
Mutation E48K in PB1 Polymerase Subunit Improves Stability of a Candidate Live Attenuated Influenza B Virus Vaccine

 ,
,  , ,
, , 

Abstract
:1. Introduction
2. Material and Methods
2.1. Tissue Culture Cells and Embryonated Chicken Eggs
2.2. Cloning and Site-Directed Mutagenesis of PB1 Segment
2.3. B/Bris PB1att 4M Strain Generation by Reverse Genetics
2.4. Sequencing
2.5. Genetic Stability of Viruses
2.6. Viral Growth Kinetics In Vitro
2.7. Structural Analysis of the PB1 Segment
2.8. Animal Use Compliance
2.9. Vaccination and Vaccine Safety Study Design
2.10. Virus Challenge Study Design
2.11. Hemagglutination Inhibition Assay
2.12. Statistical Analysis
3. Results
3.1. Mutations E48K and K391E Remain Stable in Combination after Serial Passages in MDCKs and SPF Eggs
3.2. Stability of B/Bris PB1att 4M Strain Was Confirmed after Serial Passages
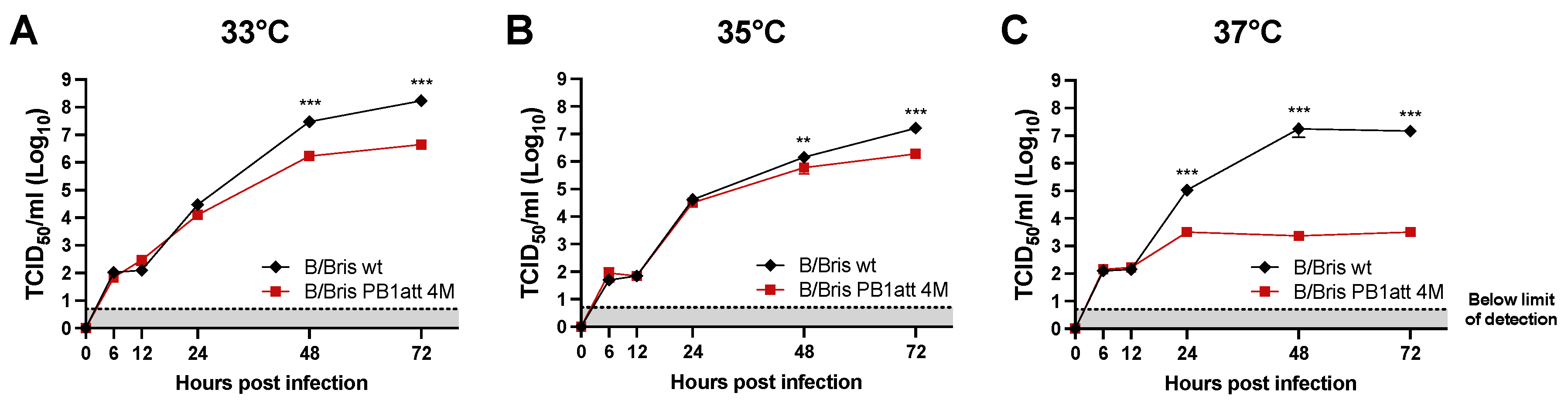
3.3. B/Bris PB1att 4M Virus Shows a Temperature-Sensitive Phenotype In Vitro
3.4. Structural Analysis of PB1 from B/Bris PB1att 4M Suggests Molecular Interactions between Amino Acid Residues at Positions 48 and 391
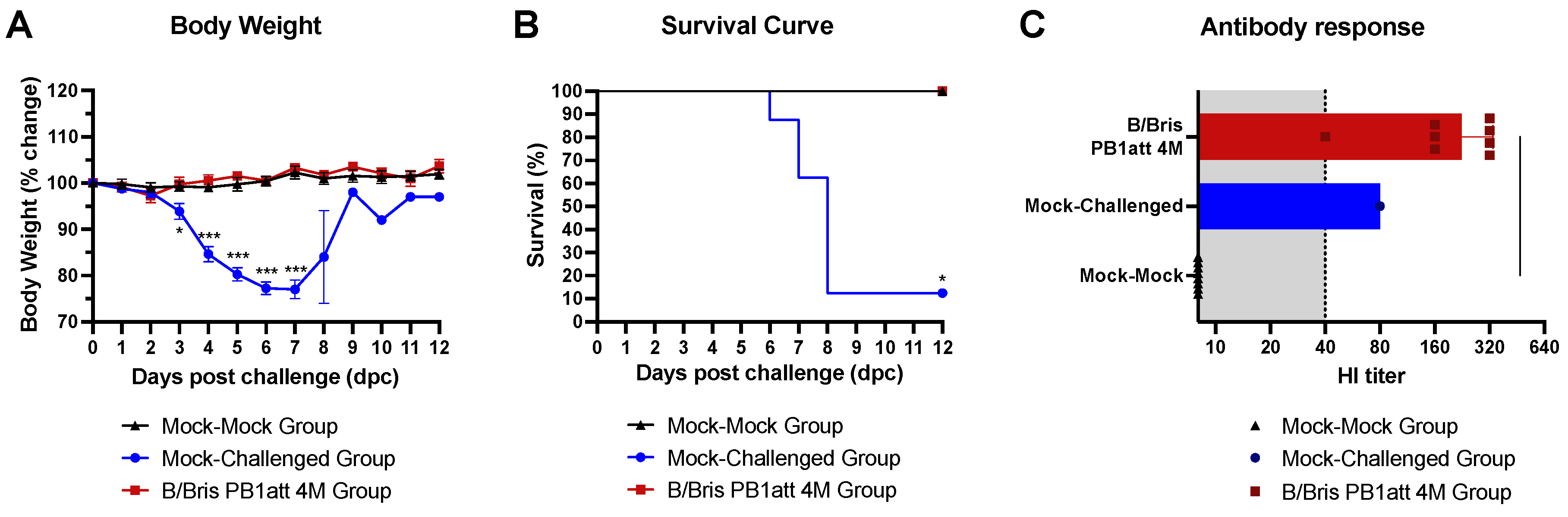
3.5. B/Bris PB1att 4M Is Safe and Immunogenic as a Potential Vaccine Candidate
3.6. B/Bris PB1att 4M Vaccine Candidate Successfully Protects Mice against Lethal Challenge
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ritchey, M.B.; Palese, P.; Kilbourne, E.D. RNAs of influenza A, B, and C viruses. J. Virol. 1976, 18, 738–744. [Google Scholar] [CrossRef] [Green Version]
- Bouvier, N.M.; Palese, P. The biology of influenza viruses. Vaccine 2008, 26 (Suppl. 4), D49–D53. [Google Scholar] [CrossRef] [Green Version]
- Hatta, M.; Goto, H.; Kawaoka, Y. Influenza B virus requires BM2 protein for replication. J. Virol. 2004, 78, 5576–5583. [Google Scholar] [CrossRef] [Green Version]
- Owusu, D.; Hand, J.; Tenforde, M.W.; Feldstein, L.R.; DaSilva, J.; Barnes, J.; Lee, G.; Tran, J.; Sokol, T.; Fry, A.M.; et al. Early season pediatric Influenza B/Victoria virus infections associated with a recently emerged virus subclade–Louisiana, 2019. Am. J. Transplant. 2020, 20, 606–609. [Google Scholar] [CrossRef]
- Caini, S.; Huang, Q.S.; Ciblak, M.A.; Kusznierz, G.; Owen, R.; Wangchuk, S.; Henriques, C.M.; Njouom, R.; Fasce, R.A.; Yu, H.; et al. Epidemiological and virological characteristics of Influenza B: Results of the Global Influenza B Study. Influenza Other Respir. Viruses 2015, 9 (Suppl. 1), 3–12. [Google Scholar] [CrossRef] [PubMed]
- Paul Glezen, W.; Schmier, J.K.; Kuehn, C.M.; Ryan, K.J.; Oxford, J. The burden of Influenza B: A structured literature review. Am. J. Public Health 2013, 103, e43–e51. [Google Scholar] [CrossRef] [PubMed]
