


New Aspects of Bilayer Lipid Membranes for the Analysis of Ion Channel Functions

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
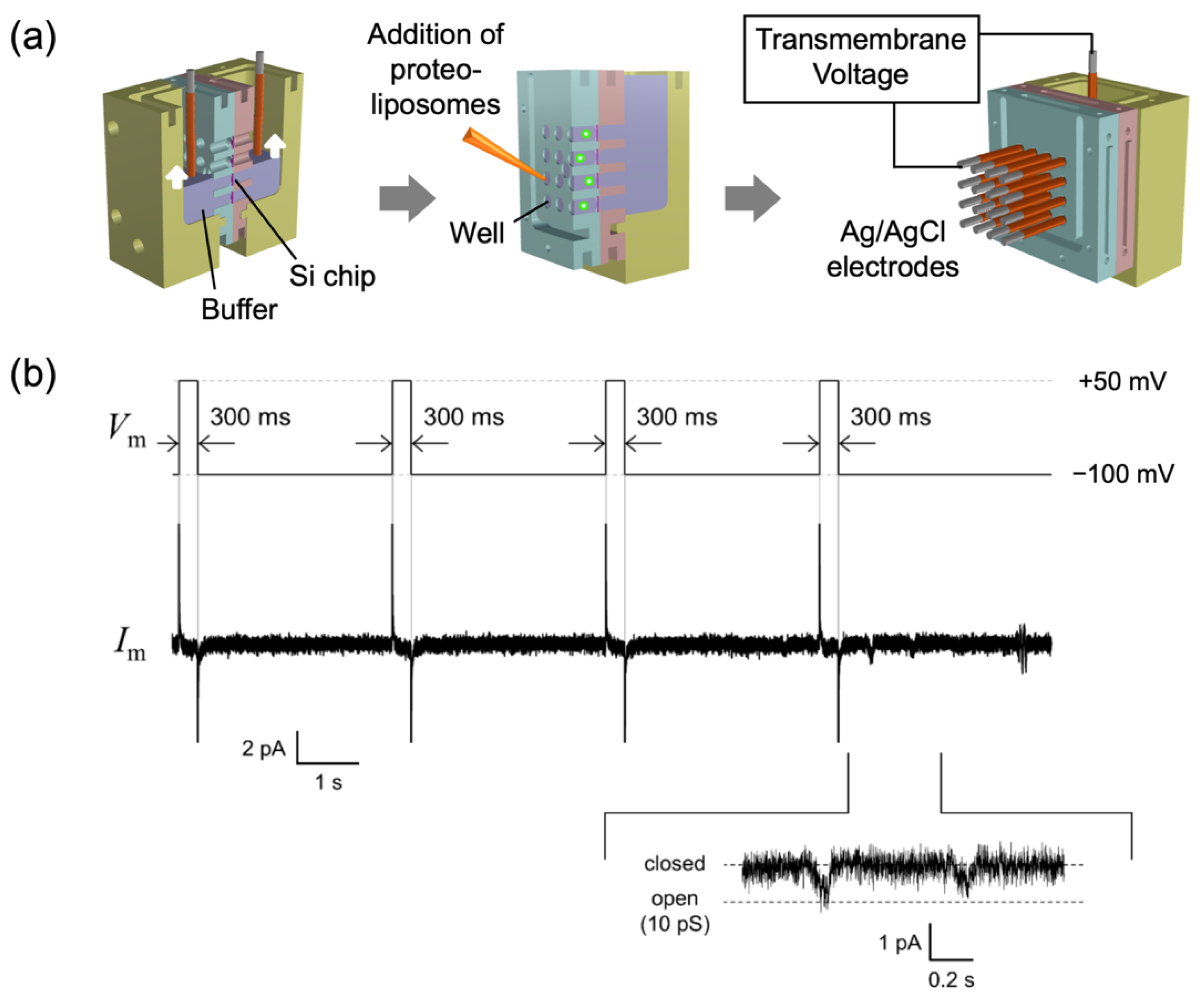{kind=link}
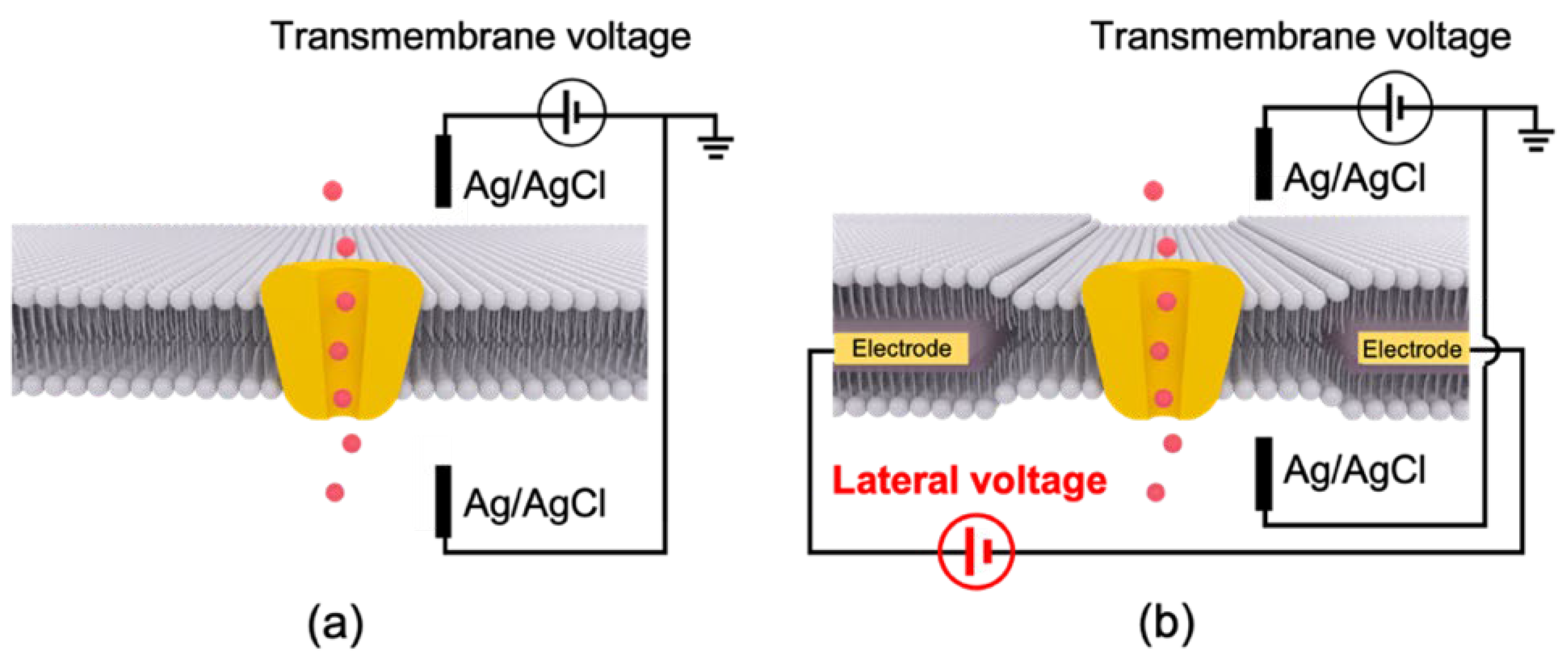{kind=link}
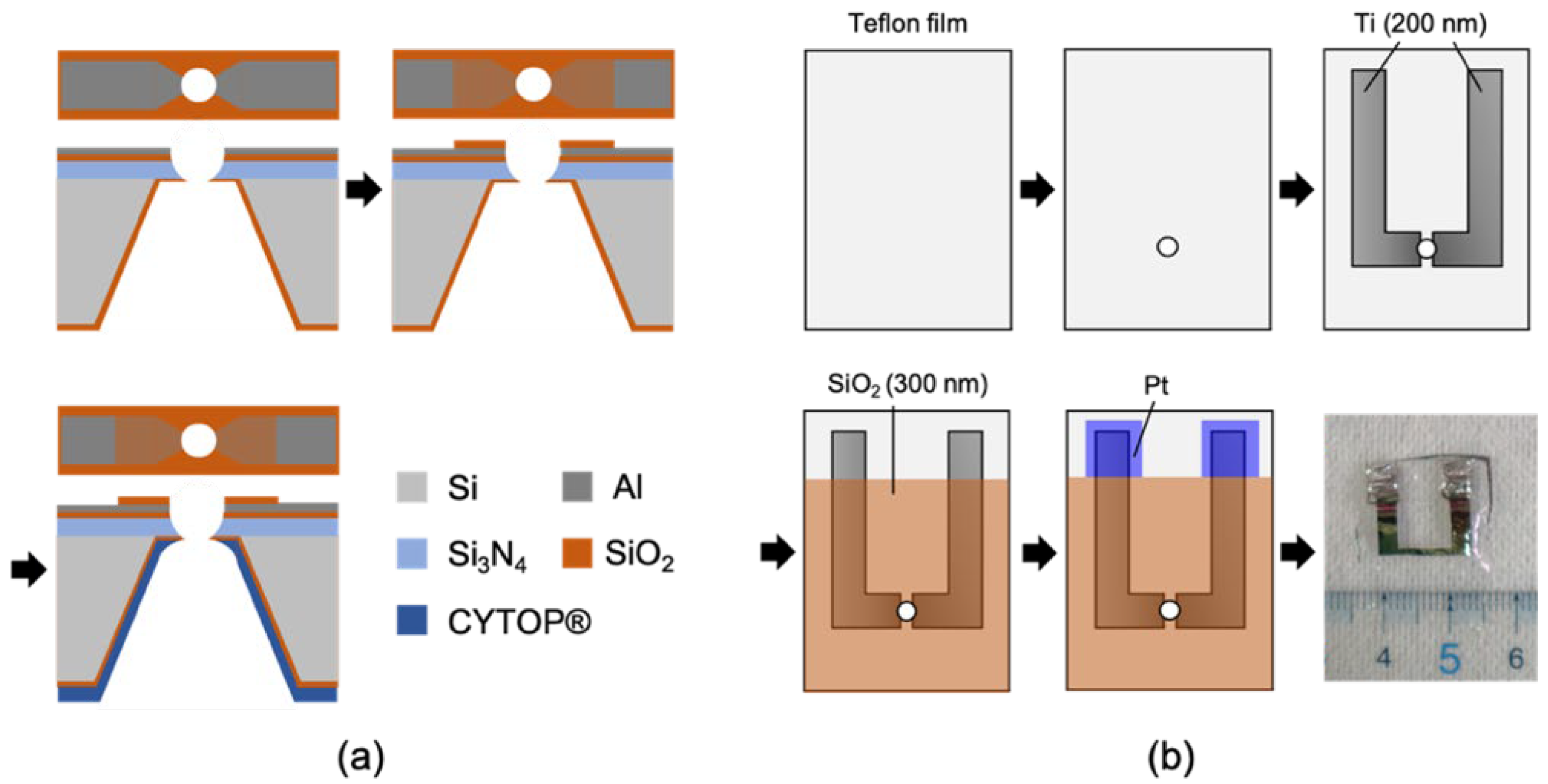{kind=link}
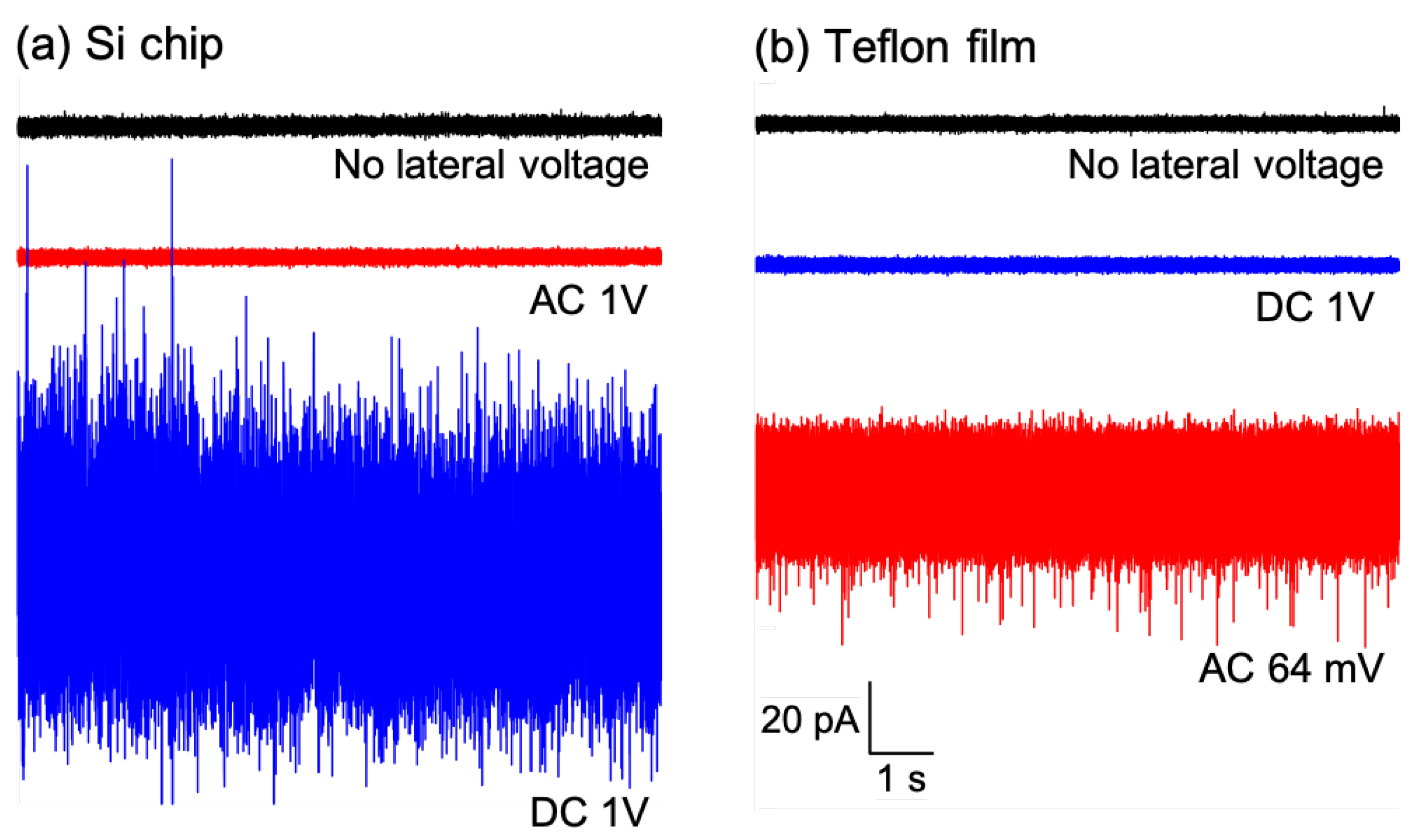{kind=link}
{kind=link}
Abstract
:1. Introduction
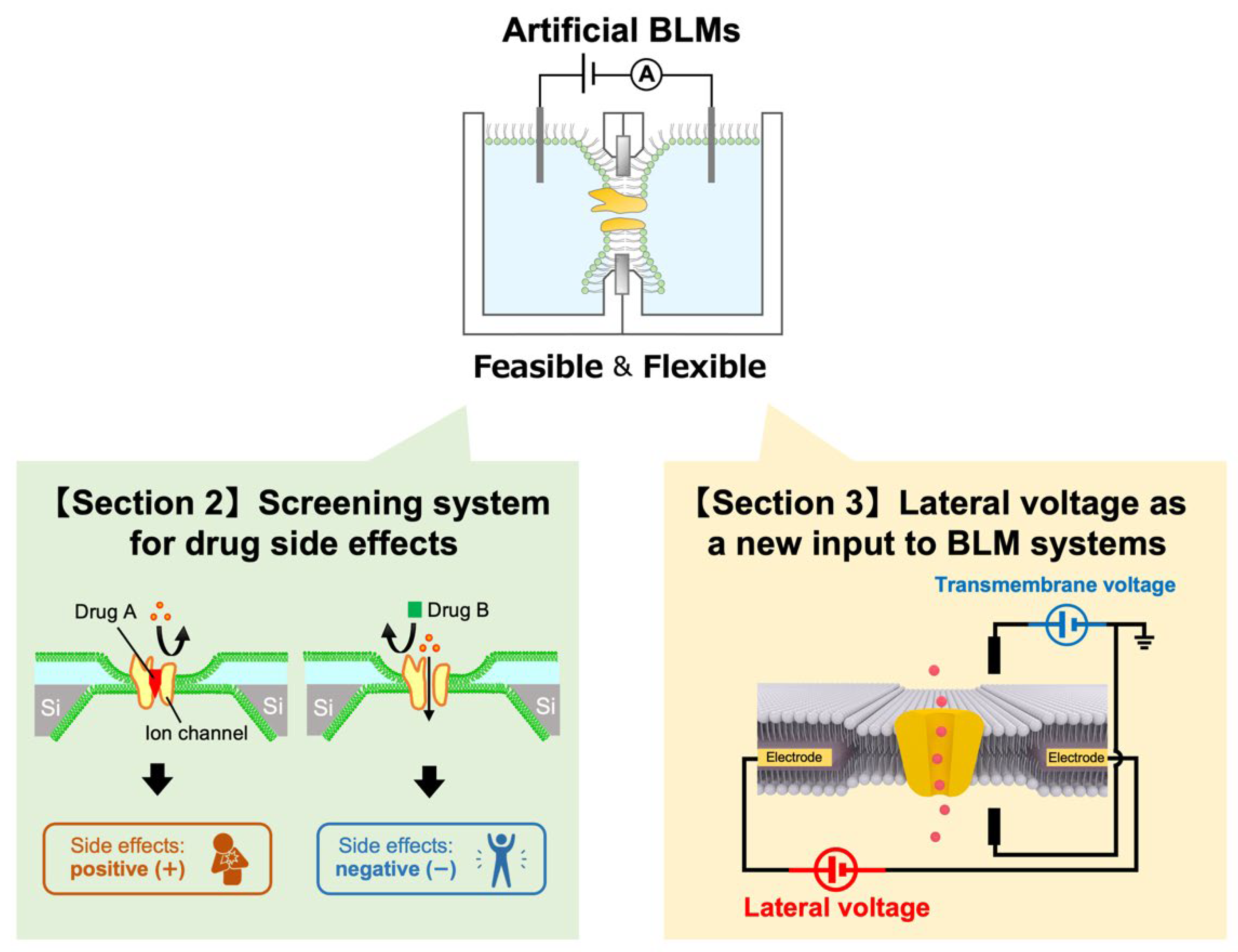
2. BLM-Based Screening Systems for Examining Drug Side Effects and Their Prospect for Personalized Medicine
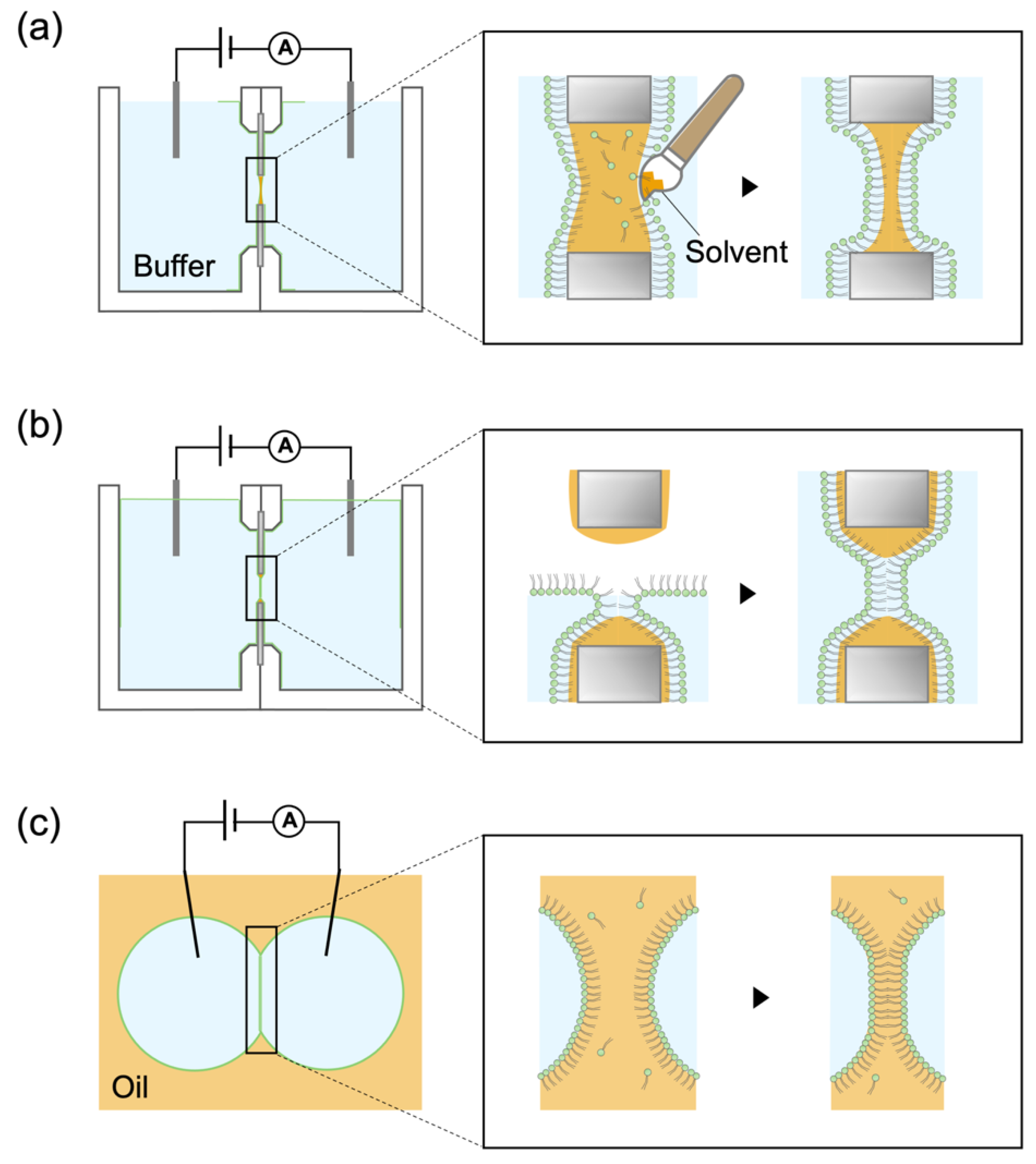
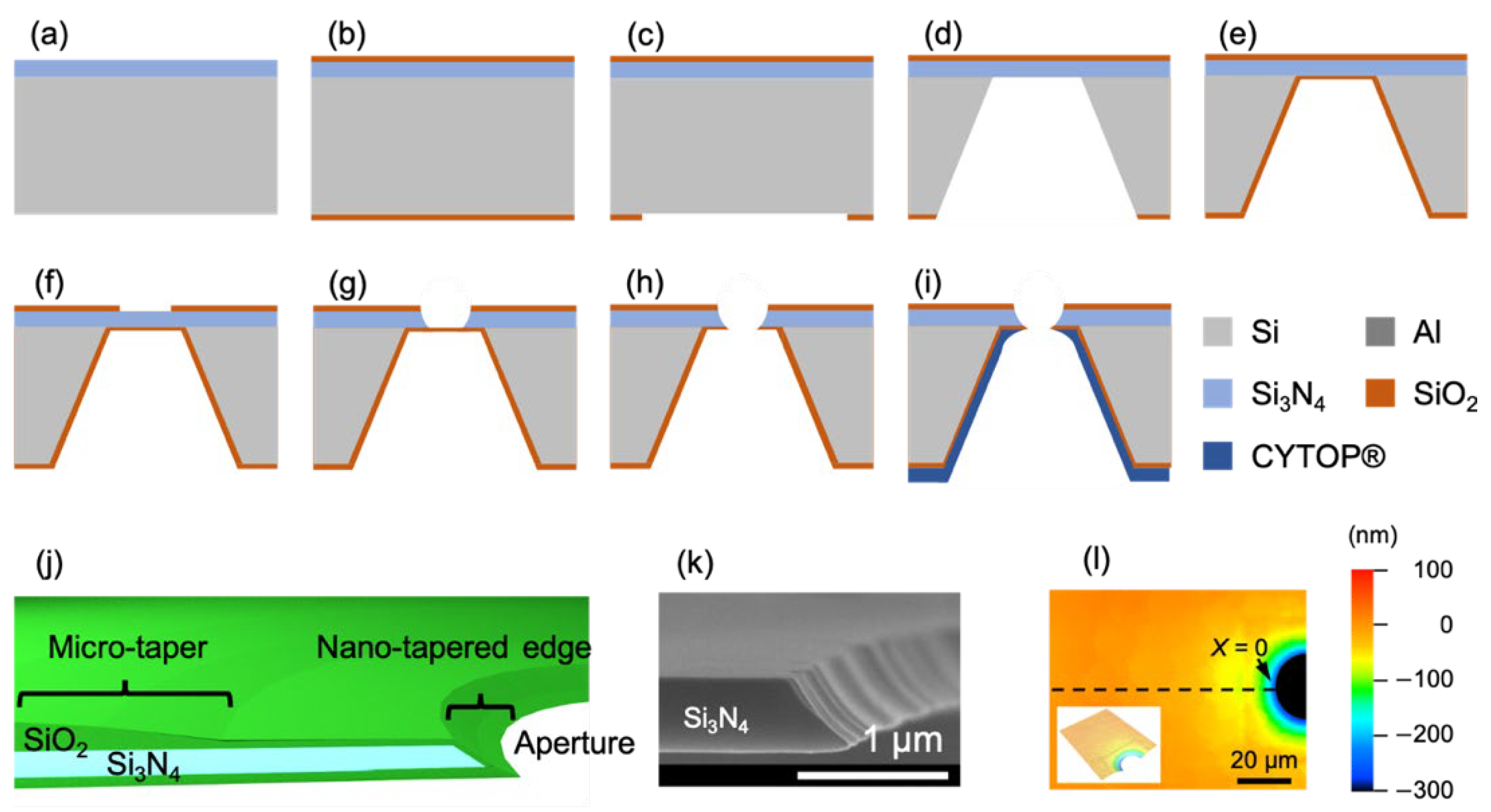
2.1. Fabrication of Tapered Apertures to Form Stable Solvent-Free BLMs
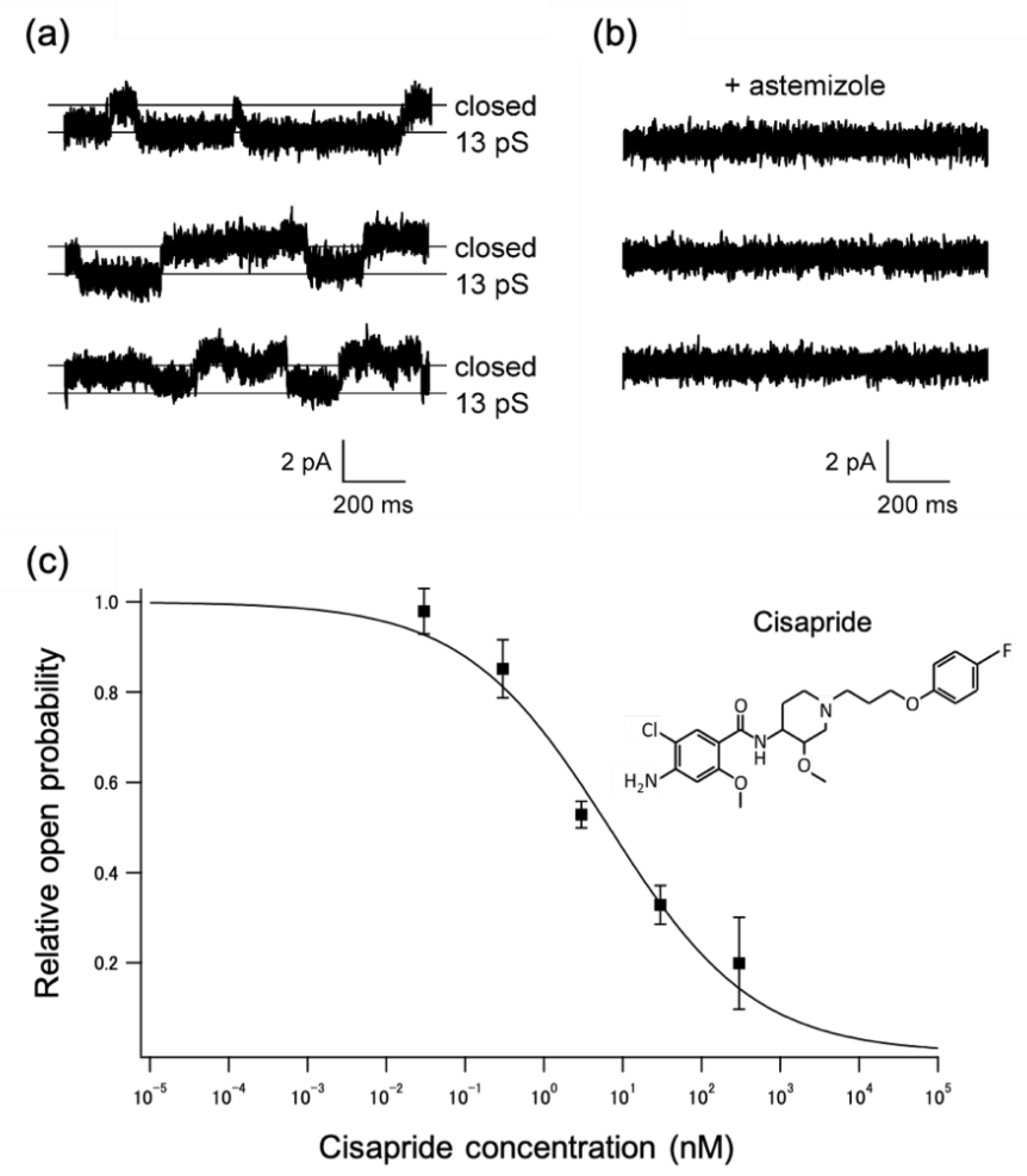
2.2. Applications to a Drug-Screening Platform
3. Lateral Voltage as a New Input to BLM Systems
3.1. Fabrication of Electrode-Wired Membrane Support for the Application of Lateral Voltage
3.2. BLM Formation in Electrode-Wired Membrane Supports
3.3. Effect of a Lateral Voltage on the Activities of Ion Channels Embedded in BLMs
4. Summary and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tomioka, Y.; Takashima, S.; Moriya, M.; Shimada, H.; Hirose, F.; Hirano-Iwata, A.; Mizugaki, Y. Equivalent Circuit Model Modified for Free-Standing Bilayer Lipid Membranes beyond 1 TΩ. Jpn. J. Appl. Phys. 2019, 58, SDDK02. [Google Scholar] [CrossRef]
- Varró, A.; Tomek, J.; Nagy, N.; Virág, L.; Passini, E.; Rodriguez, B.; Baczkó, I. Cardiac Transmembrane Ion Channels and Action Potentials: Cellular Physiology and Arrhythmogenic Behavior. Physiol. Rev. 2021, 101, 1083–1176. [Google Scholar] [CrossRef] [PubMed]
- Bagal, S.K.; Brown, A.D.; Cox, P.J.; Omoto, K.; Owen, R.M.; Pryde, D.C.; Sidders, B.; Skerratt, S.E.; Stevens, E.B.; Storer, R.I.; et al. Ion Channels as Therapeutic Targets: A Drug Discovery Perspective. J. Med. Chem. 2013, 56, 593–624. [Google Scholar] [CrossRef] [PubMed]
