
Influence of Ceramic Membrane Surface Characteristics on the Flux Behavior of a Complex Fermentation Broth

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Fermentation Medium
2.2. Cultivation of K. lactis
2.3. Harvest of Medium and Biomass
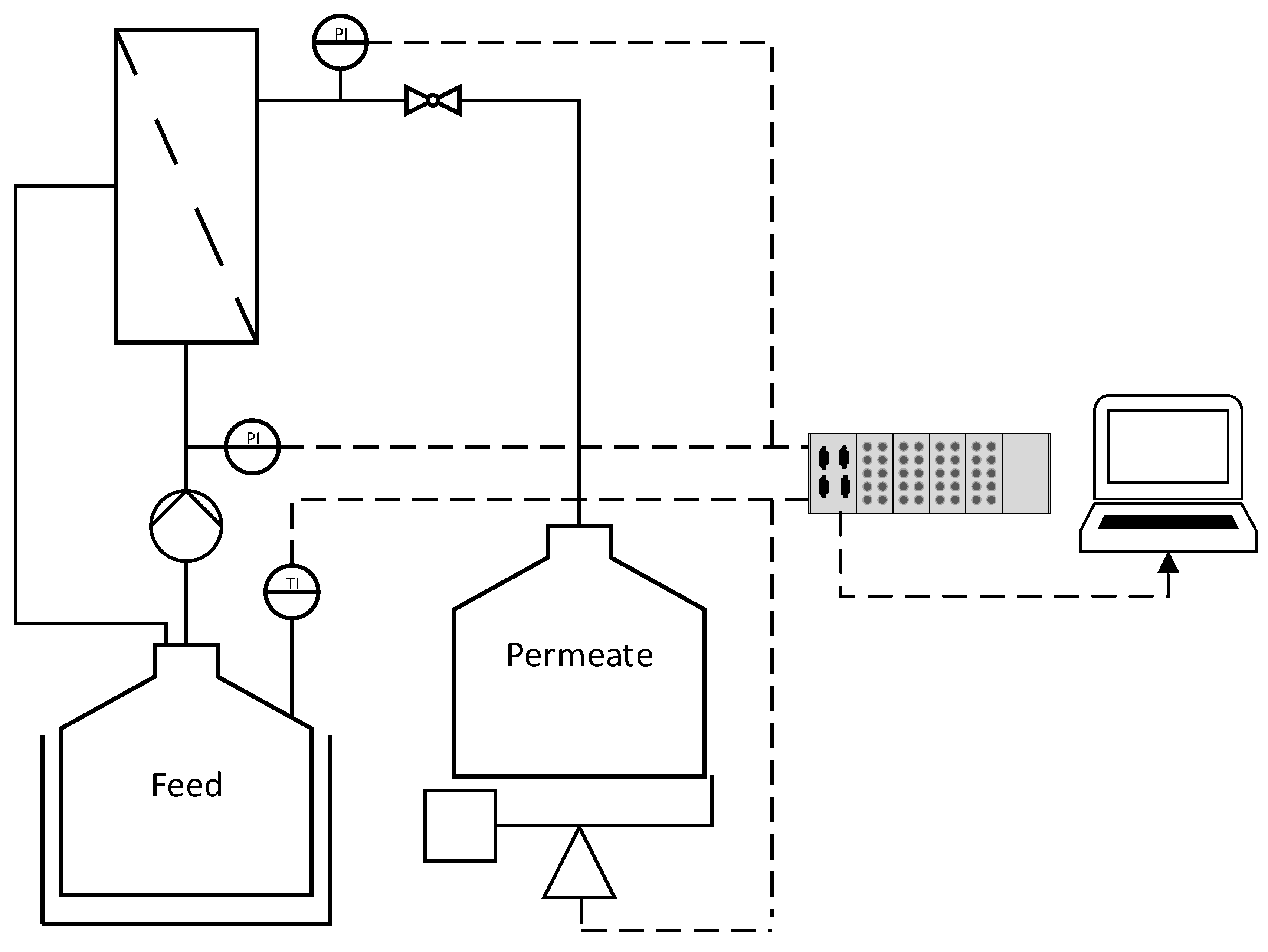
2.4. Filtration Setup
2.5. Estimation of Intrinsic Membrane Resistance and Irreversible Resistance
2.6. Investigation of Total, Fouling, and Reversible Resistance
2.7. Chemical Cleaning
2.8. Reynolds Number and Wall Shear Stress Calculations
2.9. Viscosity Measurement
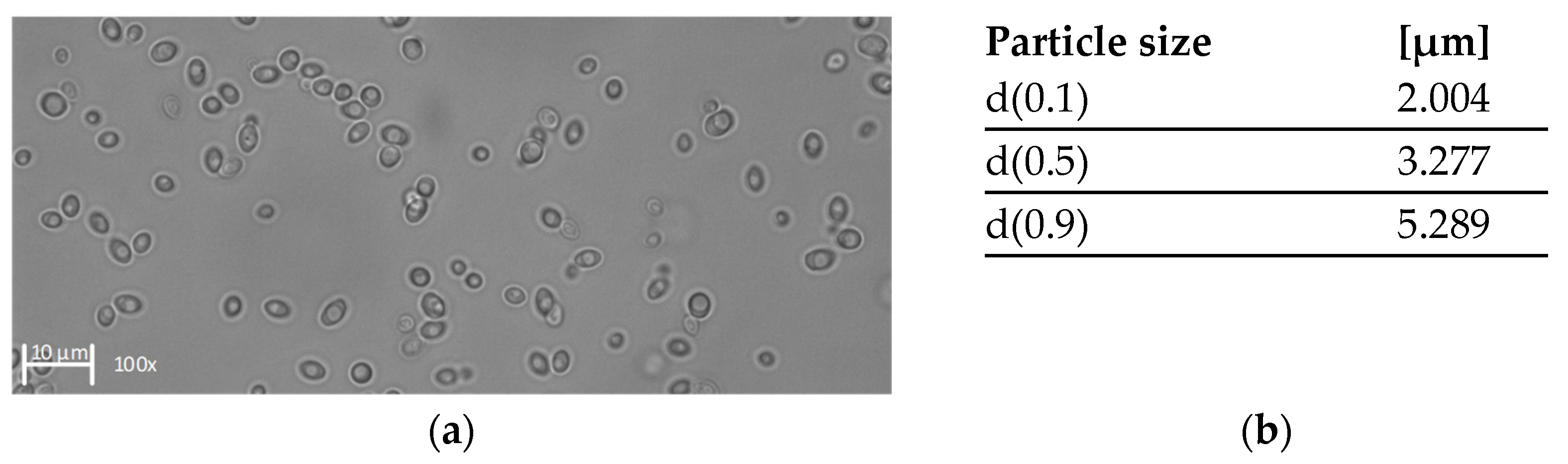
2.10. Particle Size Measurements
2.11. Measurement of Biomass Concentration
2.12. Bradford Assay
2.13. Surface Characterization of Membranes 7C_s and 7C_r
2.14. SEM Images for Fouling Analysis
3. Results
3.1. Characterization of the Filtration Feed
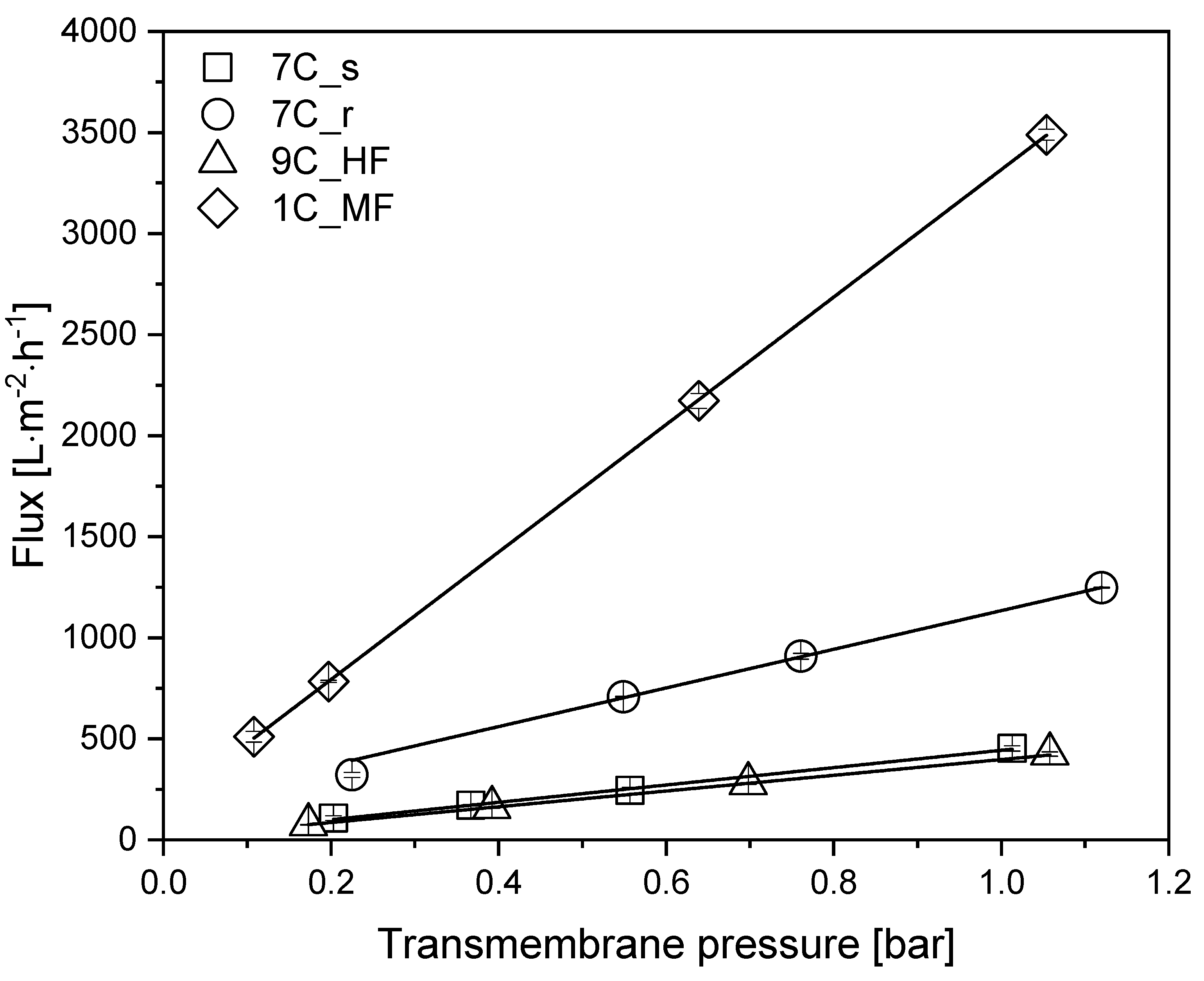
3.2. Comparison of Pure Water Fluxes
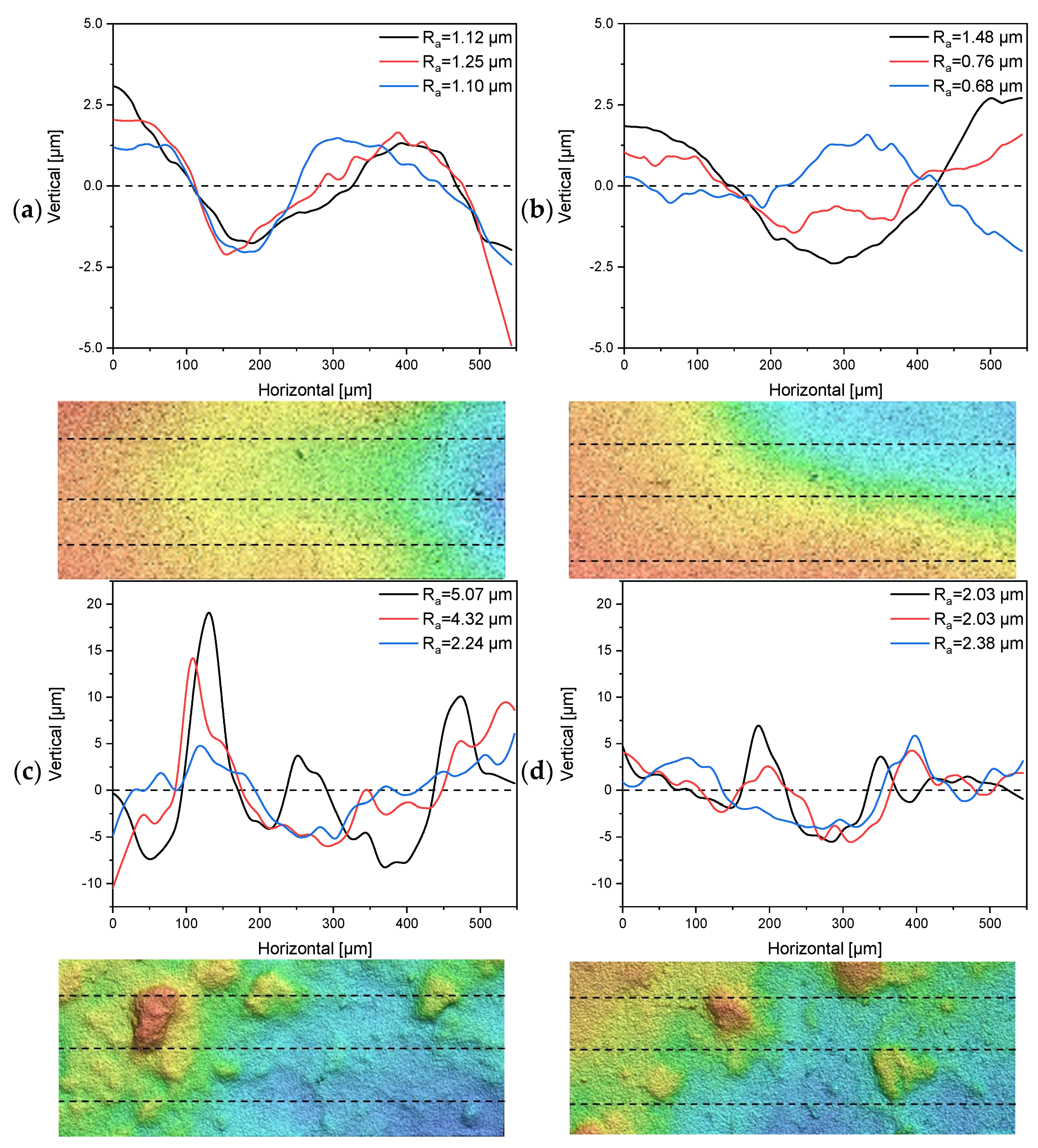
3.3. Characterization of Surface Roughness and the Separating Layers of Tubular UF Membranes
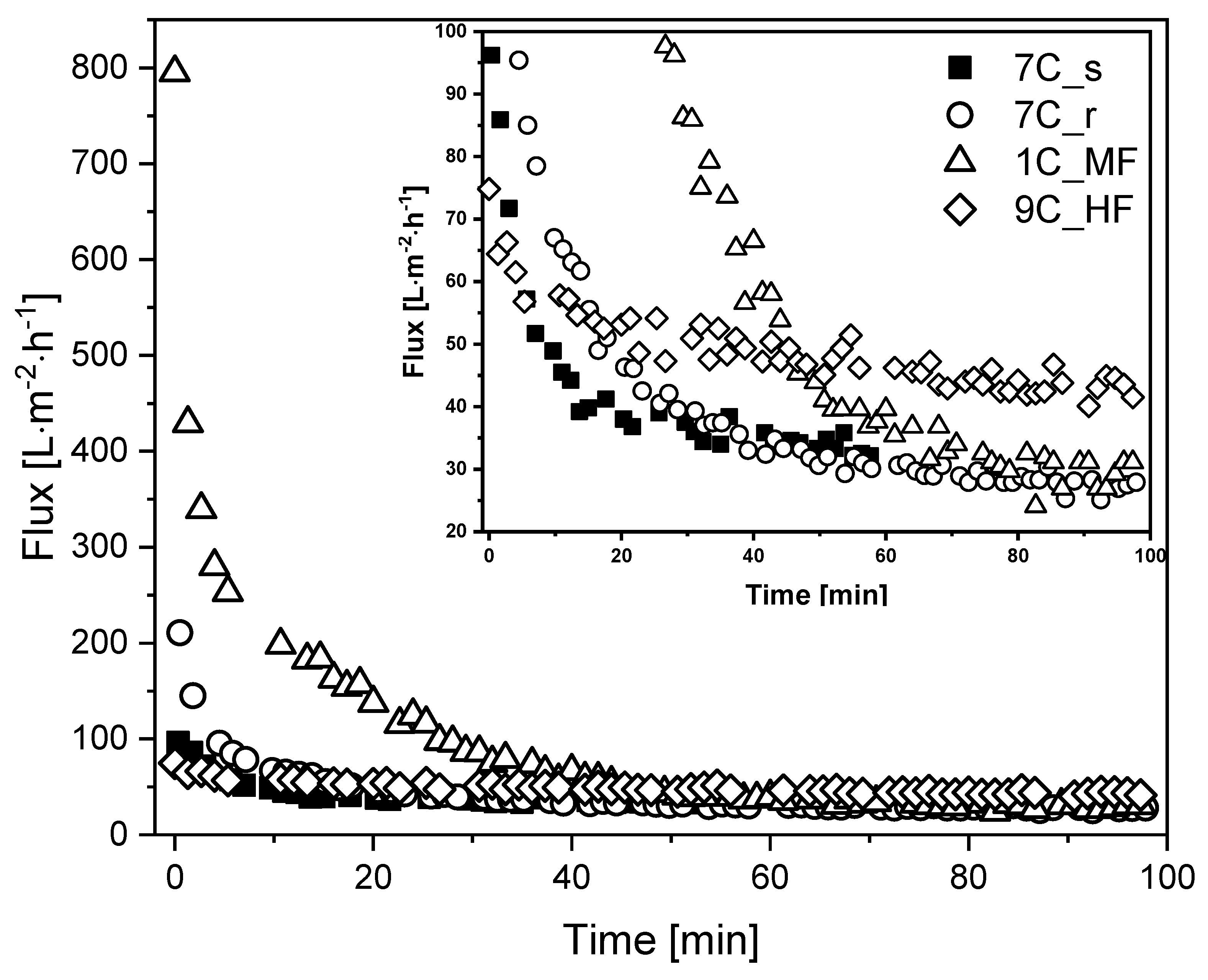
3.4. Performance of the Membranes with Fermentation Broth
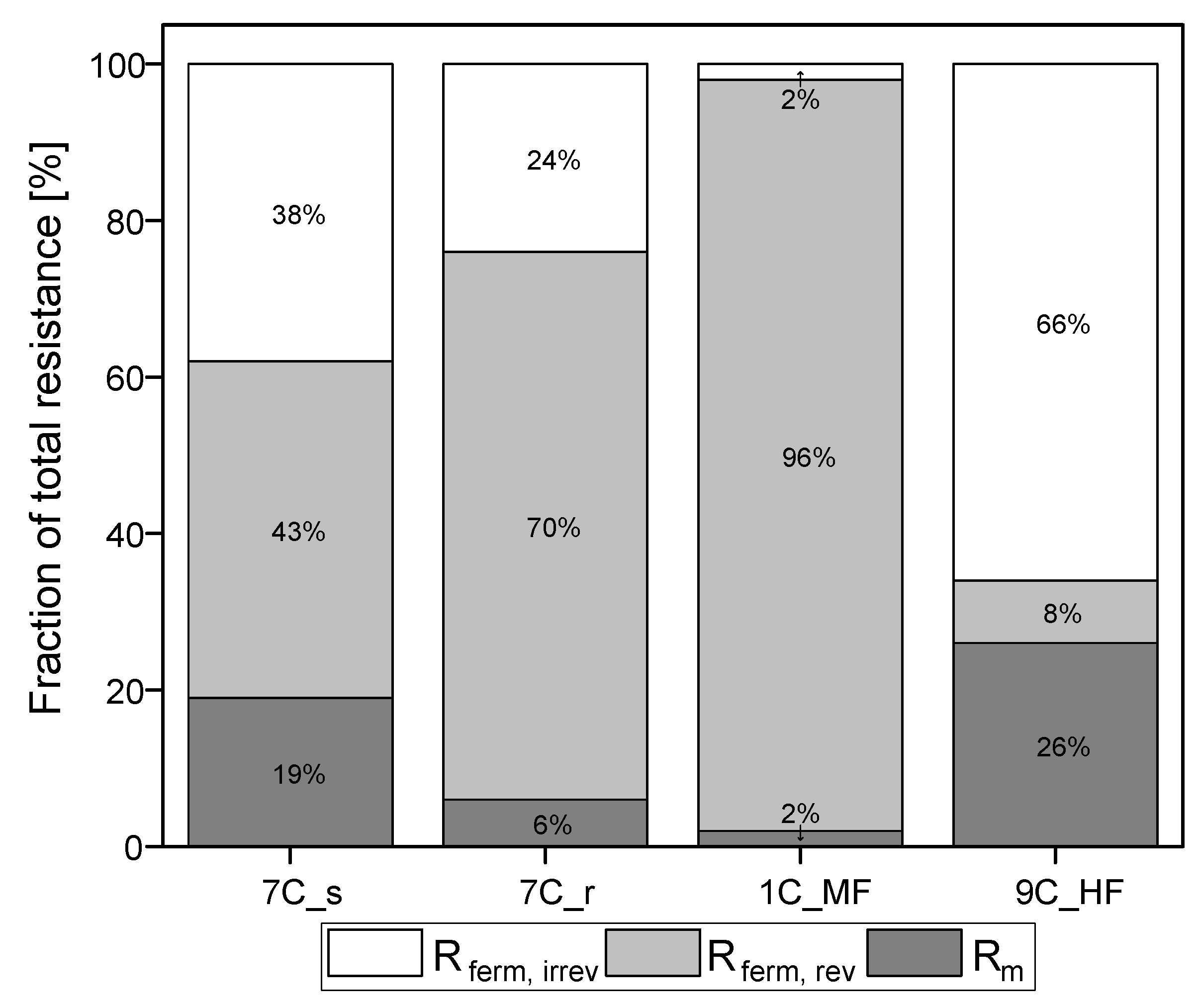
3.5. Calculated Resistances for the Filtration of Fermentation Broth
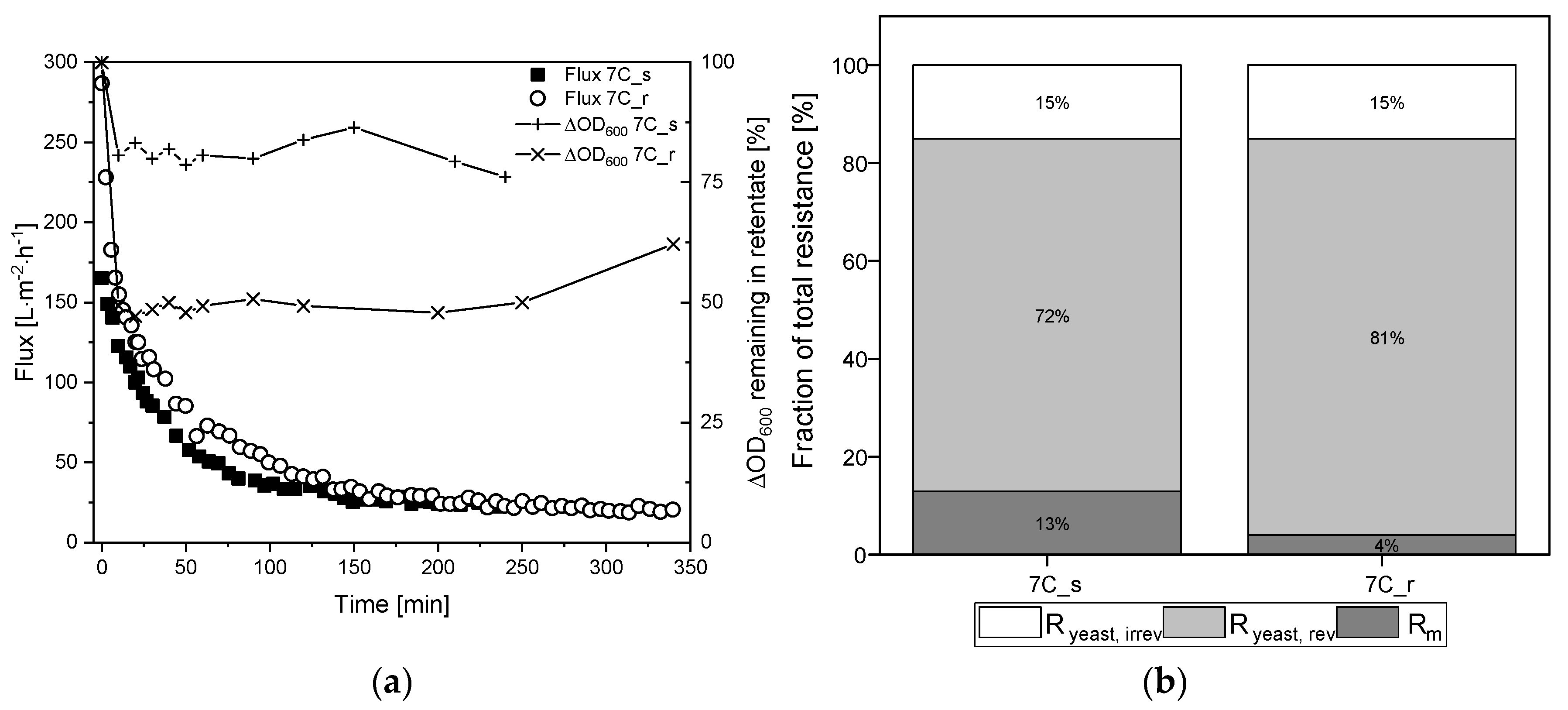
3.6. Filtration of Medium-Free K. lactis Cell Suspension
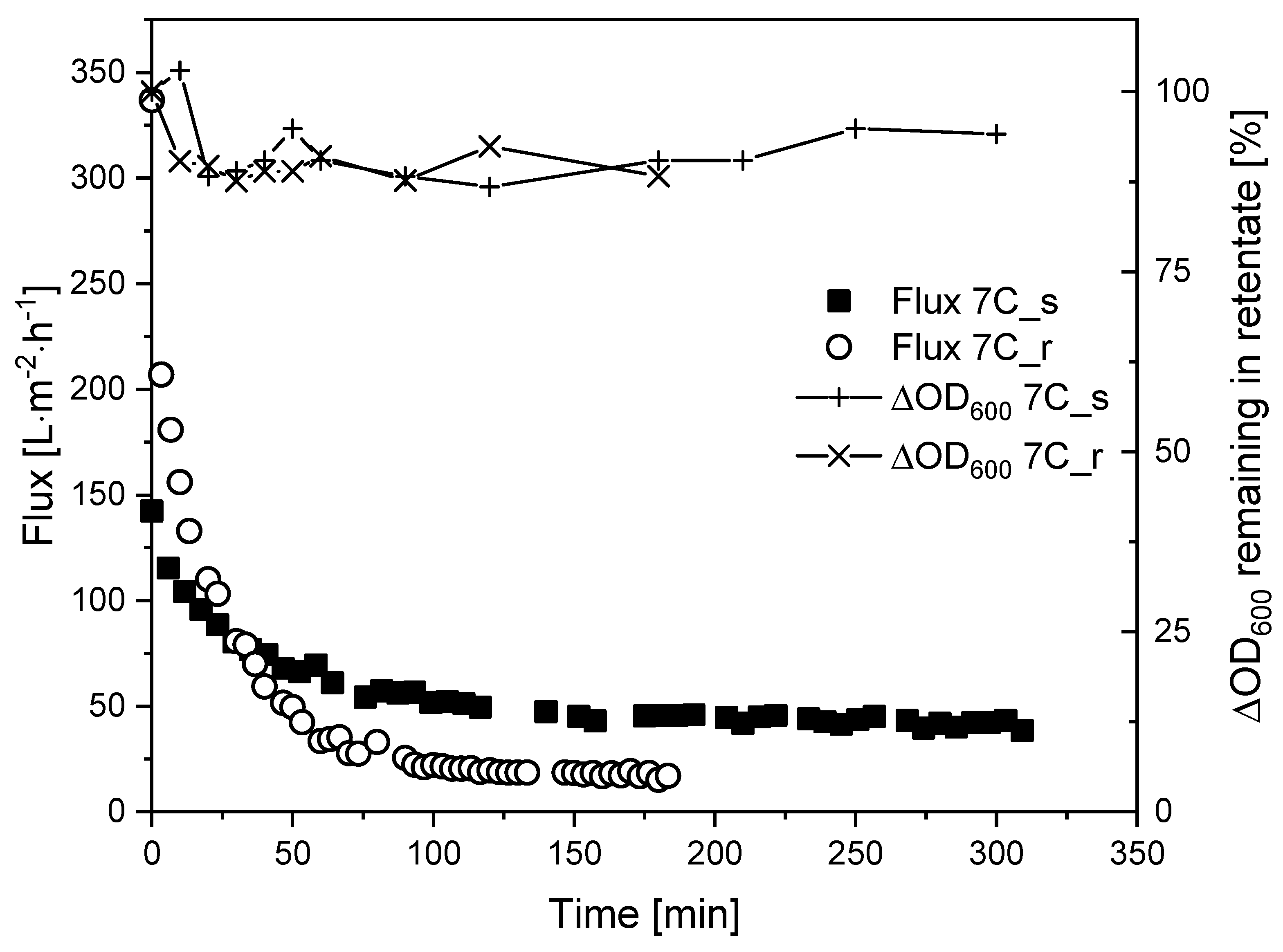
3.7. Fouling Control by Regulating the CFV
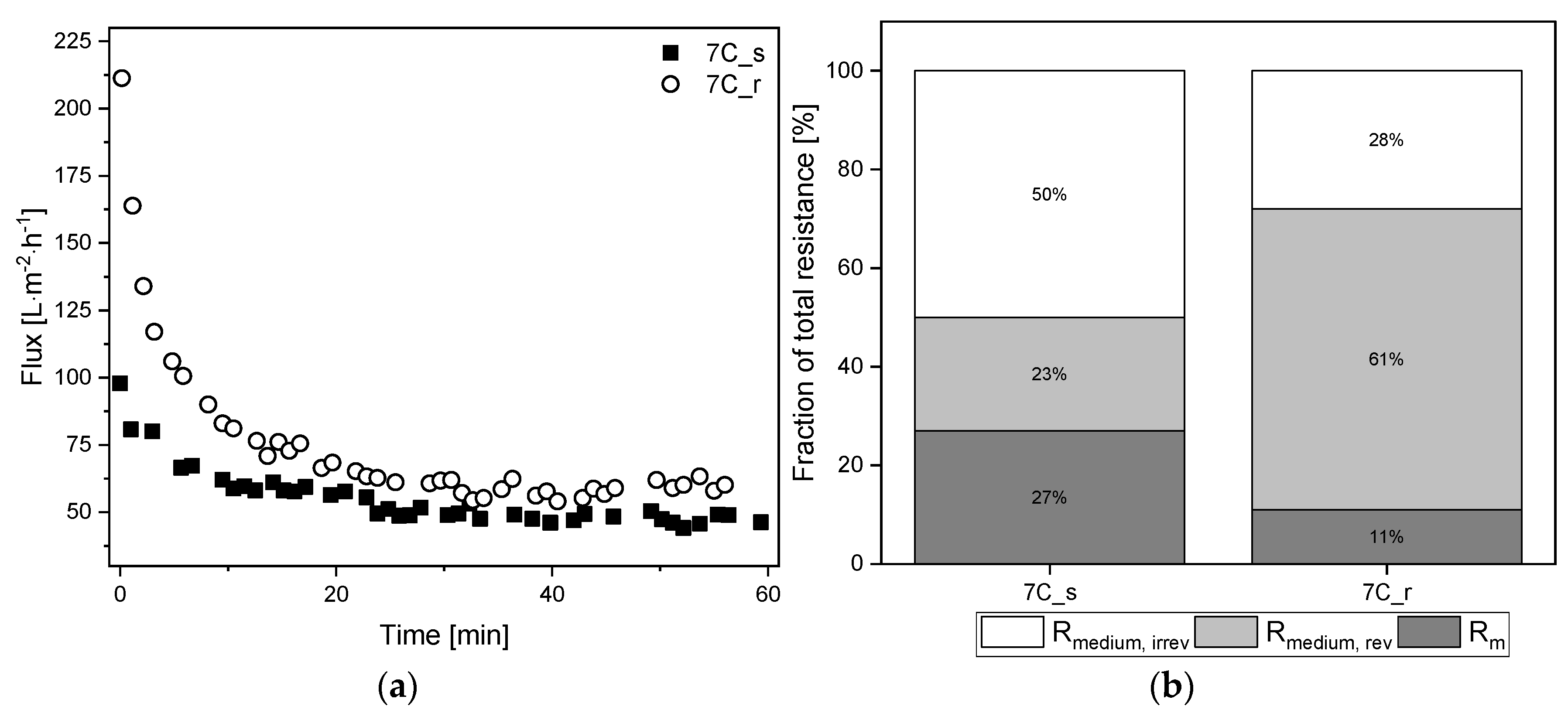
3.8. Filtration of Cell-Free Medium
3.9. Fouling Analysis by SEM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beltrán-Ramírez, F.; Orona-Tamayo, D.; Cornejo-Corona, I.; Luz Nicacio González-Cervantes, J.; de Jesús Esparza-Claudio, J.; Quintana-Rodríguez, E. Agro-Industrial Waste Revalorization: The Growing Biorefinery. In Biomass for Bioenergy-Recent Trends and Future Challenges; El-Fatah Abomohra, A., Ed.; IntechOpen: London, UK, 2019; ISBN 978-1-78923-987-4. [Google Scholar]
