
A Stochastic Spatiotemporal Model of Rat Ventricular Myocyte Calcium Dynamics Demonstrated Necessary Features for Calcium Wave Propagation




Abstract
:1. Introduction
2. Materials and Methods
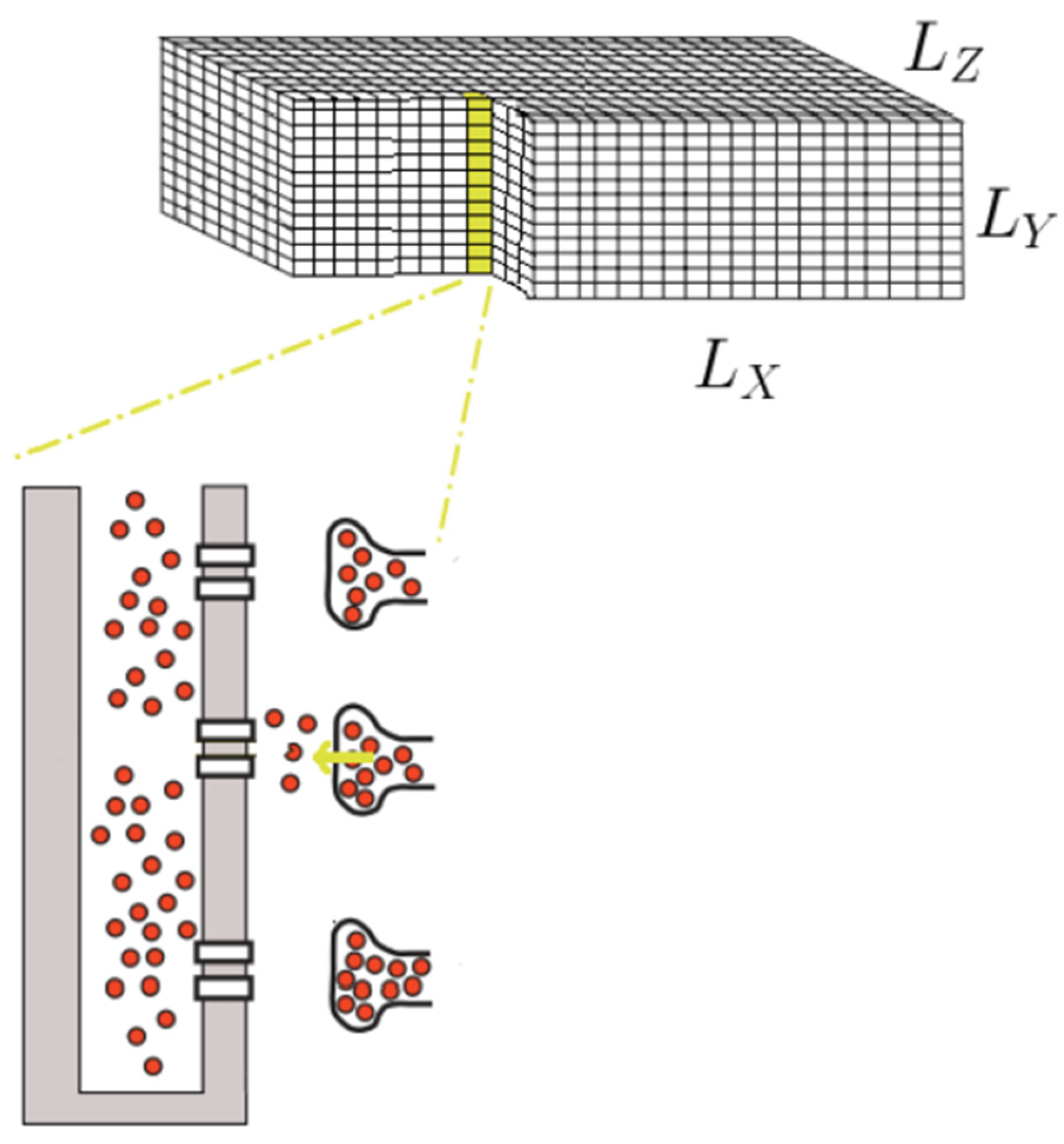
2.1. Model Development
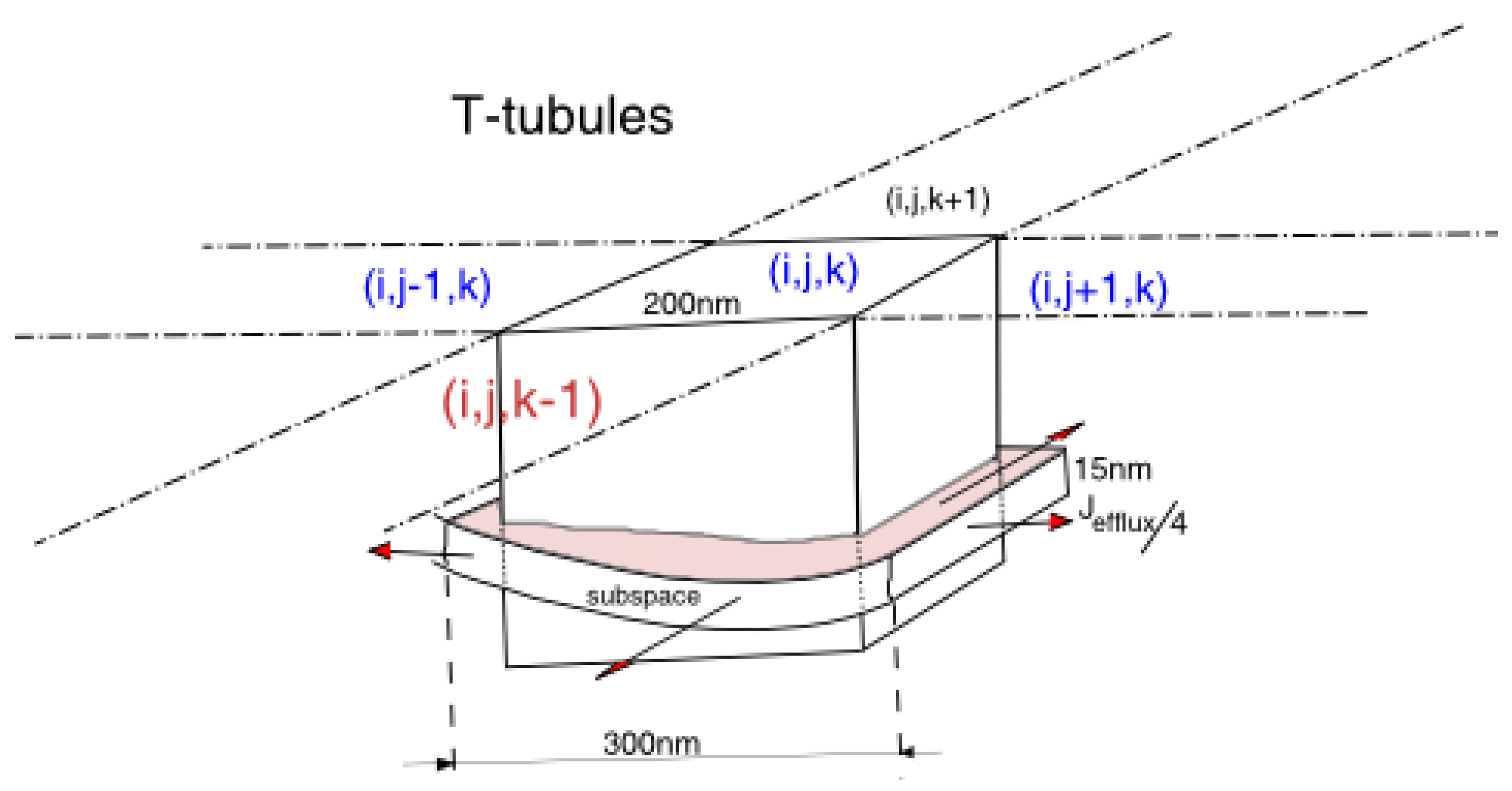
2.2. Sarcolemma(SL) and T-Tubule Membrane
2.3. Calcium Release Site
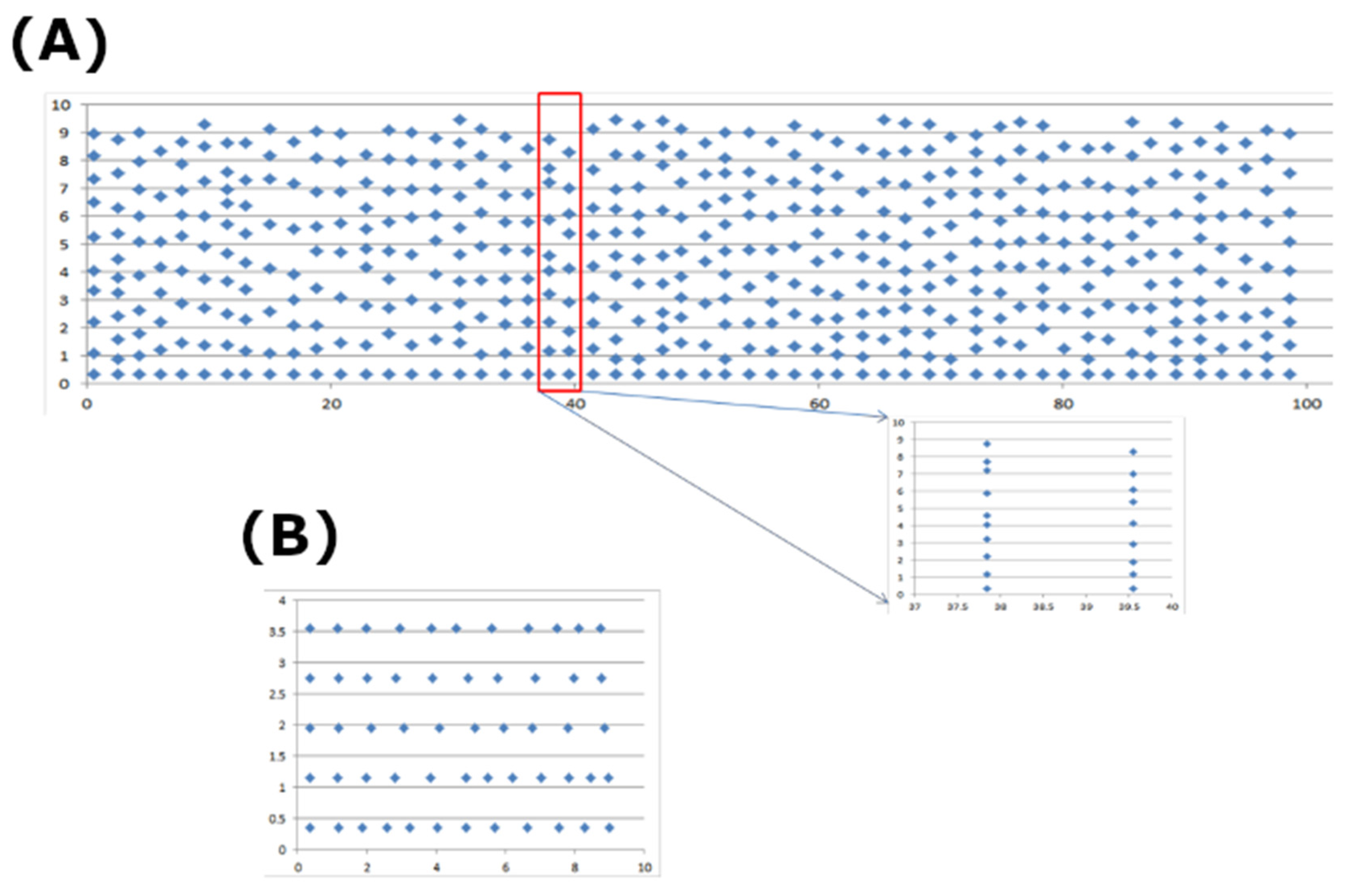
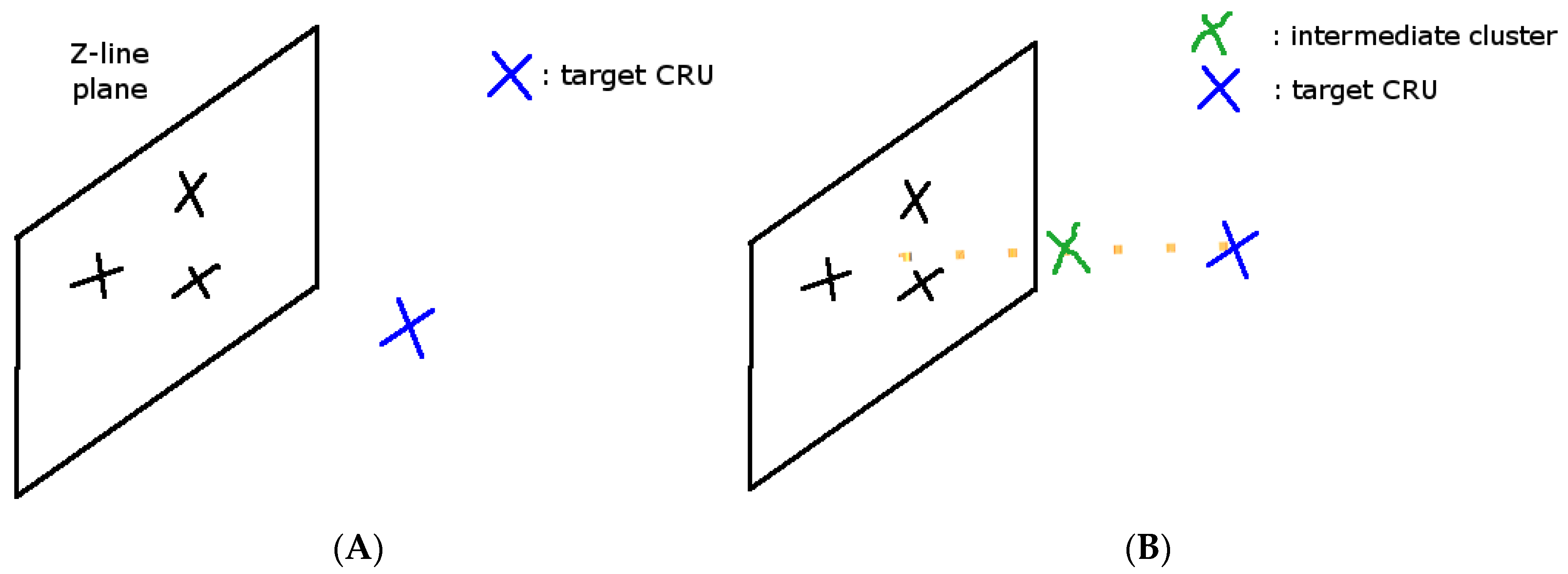
2.4. Spatial Placement of CRUs
2.5. Spatial Placement of Na+/Ca2+ Exchangers, SERCA Pump, SR and SL Buffers
2.6. Diffusion of Ions
2.7. Model Formulation in the Spatial Cell
2.8. Computational Methods
- Bit 0–3: index to the array that tells NCX distribution
- Bit 4–7: index to the array that tells SERCA distribution
- Bit 8–11: index to the array that tells SR buffer distribution
- Bit 12–15: index to the array that tells Troponin-C distribution
- Bit 16–19: index to the array that tells the grid-type (MEMBRANE, INNER-GRIDPOINT, OUTER-GRIDPOINT, NUCLEUS, MITO). Currently, we only use MEMBRANE (both SL and T-tubule) and INNER-GRIDPOINT and OUTER-GRIDPOINT (stencil grid point).
- Bit 20–31: reserved (for future use)
3. Results
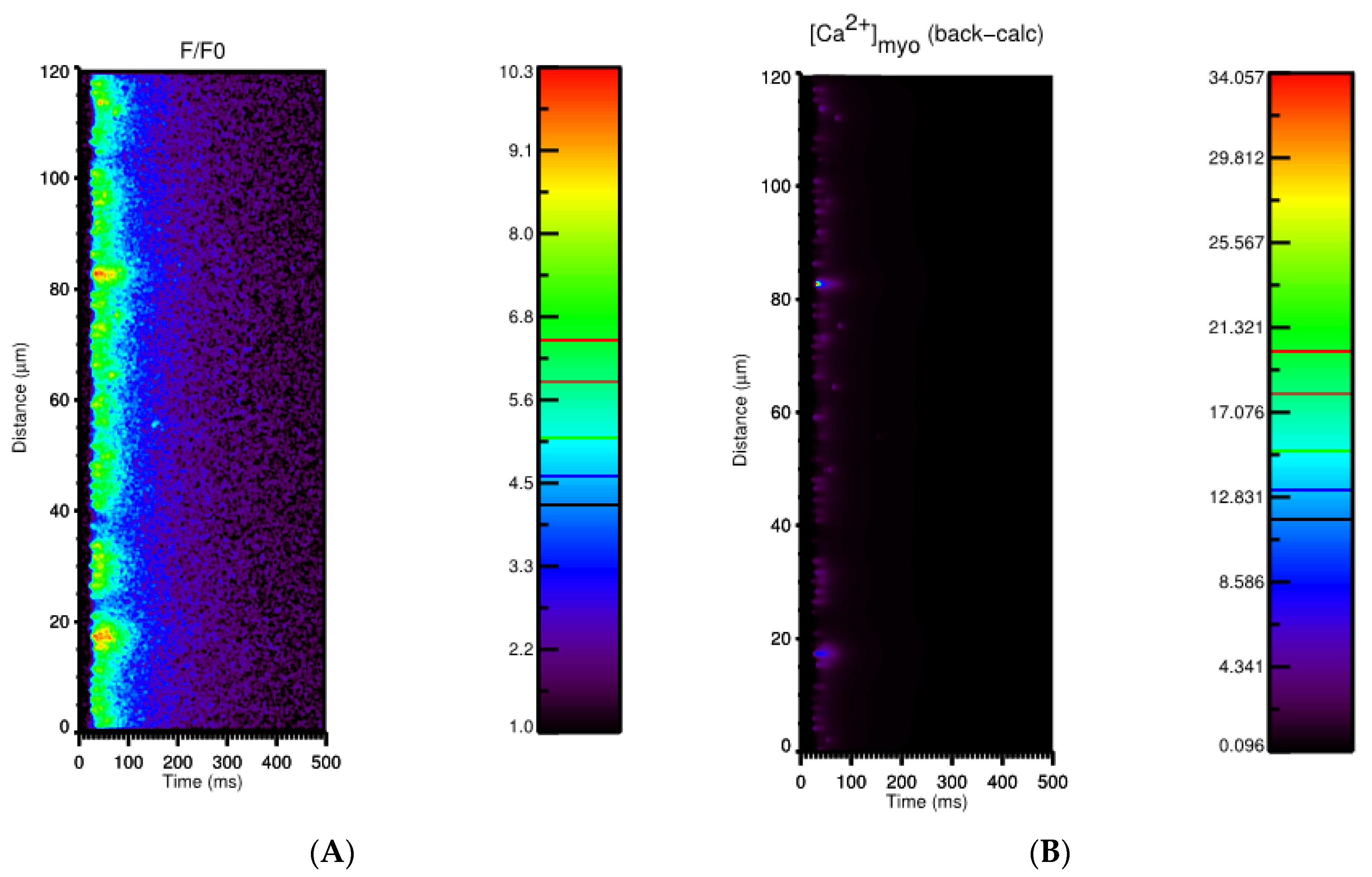
3.1. Ca2+ Transient
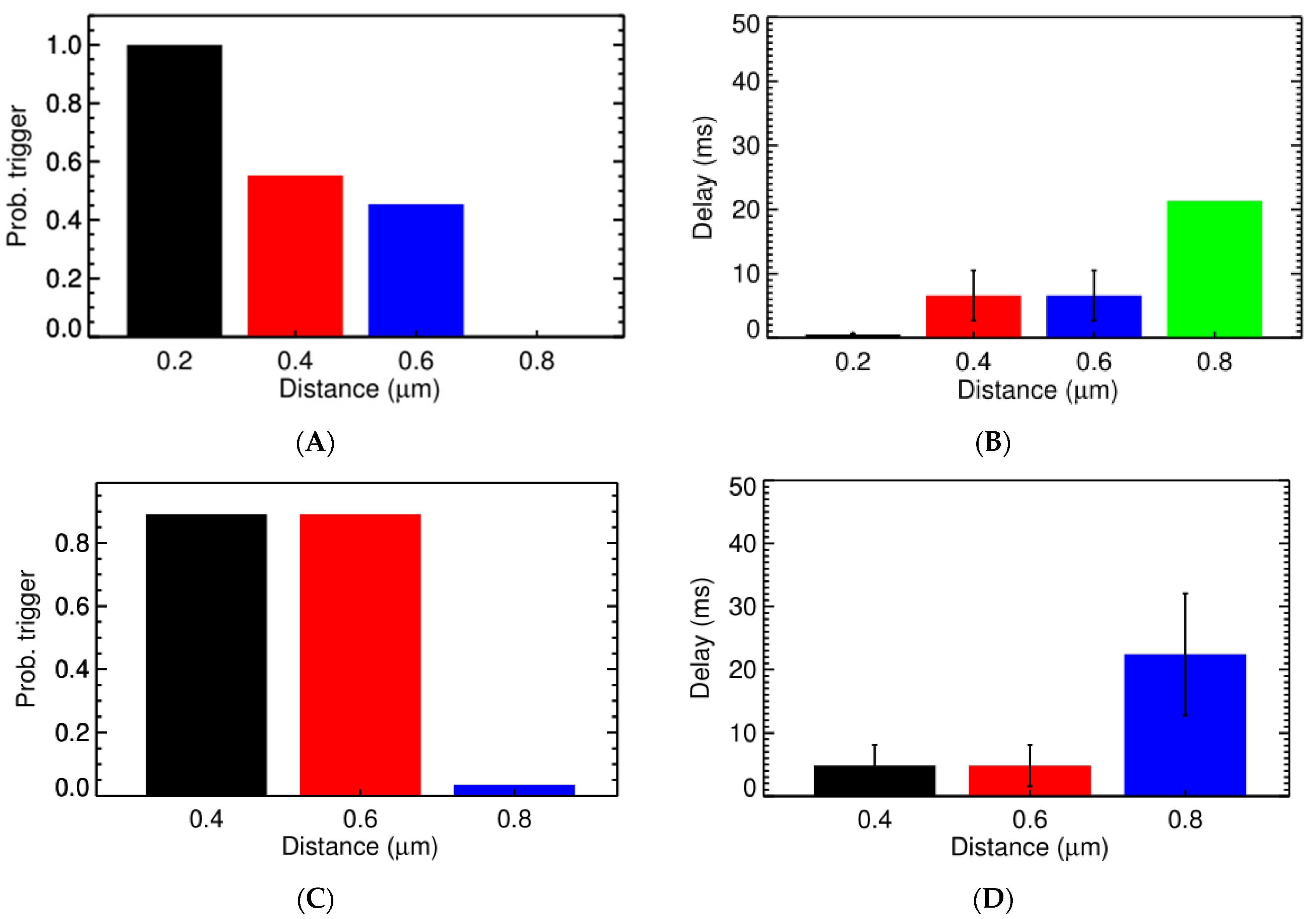
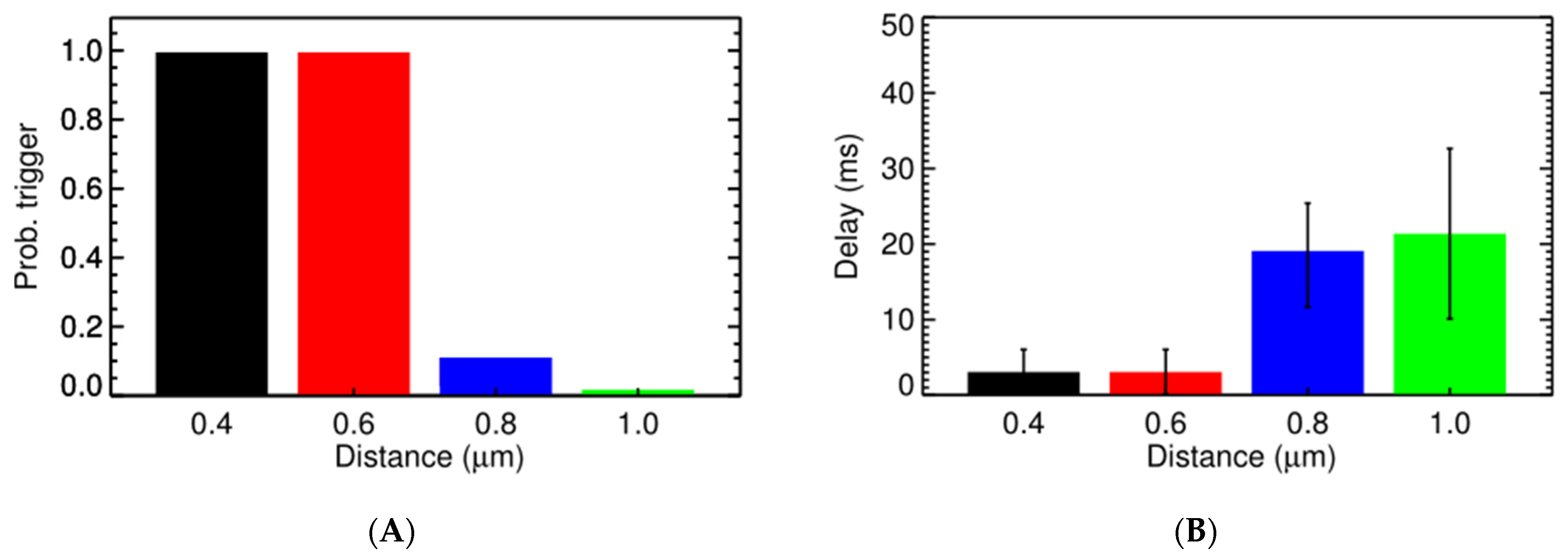
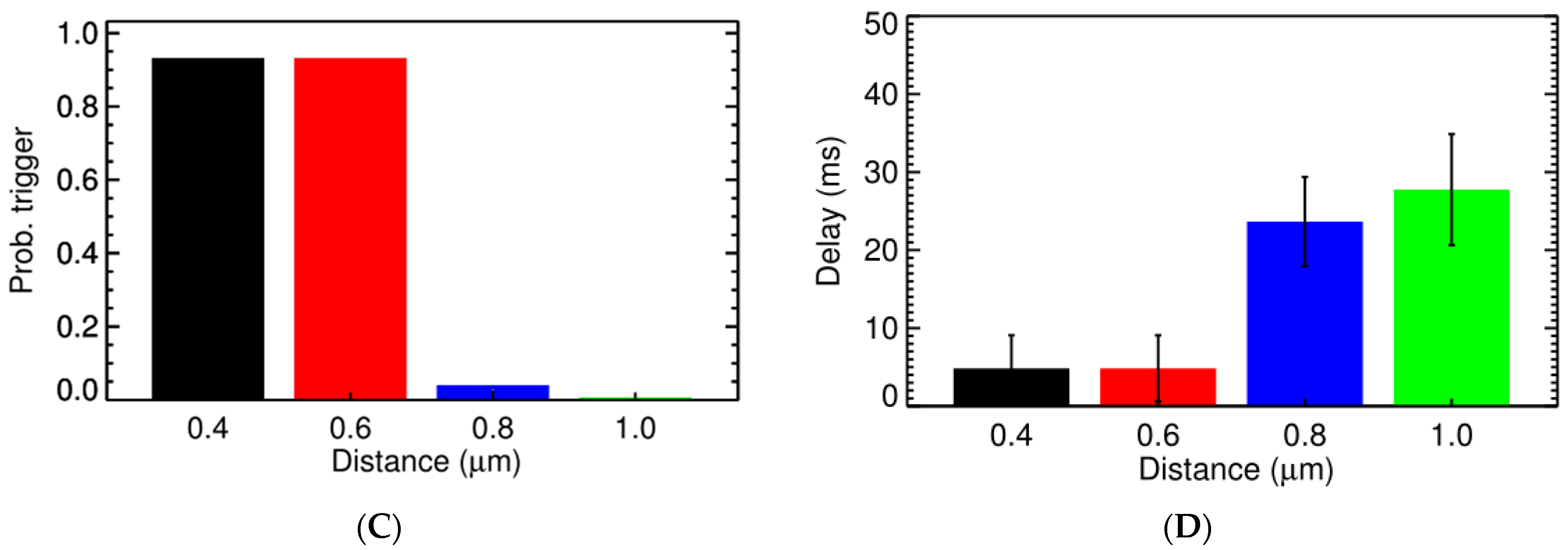
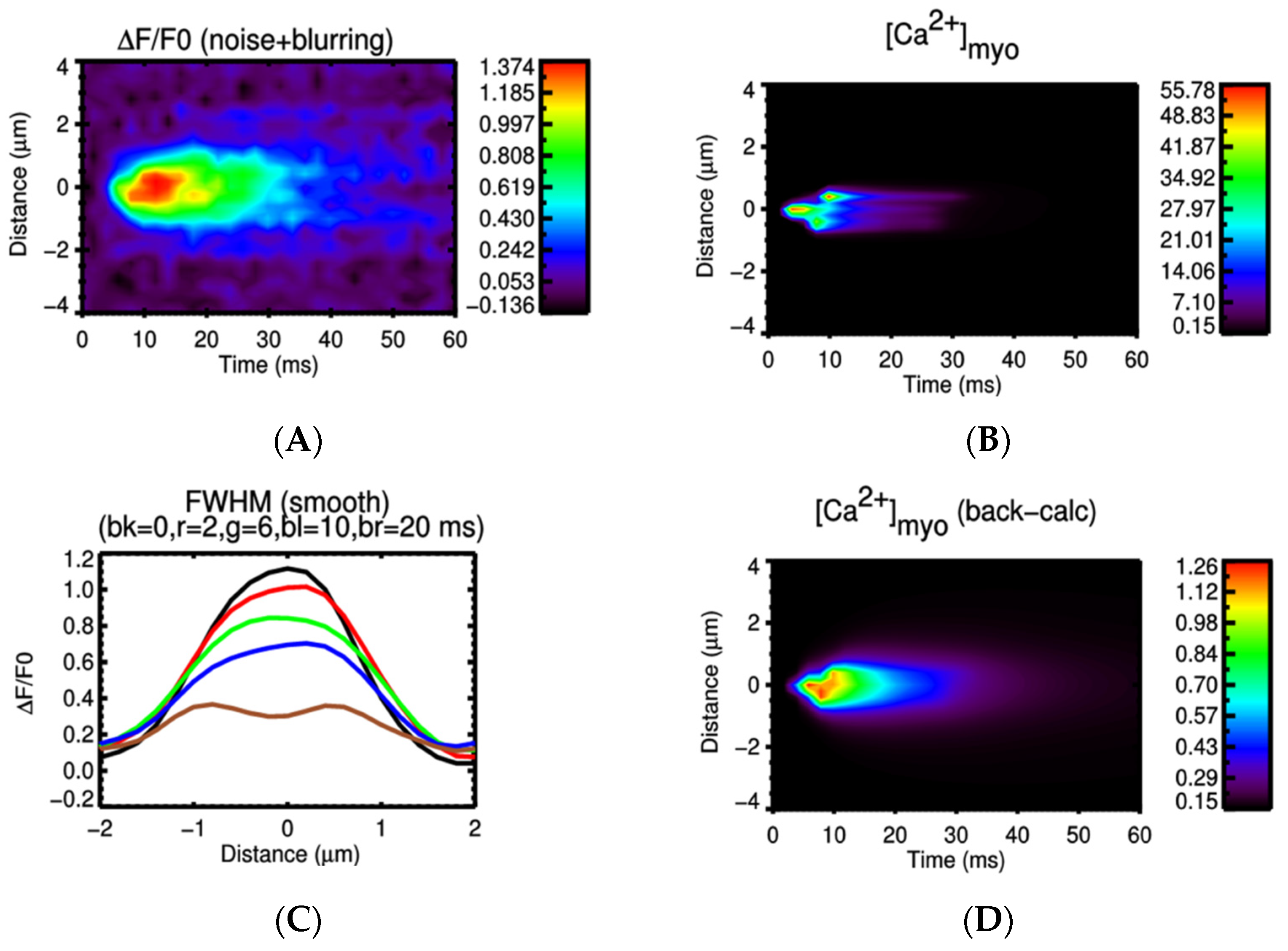
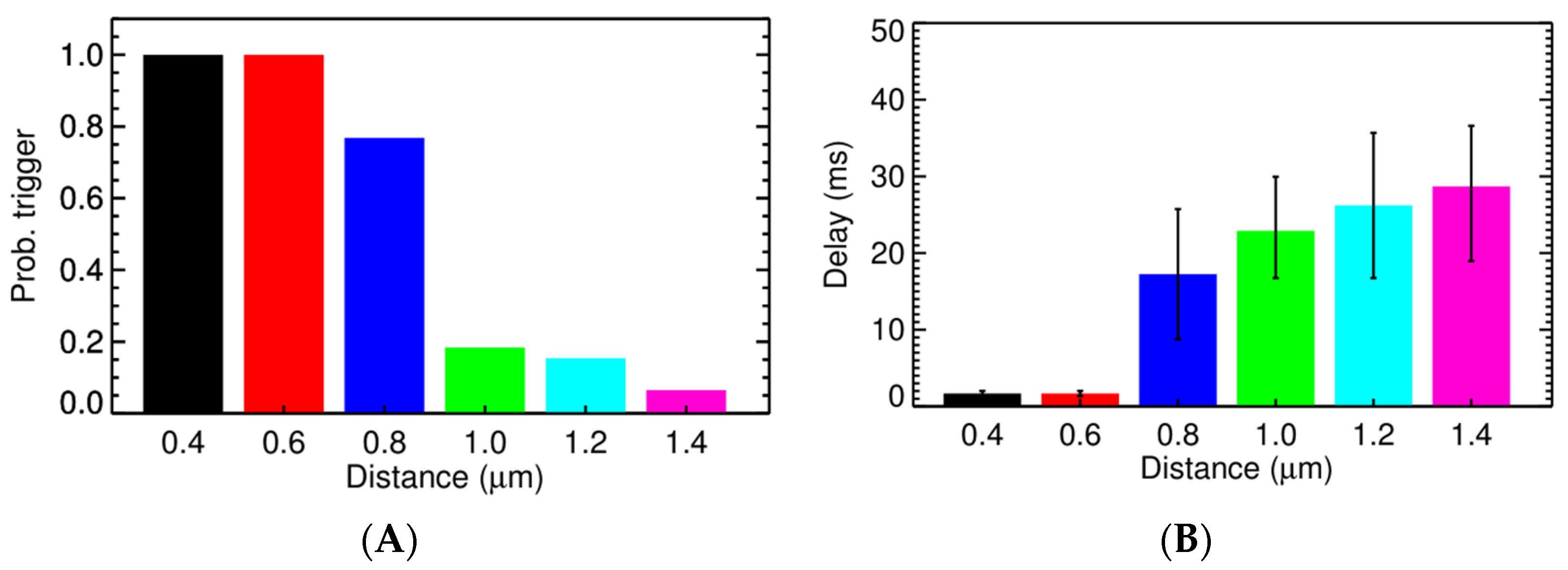
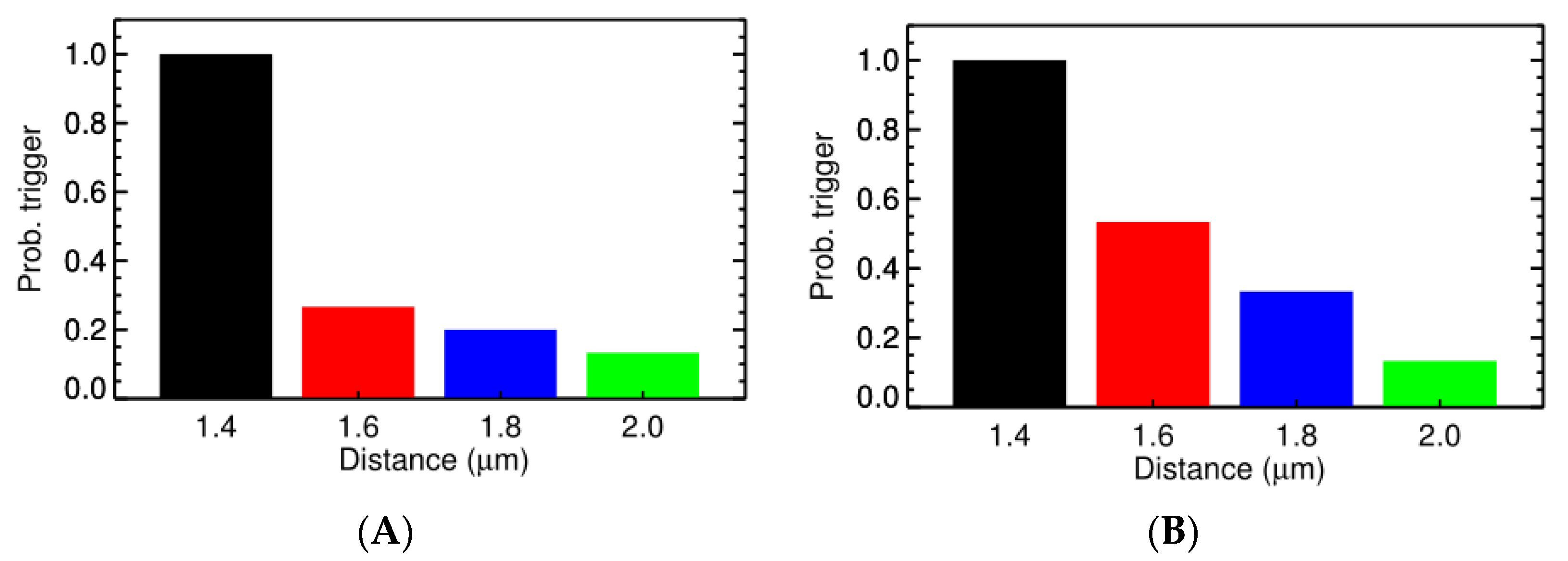
3.2. Ca2+ Spark Induced Ca2+ Sparks
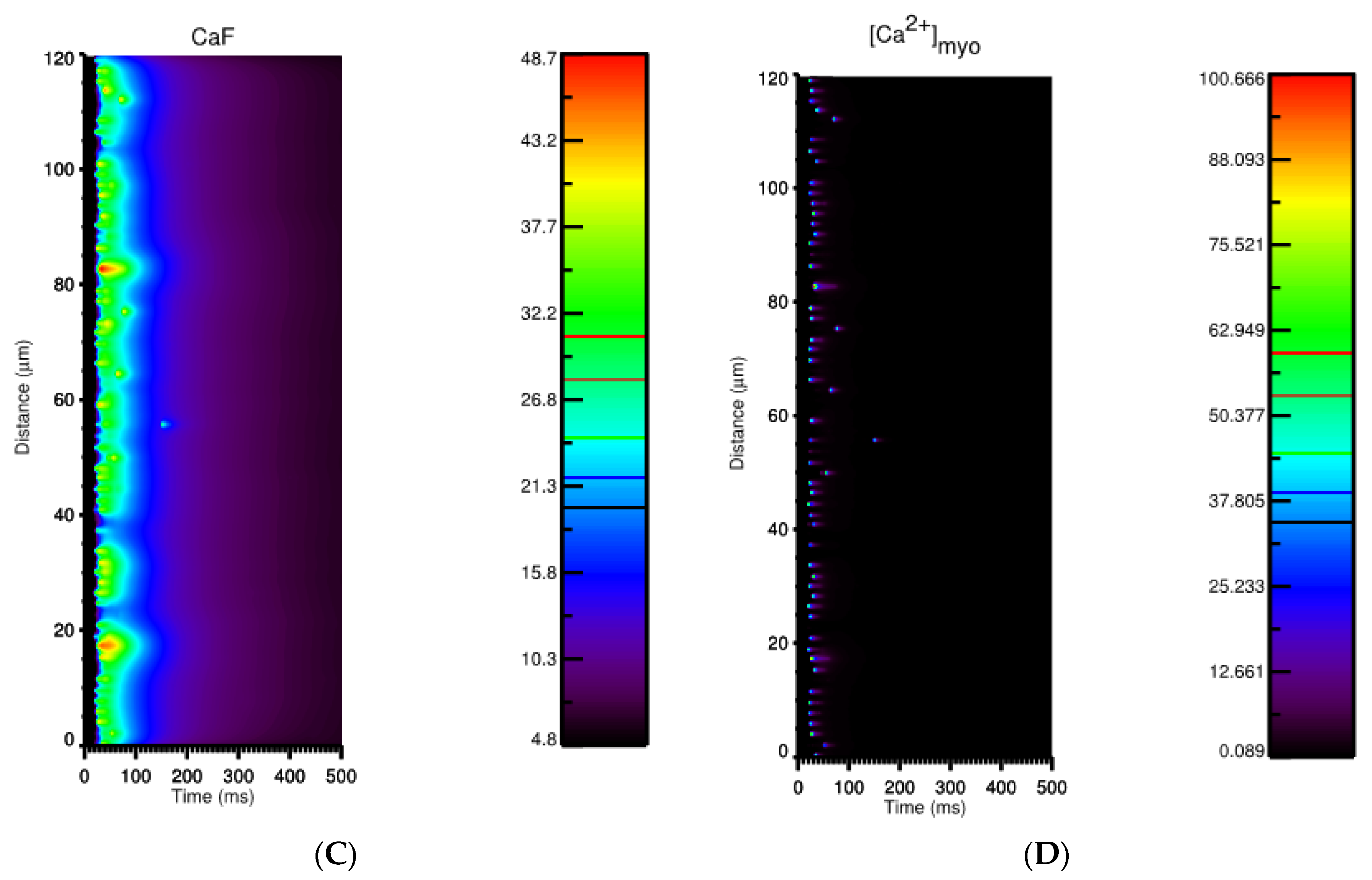
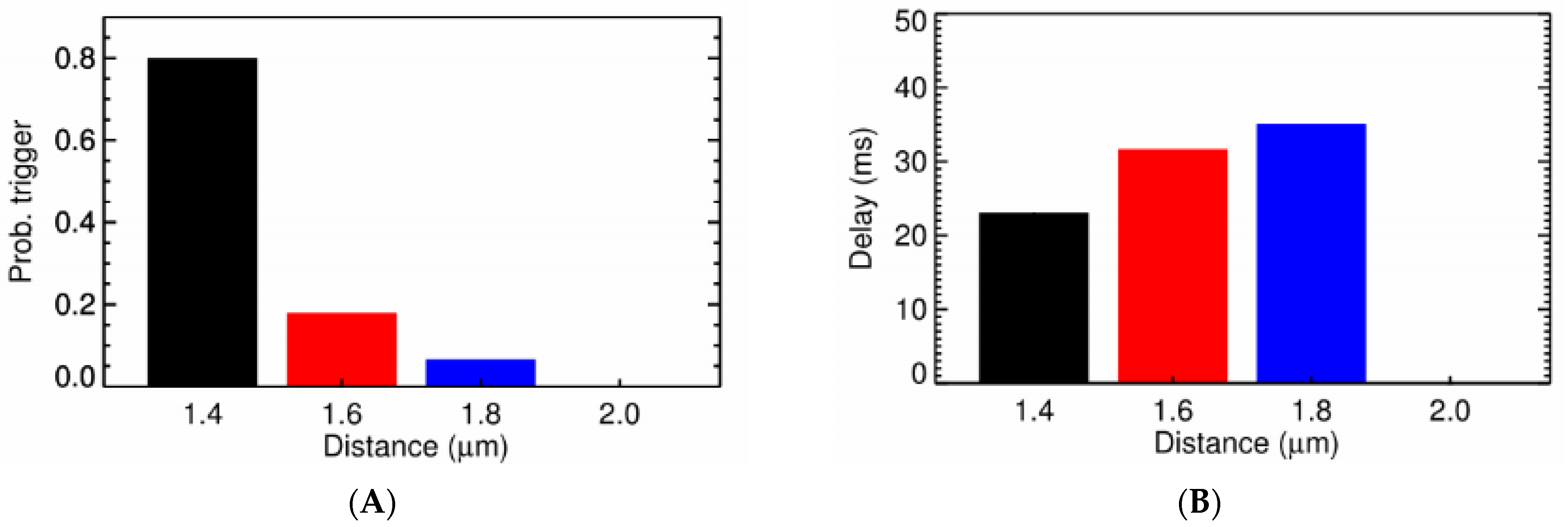
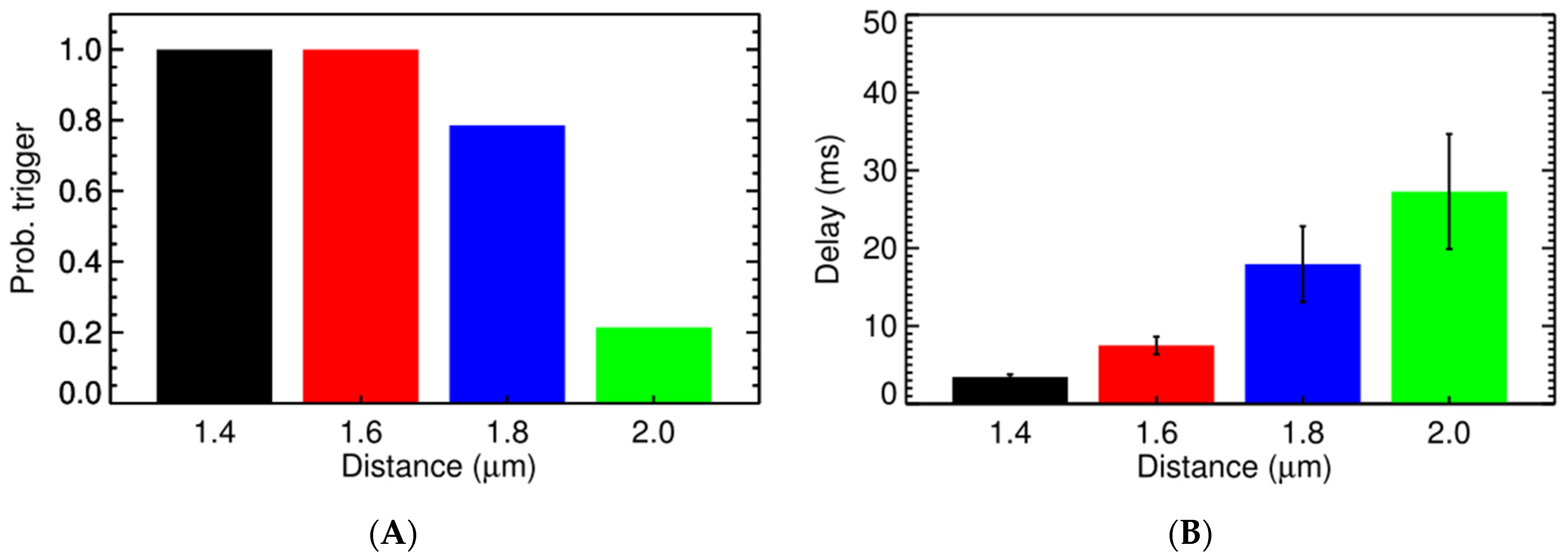
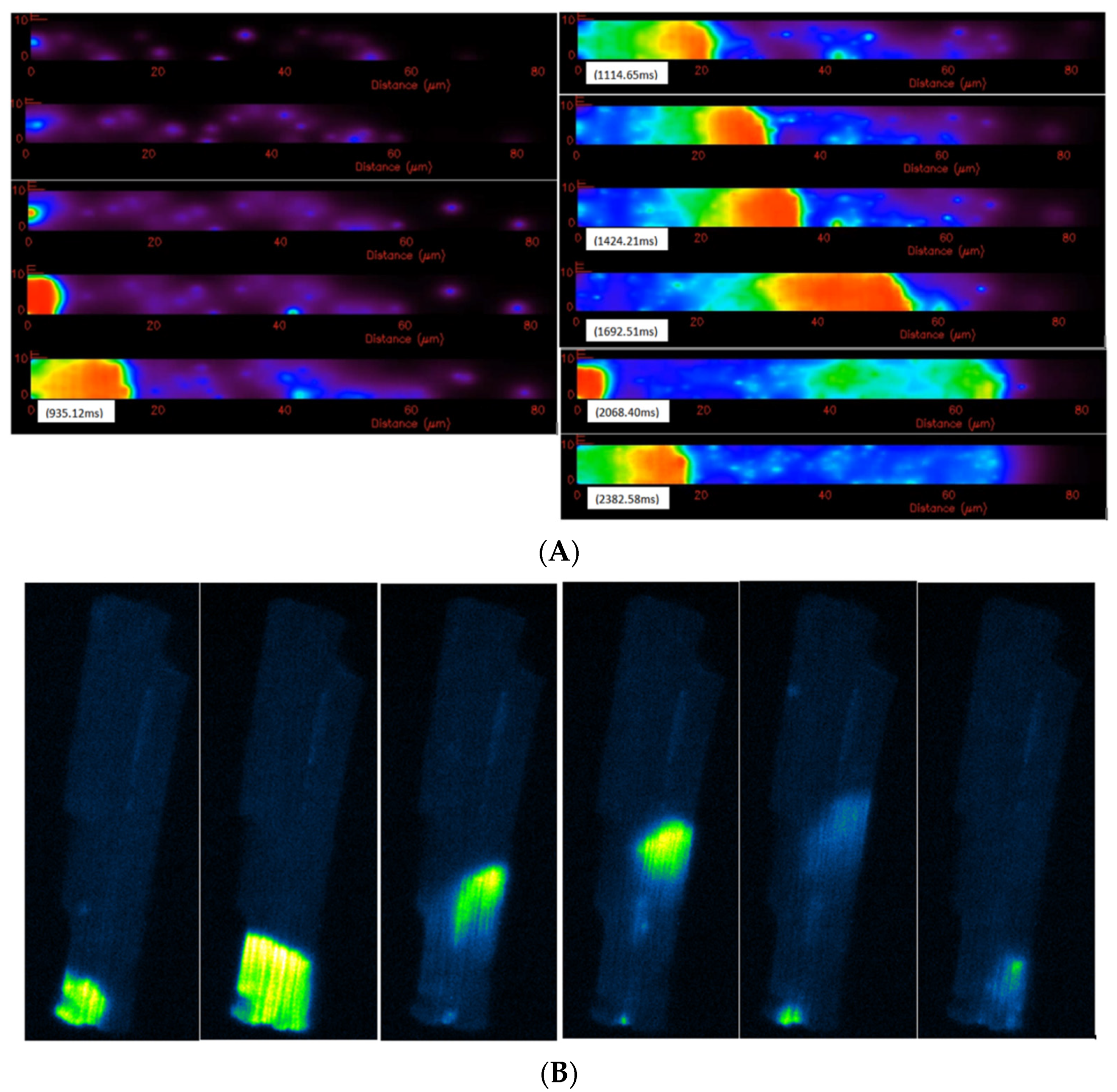
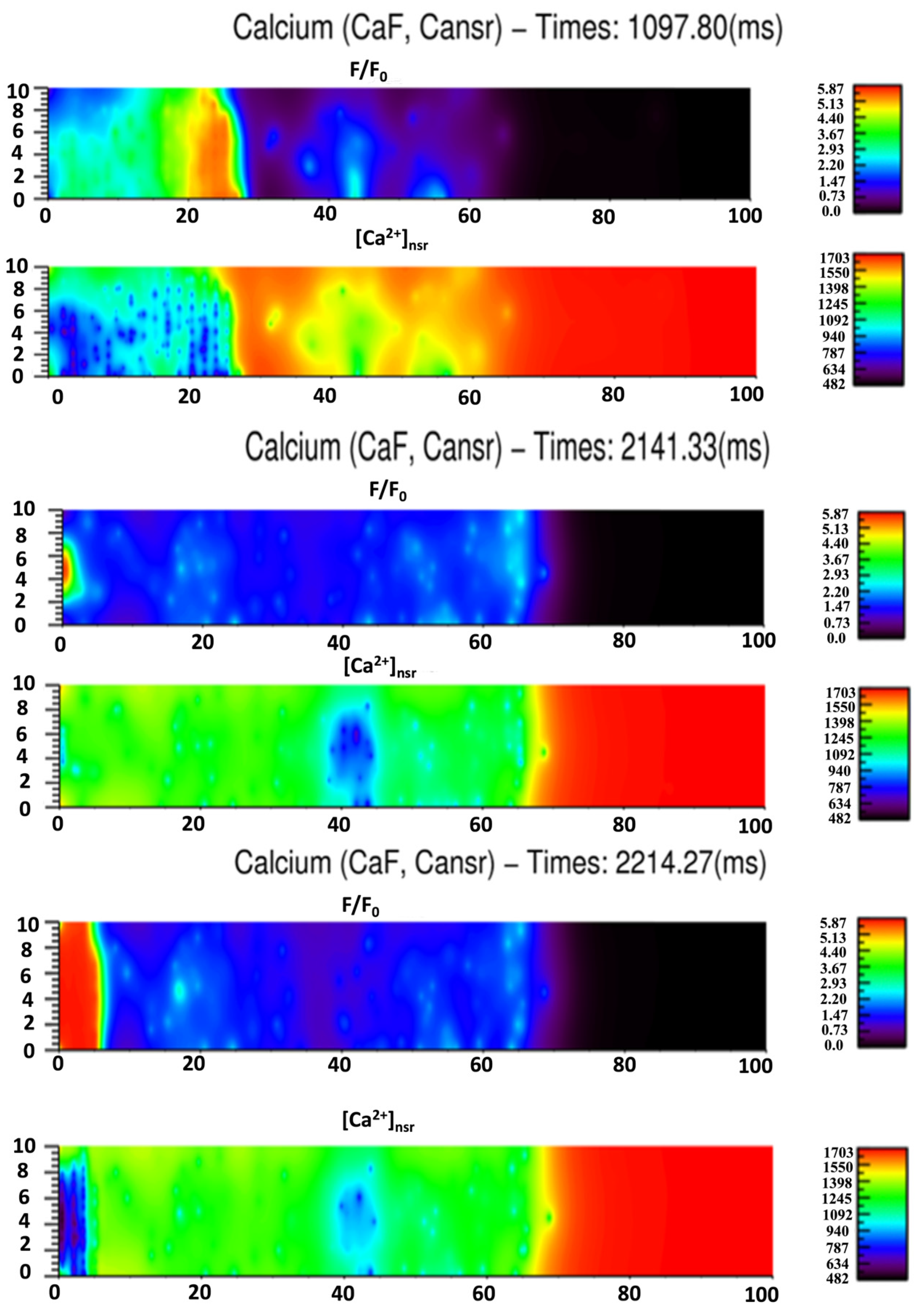
3.3. Ca2+ Spark Induced Ca2+ Waves
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Parameters

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Definition | Value |
|---|---|---|
| Vcell | Cell volume | 24.96 (pL) |
| Vmyo | Myoplasmic volume | 50% × Vcell |
| Vnsr | Network SR volume | 3.2% × Vcell |
| Vjsr | Junctional SR volume | 0.5% × Vcell |
| Dyad_height | The distance between jSR and T-tubule | 12 nm |
| Couplon_size | The size of the subspace | 300 nm |
| X_len | Cell dimension | 120 µm |
| Y_len | Cell dimension | 20.8 µm |
| Z_len | Cell dimension | 10.0 µm |
| Dmyo(x,y,z) | Free cytosolic calcium diffusion | 270 µm2/s |
| Dnsr(x,y,z) | Free SR calcium diffusion | 60 µm2/s |
| Ddye(x,y,z) | Fluorescence and calcium-bound fluorescence diffusion | 90 µm2/s |
| FT | Total fluorescence (Fluo-3) | 50 µM |
| kon | Binding constant for fluorescence | 80 µM−1.s−1 |
| koff | Unbinding constant for fluorescence | 72 s−1 |
| Ecc | Change in free energy between closed RyR2 pairs | −0.872 kBT |
| Eoo | Change in free energy between open RyR2 pairs | −1.15 kBT |
| kB | Boltzmann constant | 1.381 × 10−23 J/K |
| T | Temperature | 295.15 K |
| [K+]o | Extracellular potassium concentration | 5.4 mM |
| [Na+]o | Extracellular sodium concentration | 140 mM |
| [Ca2+]o | Extracellular calcium concentration | 1.8 mM |
| [K+]i | Cytosolic potassium concentration | 143.72 mM |
| [Na+]i | Cytosolic sodium concentration | 10.2 mM |
| [Ca2+]i | Cytosolic calcium concentration | 0.096 µM |
| [Ca2+]sr | SR calcium concentration | 1.02 mM |
| BmyoT | Total myoplasmic buffer concentration | 3.703026 × 10−2 µM |
| Km,myo | Disassociation constant of myoplasm buffer | 1.1900 µM |