- Hite, R.K.; MacKinnon, R. Structural Titration of Slo2.2, a Na + -Dependent K + Channel. Cell 2017, 168, 390–399.e11. [Google Scholar] [CrossRef]
- Yelshanskaya, M.V.; Patel, D.S.; Kottke, C.M.; Kurnikova, M.G.; Sobolevsky, A.I. Opening of Glutamate Receptor Channel to Subconductance Levels. Nature 2022, 605, 172–178. [Google Scholar] [CrossRef]
- Komiya, M.; Kato, M.; Tadaki, D.; Ma, T.; Yamamoto, H.; Tero, R.; Tozawa, Y.; Niwano, M.; Hirano-Iwata, A. Advances in Artificial Cell Membrane Systems as a Platform for Reconstituting Ion Channels. Chem. Rec. 2020, 20, 730–742. [Google Scholar] [CrossRef]
- Gregorio, N.E.; Levine, M.Z.; Oza, J.P. A User’s Guide to Cell-Free Protein Synthesis. Methods Protoc. 2019, 2, 24. [Google Scholar] [CrossRef]
- Silverman, A.D.; Karim, A.S.; Jewett, M.C. Cell-Free Gene Expression: An Expanded Repertoire of Applications. Nat. Rev. Genet. 2020, 21, 151–170. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Sato, M.; Komiya, M.; Feng, X.; Tadaki, D.; Hirano-Iwata, A. Advances in Artificial Bilayer Lipid Membranes as a Novel Biosensing Platform: From Drug-Screening to Self-Assembled Devices. Chem. Lett. 2021, 50, 418–425. [Google Scholar] [CrossRef]
- Ma, T.; Feng, X.; Ohori, T.; Miyata, R.; Tadaki, D.; Yamaura, D.; Deguchi, T.; Komiya, M.; Kanomata, K.; Hirose, F.; et al. Modulation of Photoinduced Transmembrane Currents in a Fullerene-Doped Freestanding Lipid Bilayer by a Lateral Bias. ACS Omega 2019, 4, 18299–18303. [Google Scholar] [CrossRef]
- Ma, T.; Sato, M.; Komiya, M.; Kanomata, K.; Watanabe, T.; Feng, X.; Miyata, R.; Tadaki, D.; Hirose, F.; Tozawa, Y.; et al. Lateral Voltage as a New Input for Artificial Lipid Bilayer Systems. Faraday Discuss. 2022, 233, 244–256. [Google Scholar] [CrossRef]