- Spohner, S.C.; Schaum, V.; Quitmann, H.; Czermak, P. Kluyveromyces lactis: An emerging tool in biotechnology. J. Biotechnol. 2016, 222, 104–116. [Google Scholar] [CrossRef]
- Jornitz, M.W.; Meltzer, T.H. Sterilizing filtrations with microporous membranes. Pharmacop. Forum 2004, 30, 1903–1910. [Google Scholar]
- Wang, A.; Lewus, R.; Rathore, A.S. Comparison of different options for harvest of a therapeutic protein product from high cell density yeast fermentation broth. Biotechnol. Bioeng. 2006, 94, 91–104. [Google Scholar] [CrossRef]
- Ciora, R.J.; Liu, P.K.T. Ceramic membranes for environmental related applications. Fluid-Part. Sep. J. 2003, 15, 51–60. [Google Scholar]
- Gitis, V.; Rothenberg, G. (Eds.) Economics. In Ceramic Membranes: New Opportunities and Practical Applications; Wiley: Hoboken, NJ, USA, 2016; pp. 355–388. ISBN 9783527334933. [Google Scholar]
- Gruskevica, K.; Mezule, L. Cleaning Methods for Ceramic Ultrafiltration Membranes Affected by Organic Fouling. Membranes 2021, 11, 131. [Google Scholar] [CrossRef]
- Tang, C.Y.; Chong, T.H.; Fane, A.G. Colloidal interactions and fouling of NF and RO membranes: A review. Adv. Colloid Interface Sci. 2011, 164, 126–143. [Google Scholar] [CrossRef] [PubMed]
- van der Bruggen, B. Influence of MF pretreatment on NF performance for aqueous solutions containing particles and an organic foulant. Sep. Purif. Technol. 2004, 36, 203–213. [Google Scholar] [CrossRef]