| [Trpn] | Troponin Ca2+ binding sites concentration (k+ = 2.37 µM−1.s−1, k− = 0.032 s−1) | 140 µM |
| [Calm] | The dynamics buffers with Kd = 2.38 µM (k+ = 30 µM−1.s−1, k− = 71.4 s−1) | 24 µM |
| [B]SL | The SL buffer (k+ = 115 µM−1.s−1, k− = 1000 s−1) | 750 µM |
| [B]SR | The SR buffer with Kd=0.86µM (k+ = 115 µM−1.s−1, k− = 100 s−1) | 47 µM |
| Apump | Concentration of SERCA pump | 300 µM |
| Kp,myo | The binding affinity of cytosolic calcium to SERCA | 900 µM |
| Kp,nsr | The binding affinity of SR calcium to SERCA | 2150 µM |
| vTrefill | Total refill rate for 20,000 CRUs | 3 (1/s/(L-cyt)) |
| vTefflux | Total efflux rate for 20,000 CRUs | 120 (1/s/(L-cyt)) |
| iryr | Single channel RyR2 current | 0.2 (pA) |
| gbCa | Background Ca2+ conductance | 2.9 × 10−4 (mS/µF) |
| gbNa | Background Na+ conductance | 1.066 × 10−4 (mS/µF) |
| gbK | Background K+ conductance | 0 (mS/µF) |
| gNa | Na+ conductance | 13 (mS/µF) |
| gK1 | K1 conductance | 0.2 (mS/µF) |
| gKss | Kss conductance | 4.21 × 10−2 (mS/µF) |
| gKtof | Ktof conductance | 0.0798 (mS/µF) |
| gKtos | Ktos conductance | 6.29 × 10−2 (mS/µF) |
| Am | Cell surface area | |
| Ipmca | Maximum PMCA current density | 0.10 (µA/µF) |
| Incx | Maximum NCX current density | 750 (µA/µF) |
| INaK | Maximum NaK current density | 0.88 (µA/µF) |
| eta_RyR | Hill coefficient of Ca2+-dependent in RyR | 2.2 |
| eta_LCC | Hill coefficient of Ca2+-dependent in LCC | 2.0 |
| Ej | Coupling strength energy (for 49 RyRs) | 0.0714 |
| Ecc | Coupling energy between 2 closed channels | −0.78 (kBT) |
| Eoo | Coupling energy between 2 open channels | −1.26 (kBT) |
References
- Cheng, H.; Lederer, W.J.; Cannell, M.B. Calcium Sparks: Elementary Events Underlying Excitation-Contraction Coupling in Heart Muscle. Science 1993, 262, 740–744. [Google Scholar] [CrossRef]
- Cheng, H.; Lederer, M.R.; Lederer, W.J.; Cannell, M.B. Calcium sparks and [Ca2+]i waves in cardiac myocytes. Am. J. Physiol. 1996, 270, C148–C159. [Google Scholar] [CrossRef]
- Wier, W.G.; ter Keurs, H.E.; Marban, E.; Gao, W.D.; Balke, C.W. Ca2+ ‘sparks’ and waves in intact ventricular muscle resolved by confocal imaging. Circ. Res. 1997, 81, 462–469. [Google Scholar] [CrossRef]
- Lukyanenko, V.; Gyorke, S. Ca2+ sparks and Ca2+ waves in saponin-permeabilized rat ventricular myocytes. J. Physiol. 1999, 521, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.E.; Eisner, D.A.; O’Neill, S.C. Depressed ryanodine receptor activity increases variability and duration of the systolic Ca2+ transient in rat ventricular myocytes. Circ. Res. 2002, 91, 585–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, M.E.; O’Neill, S.C.; Eisner, D.A. Sarcoplasmic reticulum calcium content fluctuation is the key to cardiac alternans. Circ. Res. 2004, 94, 650–656. [Google Scholar] [CrossRef] [Green Version]
- Petrovič, P.; Valent, I.; Cocherová, E.; Pavelková, J.; Zahradníková, A. Ryanodine receptor gating controls generation of diastolic calcium waves in cardiac myocytes. J. Gen. Physiol. 2015, 145, 489–511. [Google Scholar] [CrossRef] [Green Version]
- Orchard, C.H.; Eisner, D.A.; Allen, D.G. Oscillations of intracellular Ca2+ in mammalian cardiac muscle. Nature 1983, 304, 735–738. [Google Scholar] [CrossRef] [PubMed]
- Wier, W.G.; Blatter, L.A. Ca2+-oscillations and Ca2+-waves in mammalian cardiac and vascular smooth muscle cells. Cell Calcium 1991, 12, 241–254. [Google Scholar] [CrossRef]