- Tadaki, D.; Yamaura, D.; Araki, S.; Yoshida, M.; Arata, K.; Ohori, T.; Ishibashi, K.; Kato, M.; Ma, T.; Miyata, R.; et al. Mechanically Stable Solvent-Free Lipid Bilayers in Nano- and Micro-Tapered Apertures for Reconstitution of Cell-Free Synthesized HERG Channels. Sci. Rep. 2017, 7, 17736. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, G.; McMahon, S.B. The Physiological Function of Different Voltage-Gated Sodium Channels in Pain. Nat. Rev. Neurosci. 2021, 22, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Imbrici, P.; Liantonio, A.; Camerino, G.M.; De Bellis, M.; Camerino, C.; Mele, A.; Giustino, A.; Pierno, S.; De Luca, A.; Tricarico, D.; et al. Therapeutic Approaches to Genetic Ion Channelopathies and Perspectives in Drug Discovery. Front. Pharmacol. 2016, 7, 121. [Google Scholar] [CrossRef] [PubMed]
- Koivisto, A.-P.; Belvisi, M.G.; Gaudet, R.; Szallasi, A. Advances in TRP Channel Drug Discovery: From Target Validation to Clinical Studies. Nat. Rev. Drug Discov. 2022, 21, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Wisedchaisri, G.; Gamal El-Din, T.M. Druggability of Voltage-Gated Sodium Channels—Exploring Old and New Drug Receptor Sites. Front. Pharmacol. 2022, 13, 858348. [Google Scholar] [CrossRef]
- Vandenberg, J.I.; Perry, M.D.; Perrin, M.J.; Mann, S.A.; Ke, Y.; Hill, A.P. HERG K + Channels: Structure, Function, and Clinical Significance. Physiol. Rev. 2012, 92, 1393–1478. [Google Scholar] [CrossRef]
- Sanguinetti, M.C.; Tristani-Firouzi, M. HERG Potassium Channels and Cardiac Arrhythmia. Nature 2006, 440, 463–469. [Google Scholar] [CrossRef]
- Itoh, H.; Crotti, L.; Aiba, T.; Spazzolini, C.; Denjoy, I.; Fressart, V.; Hayashi, K.; Nakajima, T.; Ohno, S.; Makiyama, T.; et al. The Genetics Underlying Acquired Long QT Syndrome: Impact for Genetic Screening. Eur. Heart J. 2016, 37, 1456–1464. [Google Scholar] [CrossRef]
- Mueller, P.; Rudin, D.O.; Tien, H.T.; Wescott, W.C. Reconstitution of Cell Membrane Structure in Vitro and Its Transformation into an Excitable System. Nature 1962, 194, 979–980. [Google Scholar] [CrossRef]