- Dizge, N.; Soydemir, G.; Karagunduz, A.; Keskinler, B. Influence of type and pore size of membranes on cross flow microfiltration of biological suspension. J. Membr. Sci. 2011, 366, 278–285. [Google Scholar] [CrossRef]
- Zhong, Z.; Li, D.; Zhang, B.; Xing, W. Membrane surface roughness characterization and its influence on ultrafine particle adhesion. Sep. Purif. Technol. 2012, 90, 140–146. [Google Scholar] [CrossRef]
- Qin, W.; Zhang, Y.; Wu, J. Preparation of high-permeance ceramic microfiltration membranes using a pore-sealing method. RSC Adv. 2020, 10, 5560–5565. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Fan, Y.; Qi, H.; Xu, N. Co-sintering synthesis of tubular bilayer α-alumina membrane. J. Membr. Sci. 2007, 288, 20–27. [Google Scholar] [CrossRef]
- Drews, A. Membrane fouling in membrane bioreactors—Characterisation, contradictions, cause and cures. J. Membr. Sci. 2010, 363, 1–28. [Google Scholar] [CrossRef]
- Kujundzic, E.; Greenberg, A.R.; Fong, R.; Moore, B.; Kujundzic, D.; Hernandez, M. Biofouling potential of industrial fermentation broth components during microfiltration. J. Membr. Sci. 2010, 349, 44–55. [Google Scholar] [CrossRef]
- Hassan, I.B.; Ennouri, M.; Lafforgue, C.; Schmitz, P.; Ayadi, A. Experimental Study of Membrane Fouling during Crossflow Microfiltration of Yeast and Bacteria Suspensions: Towards an Analysis at the Microscopic Level. Membranes 2013, 3, 44–68. [Google Scholar] [CrossRef] [Green Version]
- Hashino, M.; Hirami, K.; Ishigami, T.; Ohmukai, Y.; Maruyama, T.; Kubota, N.; Matsuyama, H. Effect of kinds of membrane materials on membrane fouling with BSA. J. Membr. Sci. 2011, 384, 157–165. [Google Scholar] [CrossRef]
- de Roever, E.W.; Huisman, I.H. Microscopy as a tool for analysis of membrane failure and fouling. Desalination 2007, 207, 35–44. [Google Scholar] [CrossRef]