- Wier, W.G.; Cannell, M.B.; Berlin, J.R.; Marban, E.; Lederer, W.J. Cellular and subcellular heterogeneity of [Ca2+]i in single heart cells revealed by fura-2. Science 1987, 235, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Q.; Stern, M.D.; Ríos, E.; Cheng, H. The quantal nature of Ca2+ sparks and in situ operation of the ryanodine receptor array in cardiac cells. Proc. Natl. Acad. Sci. USA 2004, 101, 3979–3984. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.X.; Wang, S.; Song, L.S.; Han, T.; Cheng, H. Polymorphism of Ca2+ sparks evoked from in-focus Ca2+ release units in cardiac myocytes. Biophys. J. 2004, 86, 182–190. [Google Scholar] [CrossRef] [Green Version]
- Niggli, E.; Shirokova, N. A guide to sparkology: The taxonomy of elementary cellular Ca2+ signaling events. Cell Calcium 2007, 42, 379–387. [Google Scholar] [CrossRef]
- Cheng, H.; Lederer, W.J. Calcium sparks. Physiol. Rev. 2008, 88, 1491–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pratusevich, V.R.; Balke, C.W. Factors shaping the confocal image of the calcium spark in cardiac muscle cells. Biophys. J. 1996, 71, 2942–2957. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.D.; Keizer, J.E.; Stern, M.D.; Lederer, W.J.; Cheng, H. A simple numerical model of calcium spark formation and detection in cardiac myocytes. Biophys. J. 1998, 75, 15–32. [Google Scholar] [CrossRef] [Green Version]
- Izu, L.T.; Mauban, J.R.H.; Balke, C.W.; Wier, W.G. Large currents generate cardiac Ca2+ sparks. Biophys. J. 2001, 80, 88–102. [Google Scholar] [CrossRef] [Green Version]
- Koh, X.; Srinivasan, B.; Ching, H.S.; Levchenko, A. A 3D Monte Carlo analysis of the role of dyadic space geometry in spark generation. Biophys. J. 2006, 90, 1999–2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hake, J.; Edwards, A.G.; Yu, Z.; Kekenes-Huskey, P.M.; Michailova, A.P.; McCammon, J.A.; Holst, M.J.; Hoshijima, M.; McCulloch, A.D. Modelling cardiac calcium sparks in a three-dimensional reconstruction of a calcium release unit. J. Physiol. 2012, 590, 4403–4422. [Google Scholar] [CrossRef] [Green Version]
- Sobie, E.A.; Dilly, K.W.; dos Santos, C.J.; Lederer, W.J.; Jafri, M.S. Termination of Cardiac Ca2+ Sparks: An Investigative Mathematical Model of Calcium-Induced Calcium Release. Biophys. J. 2002, 83, 59–78. [Google Scholar] [CrossRef] [Green Version]
- Laver, D.R.; Kong, C.H.T.; Imtiaz, M.S.; Cannell, M.B. Termination of calcium-induced calcium release by induction decay: An emergent property of stochastic channel gating and molecular scale architecture. J. Mol. Cell. Cardiol. 2013, 54, 98–100. [Google Scholar] [CrossRef]
- Kong, C.H.T.; Laver, D.R.; Cannell, M.B. Extraction of Sub-microscopic Ca Fluxes from Blurred and Noisy Fluorescent Indicator Images with a Detailed Model Fitting Approach. PLoS Comput. Biol. 2013, 9, e1002931. [Google Scholar] [CrossRef] [Green Version]
- Backx, P.H.; de Tombe, P.P.; Van Deen, J.H.; Mulder, B.J.; ter Keurs, H.E. A model of propagating calcium-induced calcium release mediated by calcium diffusion. J. Gen. Physiol. 1989, 93, 963–977. [Google Scholar] [CrossRef] [Green Version]
- Keizer, J.; Smith, G.D. Spark-to-wave transition: Saltatory transmission of calcium waves in cardiac myocytes. Biophys. Chem. 1998, 72, 87–100. [Google Scholar] [CrossRef]
- Keizer, J.; Smith, G.D.; Ponce-Dawson, S.; Pearson, J.E. Saltatory propagation of Ca2+ waves by Ca2+ sparks. Biophys. J. 1998, 75, 595–600. [Google Scholar] [CrossRef] [Green Version]