- Oshima, A.; Hirano-Iwata, A.; Mozumi, H.; Ishinari, Y.; Kimura, Y.; Niwano, M. Reconstitution of Human Ether-a-Go-Go -Related Gene Channels in Microfabricated Silicon Chips. Anal. Chem. 2013, 85, 4363–4369. [Google Scholar] [CrossRef] [PubMed]
- Montal, M.; Mueller, P. Formation of Bimolecular Membranes from Lipid Monolayers and a Study of Their Electrical Properties. Proc. Natl. Acad. Sci. USA 1972, 69, 3561–3566. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.A.; Bright, L.K.; Aspinwall, C.A. Photolithographic Fabrication of Microapertures with Well-Defined, Three-Dimensional Geometries for Suspended Lipid Membrane Studies. Anal. Chem. 2013, 85, 9078–9086. [Google Scholar] [CrossRef]
- Ahmed, T.; Bafna, J.A.; Hemmler, R.; Gall, K.; Wagner, R.; Winterhalter, M.; Vellekoop, M.J.; van den Driesche, S. Silicon Nitride-Based Micro-Apertures Coated with Parylene for the Investigation of Pore Proteins Fused in Free-Standing Lipid Bilayers. Membranes 2022, 12, 309. [Google Scholar] [CrossRef]
- Yamada, T.; Sugiura, H.; Mimura, H.; Kamiya, K.; Osaki, T.; Takeuchi, S. Highly Sensitive VOC Detectors Using Insect Olfactory Receptors Reconstituted into Lipid Bilayers. Sci. Adv. 2021, 7, eabd2013. [Google Scholar] [CrossRef] [PubMed]
- Mita, K.; Sumikama, T.; Iwamoto, M.; Matsuki, Y.; Shigemi, K.; Oiki, S. Conductance Selectivity of Na + across the K + Channel via Na + Trapped in a Tortuous Trajectory. Proc. Natl. Acad. Sci. USA 2021, 118, e2017168118. [Google Scholar] [CrossRef]
- Funakoshi, K.; Suzuki, H.; Takeuchi, S. Lipid Bilayer Formation by Contacting Monolayers in a Microfluidic Device for Membrane Protein Analysis. Anal. Chem. 2006, 78, 8169–8174. [Google Scholar] [CrossRef]
- Hirano-Iwata, A.; Taira, T.; Oshima, A.; Kimura, Y.; Niwano, M. Improved Stability of Free-Standing Lipid Bilayers Based on Nanoporous Alumina Films. Appl. Phys. Lett. 2010, 96, 213706. [Google Scholar] [CrossRef]