- Belshe, R.B. The need for quadrivalent vaccine against seasonal influenza. Vaccine 2010, 28 (Suppl. 4), D45–D53. [Google Scholar] [CrossRef] [PubMed]
- Shaw, M.W.; Xu, X.; Li, Y.; Normand, S.; Ueki, R.T.; Kunimoto, G.Y.; Hall, H.; Klimov, A.; Cox, N.J.; Subbarao, K. Reappearance and global spread of variants of Influenza B/Victoria/2/87 lineage viruses in the 2000–2001 and 2001–2002 seasons. Virology 2002, 303, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Langat, P.; Raghwani, J.; Dudas, G.; Bowden, T.A.; Edwards, S.; Gall, A.; Bedford, T.; Rambaut, A.; Daniels, R.S.; Russell, C.A.; et al. Genome-wide evolutionary dynamics of Influenza B viruses on a global scale. PLoS Pathog. 2017, 13, e1006749. [Google Scholar] [CrossRef] [Green Version]
- Ermler, M.E.; Kirkpatrick, E.; Sun, W.; Hai, R.; Amanat, F.; Chromikova, V.; Palese, P.; Krammer, F. Chimeric Hemagglutinin Constructs Induce Broad Protection against Influenza B Virus Challenge in the Mouse Model. J. Virol. 2017, 91, e00286-17. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, E.; Liang, X.; Ingallinella, P.; Finotto, M.; Chastain, M.A.; Fan, J.; Fu, T.M.; Song, H.C.; Horton, M.S.; Freed, D.C.; et al. Universal Influenza B vaccine based on the maturational cleavage site of the hemagglutinin precursor. J. Virol. 2005, 79, 7380–7388. [Google Scholar] [CrossRef] [Green Version]
- Reed, C.; Meltzer, M.I.; Finelli, L.; Fiore, A. Public health impact of including two lineages of Influenza B in a quadrivalent seasonal influenza vaccine. Vaccine 2012, 30, 1993–1998. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Prevention and control of seasonal influenza with vaccines. Recommendations of the Advisory Committee on Immunization Practices–United States, 2013–2014. MMWR Recomm. Rep. 2013, 62, 1–43. [Google Scholar]
- Jin, H.; Chen, Z. Production of live attenuated influenza vaccines against seasonal and potential pandemic influenza viruses. Curr. Opin. Virol. 2014, 6, 34–39. [Google Scholar] [CrossRef]
- Shannon, I.; White, C.L.; Nayak, J.L. Understanding Immunity in Children Vaccinated With Live Attenuated Influenza Vaccine. J. Pediatric Infect. Dis. Soc. 2019, 9 (Suppl. 1), S10–S14. [Google Scholar] [CrossRef]
- Ashkenazi, S.; Vertruyen, A.; Aristegui, J.; Esposito, S.; McKeith, D.D.; Klemola, T.; Biolek, J.; Kuhr, J.; Bujnowski, T.; Desgrandchamps, D.; et al. Superior relative efficacy of live attenuated influenza vaccine compared with inactivated influenza vaccine in young children with recurrent respiratory tract infections. Pediatr. Infect. Dis. J. 2006, 25, 870–879. [Google Scholar] [CrossRef] [Green Version]