- Izu, L.T.; Wier, W.G.; Balke, C.W. Evolution of Cardiac Calcium Waves from Stochastic Calcium Sparks. Biophys. J. 2001, 80, 103–120. [Google Scholar] [CrossRef] [Green Version]
- Ramay, H.R.; Jafri, M.S.; Lederer, W.J.; Sobie, E.A. Predicting local SR Ca2+ dynamics during Ca2+ wave propagation in ventricular myocytes. Biophys. J. 2010, 98, 2515–2523. [Google Scholar] [CrossRef] [Green Version]
- Rovetti, R.; Cui, X.; Garfinkel, A.; Weiss, J.N.; Qu, Z. Spark-induced sparks as a mechanism of intracellular calcium alternans in cardiac myocytes. Circ. Res. 2010, 106, 1582–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nivala, M.; de Lange, E.; Rovetti, R.; Qu, Z. Computational modeling and numerical methods for spatiotemporal calcium cycling in ventricular myocytes. Front. Physiol. 2012, 3, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berlin, J.R. Spatiotemporal changes of Ca2+ during electrically evoked contractions in atrial and ventricular cells. Am. J. Physiol. 1995, 269, H1165–H1170. [Google Scholar] [CrossRef] [PubMed]
- Kockskamper, J.; Sheehan, K.A.; Bare, D.J.; Lipsius, S.L.; Mignery, G.A.; Blatter, L.A. Activation and propagation of Ca2+ release during excitation-contraction coupling in atrial myocytes. Biophys. J. 2001, 81, 2590–2605. [Google Scholar] [CrossRef] [Green Version]
- Thul, R.; Coombes, S.; Roderick, H.L.; Bootman, M.D. Subcellular calcium dynamics in a whole-cell model of an atrial myocyte. Proc. Natl. Acad. Sci. USA 2012, 109, 2150–2155. [Google Scholar] [CrossRef] [Green Version]
- Shiferaw, Y.; Karma, A. Turing instability mediated by voltage and calcium diffusion in paced cardiac cells. Proc. Natl. Acad. Sci. USA 2006, 103, 5670–5675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Lancaster, M.; Holden, A.V. A Three Dimensional Ventricular E-Cell (3Dv E-Cell) with Stochastic Intracellular Ca2+ Handling. In International Conference on Functional Imaging and Modeling of the Heart; Springer: Berlin/Heidelberg, Germany, 2007; pp. 180–189. [Google Scholar]
- Soeller, C.; Jayasinghe, I.D.; Li, P.; Holden, A.V.; Cannell, M.B. Three-dimensional high-resolution imaging of cardiac proteins to construct models of intracellular Ca2+ signalling in rat ventricular myocytes. Exp. Physiol. 2009, 94, 496–508. [Google Scholar] [CrossRef]
- Hoang-Trong, M.T.; Ullah, A.; Lederer, W.J.; Jafri, M.S. Cardiac Alternans Occurs through the Synergy of Voltage- and Calcium-Dependent Mechanisms. Membranes 2021, 11, 794. [Google Scholar] [CrossRef] [PubMed]
- Satoh, H.; Delbridge, L.M.; Blatter, L.A.; Bers, D.M. Surface:volume relationship in cardiac myocytes studied with confocal microscopy and membrane capacitance measurements: Species-dependence and developmental effects. Biophys. J. 1996, 70, 1494–1504. [Google Scholar] [CrossRef] [Green Version]
- Delbridge, L.M.; Satoh, H.; Yuan, W.; Bassani, J.W.; Qi, M.; Ginsburg, K.S.; Samarel, A.M.; Bers, D.M. Cardiac myocyte volume, Ca2+ fluxes, and sarcoplasmic reticulum loading in pressure-overload hypertrophy. Am. J. Physiol. 1997, 272, H2425–H2435. [Google Scholar] [CrossRef] [PubMed]
- Swift, F.; Stromme, T.A.; Amundsen, B.; Sejersted, O.M.; Sjaastad, I. Slow diffusion of K+ in the T tubules of rat cardiomyocytes. J. Appl. Physiol. 2006, 101, 1170–1176. [Google Scholar] [CrossRef] [Green Version]
- Padmala, S.; Demir, S.S. Computational model of the ventricular action potential in adult spontaneously hypertensive rats. J. Cardiovasc. Electrophysiol. 2003, 14, 990–995. [Google Scholar] [CrossRef] [PubMed]