- Tsemperouli, M.; Sugihara, K. Characterization of Di-4-ANEPPS with Nano-Black Lipid Membranes. Nanoscale 2018, 10, 1090–1098. [Google Scholar] [CrossRef]
- Syeda, R.; Holden, M.A.; Hwang, W.L.; Bayley, H. Screening Blockers Against a Potassium Channel with a Droplet Interface Bilayer Array. J. Am. Chem. Soc. 2008, 130, 15543–15548. [Google Scholar] [CrossRef]
- Tadaki, D.; Yamaura, D.; Arata, K.; Ohori, T.; Ma, T.; Yamamoto, H.; Niwano, M.; Hirano-Iwata, A. Micro- and Nanofabrication Methods for Ion Channel Reconstitution in Bilayer Lipid Membranes. Jpn. J. Appl. Phys. 2018, 57, 03EA01. [Google Scholar] [CrossRef]
- Hirano-Iwata, A.; Aoto, K.; Oshima, A.; Taira, T.; Yamaguchi, R.; Kimura, Y.; Niwano, M. Free-Standing Lipid Bilayers in Silicon Chips−Membrane Stabilization Based on Microfabricated Apertures with a Nanometer-Scale Smoothness. Langmuir 2010, 26, 1949–1952. [Google Scholar] [CrossRef] [PubMed]
- Yamaura, D.; Tadaki, D.; Araki, S.; Yoshida, M.; Arata, K.; Ohori, T.; Ishibashi, K.; Kato, M.; Ma, T.; Miyata, R.; et al. Amphiphobic Septa Enhance the Mechanical Stability of Free-Standing Bilayer Lipid Membranes. Langmuir 2018, 34, 5615–5622. [Google Scholar] [CrossRef] [PubMed]
- Hirano-Iwata, A.; Ishinari, Y.; Yoshida, M.; Araki, S.; Tadaki, D.; Miyata, R.; Ishibashi, K.; Yamamoto, H.; Kimura, Y.; Niwano, M. Reconstitution of Human Ion Channels into Solvent-Free Lipid Bilayers Enhanced by Centrifugal Forces. Biophys. J. 2016, 110, 2207–2215. [Google Scholar] [CrossRef]
- Zou, A.; Curran, M.E.; Keating, M.T.; Sanguinetti, M.C. Single HERG Delayed Rectifier K+ Channels Expressed in Xenopus Oocytes. Am. J. Physiol. Heart Circ. Physiol. 1997, 272, H1309–H1314. [Google Scholar] [CrossRef]
