ||
| 24 | + | nt | nt | nt | ||||
| 48 | + | nt | nt | nt | ||||
| 72 | + | nt | nt | nt | ||||
| 1 | 1 | + | + | nt | nt | |||
| 24 | + | + | nt | nt | ||||
| 48 | − | − | + | nt | ||||
| 72 | − | − | − | <1.0 | ||||
| 4 | 1 | − | − | − | <1.0 | |||
| 24 | − | − | − | <1.0 | ||||
| 48 | − | − | − | <1.0 | ||||
| 72 | − | − | − | <1.0 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valero, Y.; Olveira, J.G.; López-Vázquez, C.; Dopazo, C.P.; Bandín, I. BEI Inactivated Vaccine Induces Innate and Adaptive Responses and Elicits Partial Protection upon Reassortant Betanodavirus Infection in Senegalese Sole. Vaccines 2021, 9, 458. https://doi.org/10.3390/vaccines9050458
Valero Y, Olveira JG, López-Vázquez C, Dopazo CP, Bandín I. BEI Inactivated Vaccine Induces Innate and Adaptive Responses and Elicits Partial Protection upon Reassortant Betanodavirus Infection in Senegalese Sole. Vaccines. 2021; 9(5):458. https://doi.org/10.3390/vaccines9050458
Chicago/Turabian StyleValero, Yulema, José G. Olveira, Carmen López-Vázquez, Carlos P. Dopazo, and Isabel Bandín. 2021. "BEI Inactivated Vaccine Induces Innate and Adaptive Responses and Elicits Partial Protection upon Reassortant Betanodavirus Infection in Senegalese Sole" Vaccines 9, no. 5: 458. https://doi.org/10.3390/vaccines9050458
APA StyleValero, Y., Olveira, J. G., López-Vázquez, C., Dopazo, C. P., & Bandín, I. (2021). BEI Inactivated Vaccine Induces Innate and Adaptive Responses and Elicits Partial Protection upon Reassortant Betanodavirus Infection in Senegalese Sole. Vaccines, 9(5), 458. https://doi.org/10.3390/vaccines9050458