- Pandit, S.V.; Clark, R.B.; Giles, W.R.; Demir, S.S. A mathematical model of action potential heterogeneity in adult rat left ventricular myocytes. Biophys. J. 2001, 81, 3029–3051. [Google Scholar] [CrossRef] [Green Version]
- Andersen, O.S.; Koeppe, R.E., II. Bilayer thickness and membrane protein function: An energetic perspective. Annu. Rev. Biophys. Biomol. Struct. 2007, 36, 107–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, E. Quantitative ultrastructural analysis in cardiac membrane physiology. Am. J. Physiol. 1978, 235, C147–C158. [Google Scholar] [CrossRef] [Green Version]
- Stewart, J.M.; Page, E. Improved stereological techniques for studying myocardial cell growth: Application to external sarcolemma, T system, and intercalated disks of rabbit and rat hearts. J. Ultrastruct. Res. 1978, 65, 119–134. [Google Scholar] [CrossRef]
- Page, E.; Surdyk-Droske, M. Distribution, surface density, and membrane area of diadic junctional contacts between plasma membrane and terminal cisterns in mammalian ventricle. Circ. Res. 1979, 45, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Kawai, M.; Hussain, M.; Orchard, C.H. Excitation-contraction coupling in rat ventricular myocytes after formamide-induced detubulation. Am. J. Physiol. 1999, 277, H603–H609. [Google Scholar] [CrossRef] [PubMed]
- Soeller, C.; Cannell, M.B. Examination of the Transverse Tubular System in Living Cardiac Rat Myocytes by 2-Photon Microscopy and Digital Image Processing Techniques. Circ. Res. 1999, 84, 266–275. [Google Scholar] [CrossRef]
- Brette, F.; Orchard, C.H. No apparent requirement for neuronal sodium channels in excitation-contraction coupling in rat ventricular myocytes. Circ. Res. 2006, 98, 667–674. [Google Scholar] [CrossRef] [Green Version]
- Pasek, M.; Brette, F.; Nelson, A.; Pearce, C.; Qaiser, A.; Christe, G.; Orchard, C.H. Quantification of t-tubule area and protein distribution in rat cardiac ventricular myocytes. Prog. Biophys. Mol. Biol. 2008, 96, 244–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brette, F.; Orchard, C. T-tubule function in mammalian cardiac myocytes. Circ. Res. 2003, 92, 1182–1192. [Google Scholar] [CrossRef]
- Forbes, M.S.; Hawkey, L.A.; Sperelakis, N. The transverse-axial tubular system (TATS) of mouse myocardium: Its morphology in the developing and adult animal. Am. J. Anat. 1984, 170, 143–162. [Google Scholar] [CrossRef]
- Bers, D.M. Excitation-Contraction Coupling and Cardiac Contractile Force, 2nd ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001. [Google Scholar]
- Wei, S.; Guo, A.; Chen, B.; Kutschke, W.; Xie, Y.P.; Zimmerman, K.; Weiss, R.M.; Anderson, M.E.; Cheng, H.; Song, L.S. T-tubule remodeling during transition from hypertrophy to heart failure. Circ. Res. 2010, 107, 520–531. [Google Scholar] [CrossRef] [Green Version]
- Polakova, E.; Sobie, E.A. Alterations in T-tubule and dyad structure in heart disease: Challenges and opportunities for computational analyses. Cardiovasc. Res. 2013, 98, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Heinzel, F.R.; Bito, V.; Biesmans, L.; Wu, M.; Detre, E.; von Wegner, F.; Claus, P.; Dymarkowski, S.; Maes, F.; Bogaert, J.; et al. Remodeling of T-tubules and reduced synchrony of Ca2+ release in myocytes from chronically ischemic myocardium. Circ. Res. 2008, 102, 338–346. [Google Scholar] [CrossRef] [Green Version]