- Nohynek, H.; Baum, U.; Syrjanen, R.; Ikonen, N.; Sundman, J.; Jokinen, J. Effectiveness of the live attenuated and the inactivated influenza vaccine in two-year-olds–a nationwide cohort study Finland, influenza season 2015/16. Eurosurveillance 2016, 21, 30346. [Google Scholar] [CrossRef] [PubMed]
- Grohskopf, L.A.; Sokolow, L.Z.; Broder, K.R.; Olsen, S.J.; Karron, R.A.; Jernigan, D.B.; Bresee, J.S. Prevention and Control of Seasonal Influenza with Vaccines. MMWR Recomm. Rep. 2016, 65, 1–54. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Nieto, G.R.; Perez, D.R. A new generation of modified live-attenuated avian influenza viruses using a two-strategy combination as potential vaccine candidates. J. Virol. 2007, 81, 9238–9248. [Google Scholar] [CrossRef] [Green Version]
- Solorzano, A.; Ye, J.; Perez, D.R. Alternative live-attenuated influenza vaccines based on modifications in the polymerase genes protect against epidemic and pandemic flu. J. Virol. 2010, 84, 4587–4596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pena, L.; Vincent, A.L.; Ye, J.; Ciacci-Zanella, J.R.; Angel, M.; Lorusso, A.; Gauger, P.C.; Janke, B.H.; Loving, C.L.; Perez, D.R. Modifications in the polymerase genes of a swine-like triple-reassortant influenza virus to generate live attenuated vaccines against 2009 pandemic H1N1 viruses. J. Virol. 2011, 85, 456–469. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.J.S.; Finch, C.; Sutton, T.; Obadan, A.; Aguirre, I.; Wan, Z.; Lopez, D.; Geiger, G.; Gonzalez-Reiche, A.S.; Ferreri, L.; et al. Development of an Alternative Modified Live Influenza B Virus Vaccine. J. Virol. 2017, 91, e00056-17. [Google Scholar] [CrossRef] [Green Version]
- Reed, L.J.; Muench, H. A simple method for estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Mena, I.; Nelson, M.I.; Quezada-Monroy, F.; Dutta, J.; Cortes-Fernandez, R.; Lara-Puente, J.H.; Castro-Peralta, F.; Cunha, L.F.; Trovao, N.S.; Lozano-Dubernard, B.; et al. Origins of the 2009 H1N1 influenza pandemic in swine in Mexico. eLife 2016, 5, e16777. [Google Scholar] [CrossRef]
- Martin, S. Time to stake a claim to nursing functions. Nurse Pract. 1992, 17, 10. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Manual on Animal Influenza Diagnosis and Surveillance. Available online: http://www.who.int/csr/resources/publications/influenza/en/whocdscsrncs20025rev.pdf (accessed on 11 June 2013).