- Burghardt, J.P.; Coletta, L.A.; van der Bolt, R.; Ebrahimi, M.; Gerlach, D.; Czermak, P. Development and Characterization of an Enzyme Membrane Reactor for Fructo-Oligosaccharide Production. Membranes 2019, 9, 148. [Google Scholar] [CrossRef] [Green Version]
- Birrenbach, O.; Faust, F.; Ebrahimi, M.; Fan, R.; Czermak, P. Recovery and Purification of Protein Aggregates From Cell Lysates Using Ceramic Membranes: Fouling Analysis and Modeling of Ultrafiltration. Front. Chem. Eng. 2021, 3. [Google Scholar] [CrossRef]
- Loewe, D.; Grein, T.A.; Dieken, H.; Weidner, T.; Salzig, D.; Czermak, P. Tangential Flow Filtration for the Concentration of Oncolytic Measles Virus: The Influence of Filter Properties and the Cell Culture Medium. Membranes 2019, 9, 160. [Google Scholar] [CrossRef] [Green Version]
- Fan, R.; Ebrahimi, M.; Quitmann, H.; Czermak, P. Lactic acid production in a membrane bioreactor system with thermophilic Bacillus coagulans: Fouling analysis of the used ceramic membranes. Sep. Sci. Technol. 2015, 150527095459001. [Google Scholar] [CrossRef]
- Li, H.; Chen, V. Membrane Fouling and Cleaning in Food and Bioprocessing. In Membrane Technology; Elsevier: Amsterdam, The Netherlands, 2010; pp. 213–254. ISBN 9781856176323. [Google Scholar]
- Gençal, Y.; Durmaz, E.N.; Çulfaz-Emecen, P.Z. Preparation of patterned microfiltration membranes and their performance in crossflow yeast filtration. J. Membr. Sci. 2015, 476, 224–233. [Google Scholar] [CrossRef]
- Böswirth, L.; Bschorer, S. Technische Strömungslehre; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Zor, T.; Selinger, Z. Linearization of the Bradford protein assay increases its sensitivity: Theoretical and experimental studies. Anal. Biochem. 1996, 236, 302–308. [Google Scholar] [CrossRef] [Green Version]