- Wagner, E.; Lauterbach, M.A.; Kohl, T.; Westphal, V.; Williams, G.S.B.; Steinbrecher, J.H.; Streich, J.-H.; Korff, B.; Tuan, H.-T.M.; Hagen, B.; et al. Stimulated emission depletion live-cell super-resolution imaging shows proliferative remodeling of T-tubule membrane structures after myocardial infarction. Circ. Res. 2012, 111, 402–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyon, A.R.; MacLeod, K.T.; Zhang, Y.; Garcia, E.; Kanda, G.K.; Lab, M.J.; Korchev, Y.E.; Harding, S.E.; Gorelik, J. Loss of T-tubules and other changes to surface topography in ventricular myocytes from failing. Proc. Natl. Acad. Sci. USA 2009, 106, 6854–6859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, A.; Zhang, C.; Wei, S.; Chen, B.; Song, L.S. Emerging mechanisms of T-tubule remodelling in heart failure. Cardiovasc. Res. 2013, 98, 204–215. [Google Scholar] [CrossRef] [Green Version]
- Chen-Izu, Y.; Mcculle, S.L.; Ward, C.W.; Soeller, C.; Allen, B.M.; Rabang, C.; Cannell, M.B.; Balke, C.W.; Izu, L.T. Three-dimensional distribution of ryanodine receptor clusters in cardiac myocytes. Biophys. J. 2006, 91, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kieval, R.S.; Bloch, R.J.; Lindenmayer, G.E.; Ambesi, A.; Lederer, W.J. Immunofluorescence localization of the Na-Ca exchanger in heart cells. Am. J. Physiol. 1992, 263, C545–C550. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.S.; Mottino, G.; Reid, D.; Molday, R.S.; Philipson, K.D. Distribution of the Na+-Ca2+ exchange protein in mammalian cardiac myocytes: An immunofluorescence and immunocolloidal gold-labeling study. J. Cell Biol. 1992, 117, 337–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scriven, D.R.; Dan, P.; Moore, E.D. Distribution of proteins implicated in excitation-contraction coupling in rat ventricular myocytes. Biophys. J. 2000, 79, 2682–2691. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Pascarel, C.; Steele, D.S.; Komukai, K.; Brette, F.; Orchard, C.H. Na+-Ca2+ exchange activity is localized in the T-tubules of rat ventricular myocytes. Circ. Res. 2002, 91, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Despa, S.; Brette, F.; Orchard, C.H.; Bers, D.M. Na/Ca exchange and Na/K-ATPase function are equally concentrated in transverse tubules of rat ventricular myocytes. Biophys. J. 2003, 85, 3388–3396. [Google Scholar] [CrossRef] [Green Version]
- Periasamy, M.; Huke, S. SERCA pump level is a critical determinant of Ca2+ homeostasis and cardiac contractility. J. Mol. Cell. Cardiol. 2001, 33, 1053–1063. [Google Scholar] [CrossRef]
- Greene, A.L.; Lalli, M.J.; Ji, Y.; Babu, G.J.; Grupp, I.; Sussman, M.; Periasamy, M. Overexpression of SERCA2b in the heart leads to an increase in sarcoplasmic reticulum calcium transport function and increased cardiac contractility. J. Biol. Chem. 2000, 275, 24722–24727. [Google Scholar] [CrossRef] [Green Version]
- Drago, G.A.; Colyer, J.; Lederer, W.J. Immunofluorescence Localization of SERCA2a and the Phosphorylated Forms of Phospholamban in Intact Rat Cardiac Ventricular Myocytes. Ann. N. Y. Acad. Sci. 1998, 853, 273–279. [Google Scholar] [CrossRef]
- Wang, J.H. Tracer-diffusion in Liquids. IV. Self-diffusion of Calcium Ion and Chloride Ion in Aqueous Calcium Chloride Solutions. J. Am. Chem. Soc. 1953, 75, 1769–1770. [Google Scholar] [CrossRef]
- Kushmerick, M.J.; Podolsky, R.J. Ionic mobility in muscle cells. Science 1969, 166, 1297–1298. [Google Scholar] [CrossRef] [PubMed]
- Kargacin, G.; Fay, F.S. Ca2+ movement in smooth muscle cells studied with one- and two-dimensional diffusion models. Biophys. J. 1991, 60, 1088–1100. [Google Scholar] [CrossRef] [Green Version]