- Hirano-Iwata, A.; Oshima, A.; Mozumi, H.; Kimura, Y.; Niwano, M. Stable Lipid Bilayers Based on Micro- and Nano-Fabrication as a Platform for Recording Ion-Channel Activities. Anal. Sci. 2012, 28, 1049–1057. [Google Scholar] [CrossRef]
- Kamiya, K.; Osaki, T.; Nakao, K.; Kawano, R.; Fujii, S.; Misawa, N.; Hayakawa, M.; Takeuchi, S. Electrophysiological Measurement of Ion Channels on Plasma/Organelle Membranes Using an on-Chip Lipid Bilayer System. Sci. Rep. 2018, 8, 17498. [Google Scholar] [CrossRef]
- Portonovo, S.A.; Salazar, C.S.; Schmidt, J.J. HERG Drug Response Measured in Droplet Bilayers. Biomed. Microdevices 2013, 15, 255–259. [Google Scholar] [CrossRef]
- Guo, L.; Guthrie, H. Automated Electrophysiology in the Preclinical Evaluation of Drugs for Potential QT Prolongation. J. Pharmacol. Toxicol. Methods 2005, 52, 123–135. [Google Scholar] [CrossRef]
- Mohammad, S.; Zhou, Z.; Gong, Q.; January, C.T. Blockage of the HERG Human Cardiac K+ Channel by the Gastrointestinal Prokinetic Agent Cisapride. Am. J. Physiol. Heart Circ. Physiol. 1997, 273, H2534–H2538. [Google Scholar] [CrossRef]
- Perrin, M.J.; Kuchel, P.W.; Campbell, T.J.; Vandenberg, J.I. Drug Binding to the Inactivated State Is Necessary but Not Sufficient for High-Affinity Binding to Human Ether-à-Go-Go -Related Gene Channels. Mol. Pharmacol. 2008, 74, 1443–1452. [Google Scholar] [CrossRef] [PubMed]
- Toga, T.; Kohmura, Y.; Kawatsu, R. The 5-HT4 Agonists Cisapride, Mosapride, and CJ-033466, a Novel Potent Compound, Exhibit Different Human Ether-a-Go-Go-Related Gene (HERG)-Blocking Activities. J. Pharmacol. Sci. 2007, 105, 207–210. [Google Scholar] [CrossRef] [PubMed]
- de Planque, M.R.R. Lipid Bilayer Platforms for Parallel Ion Channel Recordings. Jpn. J. Appl. Phys. 2022, 61, SC0804. [Google Scholar] [CrossRef]
- Miyata, R.; Tadaki, D.; Yamaura, D.; Araki, S.; Sato, M.; Komiya, M.; Ma, T.; Yamamoto, H.; Niwano, M.; Hirano-Iwata, A. Parallel Recordings of Transmembrane HERG Channel Currents Based on Solvent-Free Lipid Bilayer Microarray. Micromachines 2021, 12, 98. [Google Scholar] [CrossRef] [PubMed]