- Zakhartsev, M.; Reuss, M. Cell size and morphological properties of yeast Saccharomyces cerevisiae in relation to growth temperature. FEMS Yeast Res. 2018, 18. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Guan, K.; Gao, P.; Peng, C.; Wu, J. A preparation method for the highly permeable ceramic microfiltration membrane–precursor film firing method. RSC Adv. 2018, 8, 2906–2914. [Google Scholar] [CrossRef] [Green Version]
- Tran-Duc, T.; Phan-Thien, N.; Wang, J. A theoretical study of permeability enhancement for ultrafiltration ceramic membranes with conical pores and slippage. Phys. Fluids 2019, 31, 22003. [Google Scholar] [CrossRef]
- Piry, A.; Heino, A.; Kühnl, W.; Grein, T.; Ripperger, S.; Kulozik, U. Effect of membrane length, membrane resistance, and filtration conditions on the fractionation of milk proteins by microfiltration. J. Dairy Sci. 2012, 95, 1590–1602. [Google Scholar] [CrossRef] [PubMed]
- Gitis, V.; Rothenberg, G. (Eds.) The Basics. In Ceramic Membranes: New Opportunities and Practical Applications; Wiley: Hoboken, NJ, USA, 2016; pp. 1–90. ISBN 9783527334933. [Google Scholar]
- Luque, S.; Gómez, D.; Álvarez, J.R. Industrial Applications of Porous Ceramic Membranes (Pressure-Driven Processes). In Inorganic Membranes: Synthesis, Characterization and Applications; Mallada, R., Menéndez, M., Eds.; Elsevier: Amsterdam, The Netherlands; Oxford, UK, 2013; pp. 177–216. ISBN 9780444530707. [Google Scholar]
- Bonekamp, B.C. Chapter 6 Preparation of asymmetric ceramic membrane supports by dip-coating. In Membrane Science and Technology: Fundamentals of Inorganic Membrane Science and Technology; Burggraaf, A.J., Cot, L., Eds.; Elsevier: Amsterdam, The Netherlands, 1996; pp. 141–225. ISBN 0927-5193. [Google Scholar]