- Allbritton, N.L.; Meyer, T.; Stryer, L. Range of messenger action of calcium ion and inositol 1,4,5-trisphosphate. Science 1992, 258, 1812–1815. [Google Scholar] [CrossRef]
- Wu, X.; Bers, D.M. Sarcoplasmic reticulum and nuclear envelope are one highly interconnected Ca2+ store throughout cardiac myocyte. Circ. Res. 2006, 99, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Swietach, P.; Spitzer, K.W.; Vaughan-Jones, R.D. Ca2+-mobility in the sarcoplasmic reticulum of ventricular myocytes is low. Biophys. J. 2008, 95, 1412–1427. [Google Scholar] [CrossRef] [Green Version]
- Picht, E.; Zima, A.V.; Shannon, T.R.; Duncan, A.M.; Blatter, L.A.; Bers, D.M. Dynamic calcium movement inside cardiac sarcoplasmic reticulum during release. Circ. Res. 2011, 108, 847–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobie, E.A.; Lederer, W.J. Dynamic local changes in sarcoplasmic reticulum calcium: Physiological and pathophysiological roles. J. Mol. Cell. Cardiol. 2012, 52, 304–311. [Google Scholar] [CrossRef] [Green Version]
- Zima, A.V.; Picht, E.; Bers, D.M.; Blatter, L.A. Termination of cardiac Ca2+ sparks: Role of intra-SR [Ca2+], release flux, and intra-SR Ca2+ diffusion. Circ. Res. 2008, 103, e105–e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brochet, D.X.P.; Yang, D.; Di Maio, A.; Lederer, W.J.; Franzini-Armstrong, C.; Cheng, H. Ca2+ blinks: Rapid nanoscopic store calcium signaling. Proc. Natl. Acad. Sci. USA 2005, 102, 3099–3104. [Google Scholar] [CrossRef] [Green Version]
- Terentyev, D.; Viatchenko-Karpinski, S.; Gyorke, I.; Volpe, P.; Williams, S.C.; Gyorke, S. Calsequestrin determines the functional size and stability of cardiac intracellular calcium stores: Mechanism for hereditary arrhythmia. Proc. Natl. Acad. Sci. USA 2003, 100, 11759–11764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubalova, Z.; Gyorke, I.; Terentyeva, R.; Viatchenko-Karpinski, S.; Terentyev, D.; Williams, S.C.; Gyorke, S. Modulation of cytosolic and intra-sarcoplasmic reticulum calcium waves by calsequestrin in rat cardiac myocytes. J. Physiol. 2004, 561, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Michailova, A.; Del Principe, F.; Egger, M.; Niggli, E. Spatiotemporal Features of Ca2+ Buffering and Diffusion in Atrial Cardiac Myocytes with Inhibited Sarcoplasmic Reticulum. Biophys. J. 2002, 83, 3134–3151. [Google Scholar] [CrossRef] [Green Version]
- Harkins, A.B.; Kurebayashi, N.; Baylor, S.M. Resting myoplasmic free calcium in frog skeletal muscle fibers estimated with fluo-3. Biophys. J. 1993, 65, 865–881. [Google Scholar] [CrossRef] [Green Version]
- Baylor, S.M.; Hollingworth, S. Model of sarcomeric Ca2+ movements, including ATP Ca2+ binding and diffusion, during activation of frog skeletal muscle. J. Gen. Physiol. 1998, 112, 297–316. [Google Scholar] [CrossRef] [Green Version]
- Loughrey, C.M.; MacEachern, K.E.; Cooper, J.; Smith, G.L. Measurement of the dissociation constant of Fluo-3 for Ca2+ in isolated rabbit cardiomyocytes using Ca2+ wave characteristics. Cell Calcium 2003, 34, 1–9. [Google Scholar] [CrossRef]
- Williams, G.S.B.; Chikando, A.C.; Tuan, H.-T.M.; Sobie, E.A.; Lederer, W.J.; Jafri, M.S. Dynamics of Calcium Sparks and Calcium Leak in the Heart. Biophys. J. 2011, 101, 1287–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- HDF-Group. Available online: http://www.hdfgroup.org/solutions/HDF5/ (accessed on 14 December 2021).