- Hodgkin, A.L.; Huxley, A.F.; Katz, B. Measurement of Current-voltage Relations in the Membrane of the Giant Axon of Loligo. J. Physiol. 1952, 116, 424–448. [Google Scholar] [CrossRef]
- Hodgkin, A.L.; Huxley, A.F. The Dual Effect of Membrane Potential on Sodium Conductance in the Giant Axon of Loligo. J. Physiol. 1952, 116, 497–506. [Google Scholar] [CrossRef]
- Kodandaramaiah, S.B.; Franzesi, G.T.; Chow, B.Y.; Boyden, E.S.; Forest, C.R. Automated Whole-Cell Patch-Clamp Electrophysiology of Neurons in Vivo. Nat. Methods 2012, 9, 585–587. [Google Scholar] [CrossRef]
- Dixon, C.; Sah, P.; Lynch, J.W.; Keramidas, A. GABAA Receptor α and γ Subunits Shape Synaptic Currents via Different Mechanisms. J. Biol. Chem. 2014, 289, 5399–5411. [Google Scholar] [CrossRef]
- Muñoz, B.; Mariqueo, T.; Murath, P.; Peters, C.; Yevenes, G.E.; Moraga-Cid, G.; Peoples, R.W.; Aguayo, L.G. Modulatory Actions of the Glycine Receptor β Subunit on the Positive Allosteric Modulation of Ethanol in A2 Containing Receptors. Front. Mol. Neurosci. 2021, 14, 763868. [Google Scholar] [CrossRef]
- Renauld, S.; Cortes, S.; Bersch, B.; Henry, X.; De Waard, M.; Schaack, B. Functional Reconstitution of Cell-Free Synthesized Purified Kv Channels. Biochim. Biophys. Acta BBA Biomembr. 2017, 1859, 2373–2380. [Google Scholar] [CrossRef]
- Friddin, M.S.; Smithers, N.P.; Beaugrand, M.; Marcotte, I.; Williamson, P.T.F.; Morgan, H.; de Planque, M.R.R. Single-Channel Electrophysiology of Cell-Free Expressed Ion Channels by Direct Incorporation in Lipid Bilayers. Analyst 2013, 138, 7294. [Google Scholar] [CrossRef] [PubMed]