- Homayoonfal, M.; Akbari, A.; Mehrnia, M.R. Preparation of polysulfone nanofiltration membranes by UV-assisted grafting polymerization for water softening. Desalination 2010, 263, 217–225. [Google Scholar] [CrossRef]
- Humpert, D.; Ebrahimi, M.; Stroh, A.; Czermak, P. Recovery of Lignosulfonates from Spent Sulfite Liquor Using Ceramic Hollow-Fiber Membranes. Membranes 2019, 9, 45. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-N.; Tang, C.Y. Protein fouling of nanofiltration, reverse osmosis, and ultrafiltration membranes—The role of hydrodynamic conditions, solution chemistry, and membrane properties. J. Membr. Sci. 2011, 376, 275–282. [Google Scholar] [CrossRef]
- Chan, C.C.V.; Bérubé, P.R.; Hall, E.R. Relationship between types of surface shear stress profiles and membrane fouling. Water Res. 2011, 45, 6403–6416. [Google Scholar] [CrossRef] [PubMed]
- Delgado, S.; Villarroel, R.; González, E. Effect of the shear intensity on fouling in submerged membrane bioreactor for wastewater treatment. J. Membr. Sci. 2008, 311, 173–181. [Google Scholar] [CrossRef]
- Hughes, D.; Field, R.W. Crossflow filtration of washed and unwashed yeast suspensions at constant shear under nominally sub-critical conditions. J. Membr. Sci. 2006, 280, 89–98. [Google Scholar] [CrossRef]
- Güell, C.; Czekaj, P.; Davis, R.H. Microfiltration of protein mixtures and the effects of yeast on membrane fouling. J. Membr. Sci. 1999, 155, 113–122. [Google Scholar] [CrossRef]
- Kuberkar, V.T.; Davis, R.H. Effects of added yeast on protein transmission and flux in cross-flow membrane microfiltration. Biotechnol. Prog. 1999, 15, 472–479. [Google Scholar] [CrossRef]