- Zhao, X.; Fang, V.J.; Ohmit, S.E.; Monto, A.S.; Cook, A.R.; Cowling, B.J. Quantifying protection against influenza virus infection measured by hemagglutination-inhibition assays in vaccine trials. Epidemiology 2016, 27, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Zengel, J.R.; Suguitan, A.L., Jr.; Xu, Q.; Wang, W.; Lin, J.; Jin, H. Evaluation of the humoral and cellular immune responses elicited by the live attenuated and inactivated influenza vaccines and their roles in heterologous protection in ferrets. J. Infect. Dis. 2013, 208, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Loving, C.L.; Vincent, A.L.; Pena, L.; Perez, D.R. Heightened adaptive immune responses following vaccination with a temperature-sensitive, live-attenuated influenza virus compared to adjuvanted, whole-inactivated virus in pigs. Vaccine 2012, 30, 5830–5838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loving, C.L.; Lager, K.M.; Vincent, A.L.; Brockmeier, S.L.; Gauger, P.C.; Anderson, T.K.; Kitikoon, P.; Perez, D.R.; Kehrli, M.E., Jr. Efficacy in pigs of inactivated and live attenuated influenza virus vaccines against infection and transmission of an emerging H3N2 similar to the 2011-2012 H3N2v. J. Virol. 2013, 87, 9895–9903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pena, L.; Sutton, T.; Chockalingam, A.; Kumar, S.; Angel, M.; Shao, H.; Chen, H.; Li, W.; Perez, D.R. Influenza viruses with rearranged genomes as live-attenuated vaccines. J. Virol. 2013, 87, 5118–5127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Z.; Cardenas Garcia, S.; Liu, J.; Santos, J.; Carnaccini, S.; Geiger, G.; Ferreri, L.; Rajao, D.; Perez, D.R. Alternative Strategy for a Quadrivalent Live Attenuated Influenza Virus Vaccine. J. Virol. 2018, 92, e01025-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szamecz, B.; Boross, G.; Kalapis, D.; Kovacs, K.; Fekete, G.; Farkas, Z.; Lazar, V.; Hrtyan, M.; Kemmeren, P.; Groot Koerkamp, M.J.; et al. The genomic landscape of compensatory evolution. PLoS Biol. 2014, 12, e1001935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, B.H.; Poon, A.F.; Whitlock, M.C. Compensatory mutations are repeatable and clustered within proteins. Proc. Biol. Sci. 2009, 276, 1823–1827. [Google Scholar] [CrossRef] [Green Version]
- Soskine, M.; Tawfik, D.S. Mutational effects and the evolution of new protein functions. Nat. Rev. Genet. 2010, 11, 572–582. [Google Scholar] [CrossRef]
- Kajikawa, T.; Kataoka, K.; Sakurai, T. Modifications on the hydrogen bond network by mutations of Escherichia coli copper efflux oxidase affect the process of proton transfer to dioxygen leading to alterations of enzymatic activities. Biochem. Biophys. Res. Commun. 2012, 422, 152–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, H.J.; Jang, D.S.; Kim, Y.G.; Hong, B.H.; Woo, J.S.; Kim, K.T.; Choi, K.Y. Rescue of deleterious mutations by the compensatory Y30F mutation in ketosteroid isomerase. Mol. Cells 2013, 36, 39–46. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | SPF Egg Passage | |