- Lukyanenko, V.; Viatchenko-Karpinski, S.; Smirnov, A.; Wiesner, T.F.; Györke, S. Dynamic regulation of sarcoplasmic reticulum Ca2+ content and release by luminal Ca2+-sensitive leak in rat ventricular myocytes. Biophys. J. 2001, 81, 785–798. [Google Scholar] [CrossRef] [Green Version]
- Qin, J.; Valle, G.; Nani, A.; Nori, A.; Rizzi, N.; Priori, S.G.; Volpe, P.; Fill, M. Luminal Ca2+ regulation of single cardiac ryanodine receptors: Insights provided by calsequestrin and its mutants. J. Gen. Physiol. 2008, 131, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Gyorke, S.; Stevens, S.C.; Terentyev, D. Cardiac calsequestrin: Quest inside the SR. J. Physiol. 2009, 587, 3091–3094. [Google Scholar] [CrossRef]
- Brochet, D.X.P.; Lederer, W.J. Decomposition of a Calcium Spark in Cardiac Myocytes. Biophys. J. 2014, 104, 483a. [Google Scholar] [CrossRef] [Green Version]
- Macquaide, N.; Tuan, H.-T.M.; Hotta, J.-I.; Sempels, W.; Lenaerts, I.; Holemans, P.; Hofkens, J.; Jafri, M.S.; Willems, R.; Sipido, K.R. Functional consequences of RyR cluster fragmentation and redistribution in persistent atrial fibrillation. Cardiovasc. Res. 2014, 108, 387–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baddeley, D.; Jayasinghe, I.D.; Lam, L.; Rossberger, S.; Cannell, M.B.; Soeller, C. Optical single-channel resolution imaging of the ryanodine receptor distribution in rat cardiac myocytes. Proc. Natl. Acad. Sci. USA 2009, 106, 22275–22280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacQuaide, N.; Ramay, H.R.; Sobie, E.A.; Smith, G.L. Differential sensitivity of Ca2+ wave and Ca2+ spark events to ruthenium red in isolated permeabilised rabbit cardiomyocytes. J. Physiol. 2010, 588, 4731–4742. [Google Scholar] [CrossRef]
- Parker, I.; Zang, W.-J.; Wier, W.G. Ca2+ sparks involving multiple Ca2+ release sites along Z-lines in rat heart cells. J. Physiol. 1996, 497, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Blatter, L.A.; Huser, J.; Rios, E. Sarcoplasmic reticulum Ca2+ release flux underlying Ca2+ sparks in cardiac muscle. Proc. Natl. Acad. Sci. USA 1997, 94, 4176–4181. [Google Scholar] [CrossRef] [Green Version]
- Trafford, A.W.; Lipp, P.; O’Neill, S.C.; Niggli, E.; Eisner, D.A. Propagating calcium waves initiated by local caffeine application in rat ventricular myocytes. J. Physiol. 1995, 489, 319–326. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.L.; O’Neill, S.C. A comparison of the effects of ATP and tetracaine on spontaneous Ca2+ release from rat permeabilised cardiac myocytes. J. Physiol. 2001, 534, 37–47. [Google Scholar] [CrossRef]
- Jaffe, L.F. The path of calcium in cytosolic calcium oscillations: A unifying hypothesis. Proc. Natl. Acad. Sci. USA 1991, 88, 9883–9887. [Google Scholar] [CrossRef] [Green Version]
- Swietach, P.; Spitzer, K.W.; Vaughan-Jones, R.D. Modeling calcium waves in cardiac myocytes: Importance of calcium diffusion. Front. Biosci. 2010, 15, 661–680. [Google Scholar] [CrossRef] [Green Version]
- Kort, A.A.; Capogrossi, M.C.; Lakatta, E.G. Frequency, amplitude, and propagation velocity of spontaneous Ca++-dependent contractile waves in intact adult rat cardiac muscle and isolated myocytes. Circ. Res. 1985, 57, 844–855. [Google Scholar] [CrossRef] [Green Version]
- Stuyvers, B.D.; Boyden, P.A.; Keurs, H.E.D.J.T. Calcium Waves. Physiological Relevance in Cardiac Function. Circ. Res. 2000, 86, 1016–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, T.; Tanaka, H.; Oyamada, M.; Kawata, S.; Takamatsu, T. Three distinct types of Ca2+ waves in Langendorff-perfused rat heart revealed by real-time confocal microscopy. Circ. Res. 2000, 86, 1093–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jafri, M.S. On the Roles of Ca2+ Diffusion, Ca2 + Buffers, and the Endoplasmic Reticulum in IP3-lnduced Ca2+ Waves. Biophys. J. 1995, 69, 2139–2153. [Google Scholar] [CrossRef] [Green Version]
- Rossi, F.M.; Kao, J.P. Nmoc-DBHQ, a new caged molecule for modulating sarcoplasmic/endoplasmic reticulum Ca2+ ATPase activity with light flashes. J. Biol. Chem. 1997, 272, 3266–3271. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.; Kao, J.P.; Egger, M.; Niggli, E. Calcium waves driven by "sensitization" wave-fronts. Cardiovasc. Res. 2007, 74, 39–45. [Google Scholar] [CrossRef]
- Sobie, E.A.; Guatimosim, S.; Gómez-Viquez, L.; Song, L.-S.; Hartmann, H.; Jafri, S.M.; Lederer, W.J. The Ca2+ leak paradox and rogue ryanodine receptors: SR Ca2+ efflux theory and practice. Prog. Biophys. Mol. Biol. 2006, 90, 172–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, M.; Sikkel, M.B.; Desplantez, T.; Collins, T.P.; O’Gara, P.; Harding, S.E.; Lyon, A.R.; Macleod, K.T. Flecainide reduces wave frequency and mean spark amplitude in isolated rat ventricular cardiomyocytes. Cardiovasc. Res. 2012, 93, S90. [Google Scholar]
- Dedkova, E.N.; Blatter, L.A. Mitochondrial Ca2+ and the heart. Cell Calcium 2008, 44, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Aon, M.A.; Cortassa, S.; Akar, F.G.; Brown, D.A.; Zhou, L.; O’Rourke, B. From mitochondrial dynamics to arrhythmias. Int. J. Biochem. Cell Biol. 2009, 41, 1940–1948. [Google Scholar] [CrossRef] [Green Version]
- O’Rourke, B.; Blatter, L.A. Mitochondrial Ca2+ uptake: Tortoise or hare? J. Mol. Cell. Cardiol. 2009, 46, 767–774. [Google Scholar] [CrossRef] [Green Version]
- Setterberg, I.E.; Le, C.; Frisk, M.; Perdreau-Dahl, H.; Li, J.; Louch, W.E. The Physiology and Pathophysiology of T-Tubules in the Heart. Front. Physiol. 2021, 12, 1193. [Google Scholar] [CrossRef] [PubMed]
- Gadeberg, H.C.; Bond, R.C.; Kong, C.H.T.; Chanoit, G.P.; Ascione, R.; Cannell, M.B.; James, A.F. Heterogeneity of T-Tubules in Pig Hearts. PLoS ONE 2016, 11, e0156862. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.; Gorelik, J.; Yacoub, M.H.; Terracciano, C.M. The structure and function of cardiac t-tubules in health and disease. Proceedings Biol. Sci. 2011, 278, 2714–2723. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Treviño, P.; Pérez-Treviño, J.; Borja-Villa, C.; García, N.; Altamirano, J. Changes in T-Tubules and Sarcoplasmic Reticulum in Ventricular Myocytes in Early Cardiac Hypertrophy in a Pressure Overload Rat Model. Cell. Physiol. Biochem. 2015, 37, 1329–1344. [Google Scholar] [CrossRef] [PubMed]
- Schobesberger, S.; Wright, P.; Tokar, S.; Bhargava, A.; Mansfield, C.; Glukhov, A.V.; Poulet, C.; Buzuk, A.; Monszpart, A.; Sikkel, M.; et al. T-tubule remodelling disturbs localized β2-adrenergic signalling in rat ventricular myocytes during the progression of heart failure. Cardiovasc. Res. 2017, 113, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Liu, M.B.; Qu, Z. Transverse tubular network structures in the genesis of intracellular calcium alternans and triggered activity in cardiac cells. J. Mol. Cell. Cardiol. 2018, 114, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Nivala, M.; Ko, C.Y.; Nivala, M.; Weiss, J.N.; Qu, Z. Criticality in Intracellular Calcium Signaling in Cardiac Myocytes. Biophys. J. 2012, 102, 2433–2442. [Google Scholar] [CrossRef] [Green Version]
- Colman, M.A.; Pinali, C.; Trafford, A.W.; Zhang, H.; Kitmitto, A. A computational model of spatio-temporal cardiac intracellular calcium handling with realistic structure and spatial flux distribution from sarcoplasmic reticulum and t-tubule reconstructions. PLoS Comput. Biol. 2017, 13, e1005714. [Google Scholar] [CrossRef] [Green Version]
- Hoang-Trong, T.M.; Lederer, W.J.; Jafri, M.S. Exploring SR Calcium and Cytosolic Calcium Wave Dynamics using a 3D Stochastic Myocyte Model. Biophys. J. 2014, 106, 320a. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Feng, Y.; Huo, Y.; Tan, W. The Interplay of Rogue and Clustered Ryanodine Receptors Regulates Ca2+ Waves in Cardiac Myocytes. Front. Physiol. 2018, 9, 393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannell, M.B.; Kong, C.H.T.; Imtiaz, M.S.; Laver, D.R. Control of sarcoplasmic reticulum Ca2+ release by stochastic RyR gating within a 3D model of the cardiac dyad and importance of induction decay for CICR termination. Biophys. J. 2013, 104, 2149–2159. [Google Scholar] [CrossRef] [Green Version]
















| CRU Dist. | 0.6 | 0.8 | 1.0 | |
|---|---|---|---|---|
| RyR2 Sense | ||||
| [Ca2+]sr,max = 1.13 mM | 93.22% | 1.27% | 0.8% | |
| [Ca2+]sr,max = 1.3 mM | 95.48% | 1.55% | 1% | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoang-Trong, T.M.; Ullah, A.; Lederer, W.J.; Jafri, M.S. A Stochastic Spatiotemporal Model of Rat Ventricular Myocyte Calcium Dynamics Demonstrated Necessary Features for Calcium Wave Propagation. Membranes 2021, 11, 989. https://doi.org/10.3390/membranes11120989
Hoang-Trong TM, Ullah A, Lederer WJ, Jafri MS. A Stochastic Spatiotemporal Model of Rat Ventricular Myocyte Calcium Dynamics Demonstrated Necessary Features for Calcium Wave Propagation. Membranes. 2021; 11(12):989. https://doi.org/10.3390/membranes11120989
Chicago/Turabian StyleHoang-Trong, Tuan Minh, Aman Ullah, William Jonathan Lederer, and Mohsin Saleet Jafri. 2021. "A Stochastic Spatiotemporal Model of Rat Ventricular Myocyte Calcium Dynamics Demonstrated Necessary Features for Calcium Wave Propagation" Membranes 11, no. 12: 989. https://doi.org/10.3390/membranes11120989
APA StyleHoang-Trong, T. M., Ullah, A., Lederer, W. J., & Jafri, M. S. (2021). A Stochastic Spatiotemporal Model of Rat Ventricular Myocyte Calcium Dynamics Demonstrated Necessary Features for Calcium Wave Propagation. Membranes, 11(12), 989. https://doi.org/10.3390/membranes11120989