- Choi, H.; Zhang, K.; Dionysiou, D.D.; Oerther, D.B.; Sorial, G.A. Influence of cross-flow velocity on membrane performance during filtration of biological suspension. J. Membr. Sci. 2005, 248, 189–199. [Google Scholar] [CrossRef]
- Zhong, Z.; Xing, W.; Zhang, B. Fabrication of ceramic membranes with controllable surface roughness and their applications in oil/water separation. Ceram. Int. 2013, 39, 4355–4361. [Google Scholar] [CrossRef]
- Panda, S.R.; Bhandaru, N.; Mukherjee, R.; De, S. Ultrafiltration of oily waste water: Contribution of surface roughness in membrane properties and fouling characteristics of polyacrylonitrile membranes. Can. J. Chem. Eng. 2015, 93, 2031–2042. [Google Scholar] [CrossRef]
- Vrijenhoek, E.M.; Hong, S.; Elimelech, M. Influence of membrane surface properties on initial rate of colloidal fouling of reverse osmosis and nanofiltration membranes. J. Membr. Sci. 2001, 188, 115–128. [Google Scholar] [CrossRef]
- Woo, S.H.; Park, J.; Min, B.R. Relationship between permeate flux and surface roughness of membranes with similar water contact angle values. Sep. Purif. Technol. 2015, 146, 187–191. [Google Scholar] [CrossRef]
- Hofs, B.; Ogier, J.; Vries, D.; Beerendonk, E.F.; Cornelissen, E.R. Comparison of ceramic and polymeric membrane permeability and fouling using surface water. Sep. Purif. Technol. 2011, 79, 365–374. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | Membrane | |||
|---|---|---|---|---|
| 7C_s | 7C_r | 1C MF | 9C HF | |
| Geometry | tubular | tubular | tubular | hollow fiber |
| Support material | Al2O3 | Al2O3/TiO2 | Al2O3 | Al2O3 |
| Separating layers | Al2O3 | Al2O3 | Al2O3 | Al2O3 |
| Cut-off [nm] | 30 | 30 | 200 | 30 |
| Length [mm] | 225 | 230 | 225 | 200 |
| Number of channels | 7 | 7 | 1 | 9 |
| Inner diameter of each channel [mm] | 6 | 6 | 6 | 2 |