| sE1 | sE4 | |
| B/Bris PB1 3M | K391E, E580G, S660A | E48K, K391E, E580G, S660A, G161C |
| B/Bris PB1att 4M | E48K, K391E, E580G, S660A | E48K, K391E, E580G, S660A |
| Strain | MDCK Passage | |
| sP1 | sP4 | |
| B/Bris PB1 3M | K391E, E580G, S660A | K391E, E580G, S660A |
| B/Bris PB1att 4M | E48K, K391E, E580G, S660A | E48K, K391E, E580G, S660A |
| Segments | Predicted Mutations | Viral Stock Amplified in MDCK Cells (P1) | Serially Passaged in MDCKS (sP4) | Serially Passaged in SPF Eggs (sE4) |
|---|---|---|---|---|
| PB2 | None | None | None | None |
| PB1 | E48K (g163a) K391E (a1192g) E580G (a1760g) S660A (t1999g) | E48K (g163a) K391E (a1192g) E580G (a1760g) S660A (t1999g) Stop a,s (g2280a) | E48K (g163a) K391E (a1192g) E580G (a1760g) S660A (t1999g) Stop a,s (g2280a) | E48K (g163a) K391E (a1192g) E580G (a1760g) S660A (t1999g) Stop a,s (g2280a) |
| PA | None | None | None | None |
| HA | None | None | None | None |
| NP | None | None | None | None |
| NA | None | None | None | None |
| NB | None | None | None | None |
| BM1 | None | None | None | None |
| BM2 | None | None | None | None |
| NS1 | None | None | None | None |
| NS2/NEP | None | None | None | None |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, J.; Cardenas-Garcia, S.; Santos, J.J.S.; Ferreri, L.M.; Cáceres, C.J.; Geiger, G.; Perez, D.R.; Rajao, D.S. Mutation E48K in PB1 Polymerase Subunit Improves Stability of a Candidate Live Attenuated Influenza B Virus Vaccine. Vaccines 2021, 9, 800. https://doi.org/10.3390/vaccines9070800
Mo J, Cardenas-Garcia S, Santos JJS, Ferreri LM, Cáceres CJ, Geiger G, Perez DR, Rajao DS. Mutation E48K in PB1 Polymerase Subunit Improves Stability of a Candidate Live Attenuated Influenza B Virus Vaccine. Vaccines. 2021; 9(7):800. https://doi.org/10.3390/vaccines9070800
Chicago/Turabian StyleMo, Jongsuk, Stivalis Cardenas-Garcia, Jefferson J. S. Santos, Lucas M. Ferreri, C. Joaquín Cáceres, Ginger Geiger, Daniel R. Perez, and Daniela S. Rajao. 2021. "Mutation E48K in PB1 Polymerase Subunit Improves Stability of a Candidate Live Attenuated Influenza B Virus Vaccine" Vaccines 9, no. 7: 800. https://doi.org/10.3390/vaccines9070800
APA StyleMo, J., Cardenas-Garcia, S., Santos, J. J. S., Ferreri, L. M., Cáceres, C. J., Geiger, G., Perez, D. R., & Rajao, D. S. (2021). Mutation E48K in PB1 Polymerase Subunit Improves Stability of a Candidate Live Attenuated Influenza B Virus Vaccine. Vaccines, 9(7), 800. https://doi.org/10.3390/vaccines9070800