| Filtration area [m2] | 2.97 × 10−2 | 3.03 × 10−2 | 0.42 × 10−2 | 1.13 × 10−2 |
| Cross-flow area [m2] | 1.98 × 10−4 | 1.98 × 10−4 | 2.83 × 10−5 | 2.83 × 10−5 |
| Solution | Viscosity [mPa∙s] |
|---|---|
| Medium + yeast cells | 1.17 |
| Yeast cells (2.5 g/L) | 1.11 |
| Medium | 1.12 |
| Pure water | 0.89 |
| Property | Membrane | |
|---|---|---|
| 7C_s | 7C_r | |
| Number of layers | 3 | 2 |
| Thickness of first layer | 9.57 ± 1.37 µm | 2.51 ± 0.49 µm |
| Thickness of second layer | 26.41 ± 1.99 µm | 43.42 ± 6.19 µm |
| Thickness of third layer | 49.09 ± 2.80 µm | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maguire, N.A.P.; Ebrahimi, M.; Fan, R.; Gießelmann, S.; Ehlen, F.; Schütz, S.; Czermak, P. Influence of Ceramic Membrane Surface Characteristics on the Flux Behavior of a Complex Fermentation Broth. Membranes 2021, 11, 402. https://doi.org/10.3390/membranes11060402
Maguire NAP, Ebrahimi M, Fan R, Gießelmann S, Ehlen F, Schütz S, Czermak P. Influence of Ceramic Membrane Surface Characteristics on the Flux Behavior of a Complex Fermentation Broth. Membranes. 2021; 11(6):402. https://doi.org/10.3390/membranes11060402
Chicago/Turabian StyleMaguire, Nicolas A. P., Mehrdad Ebrahimi, Rong Fan, Sabine Gießelmann, Frank Ehlen, Steffen Schütz, and Peter Czermak. 2021. "Influence of Ceramic Membrane Surface Characteristics on the Flux Behavior of a Complex Fermentation Broth" Membranes 11, no. 6: 402. https://doi.org/10.3390/membranes11060402
APA StyleMaguire, N. A. P., Ebrahimi, M., Fan, R., Gießelmann, S., Ehlen, F., Schütz, S., & Czermak, P. (2021). Influence of Ceramic Membrane Surface Characteristics on the Flux Behavior of a Complex Fermentation Broth. Membranes, 11(6), 402. https://doi.org/10.3390/membranes11060402