- Ando, M.; Akiyama, M.; Okuno, D.; Hirano, M.; Ide, T.; Sawada, S.; Sasaki, Y.; Akiyoshi, K. Liposome Chaperon in Cell-Free Membrane Protein Synthesis: One-Step Preparation of KcsA-Integrated Liposomes and Electrophysiological Analysis by the Planar Bilayer Method. Biomater. Sci. 2016, 4, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Winterstein, L.-M.; Kukovetz, K.; Rauh, O.; Turman, D.L.; Braun, C.; Moroni, A.; Schroeder, I.; Thiel, G. Reconstitution and Functional Characterization of Ion Channels from Nanodiscs in Lipid Bilayers. J. Gen. Physiol. 2018, 150, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.; Kriebel, J.K.; Tosteson, M.T.; Whitesides, G.M. Microfabricated Teflon Membranes for Low-Noise Recordings of Ion Channels in Planar Lipid Bilayers. Biophys. J. 2003, 85, 2684–2695. [Google Scholar] [CrossRef]
- Mantri, S.; Sapra, K.T.; Cheley, S.; Sharp, T.H.; Bayley, H. An Engineered Dimeric Protein Pore That Spans Adjacent Lipid Bilayers. Nat. Commun. 2013, 4, 1725. [Google Scholar] [CrossRef]
- Nakajima, T.; Kaneko, Y.; Dharmawan, T.; Kurabayashi, M. Role of the Voltage Sensor Module in Na v Domain IV on Fast Inactivation in Sodium Channelopathies: The Implication of Closed-State Inactivation. Channels 2019, 13, 331–343. [Google Scholar] [CrossRef]
- Amin, A.S.; Asghari-Roodsari, A.; Tan, H.L. Cardiac Sodium Channelopathies. Pflüg. Arch. Eur. J. Physiol. 2010, 460, 223–237. [Google Scholar] [CrossRef]
- Catterall, W.A. Voltage-Gated Sodium Channels at 60: Structure, Function and Pathophysiology: Voltage-Gated Sodium Channels. J. Physiol. 2012, 590, 2577–2589. [Google Scholar] [CrossRef]
- Beyder, A.; Rae, J.L.; Bernard, C.; Strege, P.R.; Sachs, F.; Farrugia, G. Mechanosensitivity of Nav 1.5, a Voltage-Sensitive Sodium Channel: Nav1.5 Mechanosensitivity. J. Physiol. 2010, 588, 4969–4985. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kageyama, H.; Ma, T.; Sato, M.; Komiya, M.; Tadaki, D.; Hirano-Iwata, A. New Aspects of Bilayer Lipid Membranes for the Analysis of Ion Channel Functions. Membranes 2022, 12, 863. https://doi.org/10.3390/membranes12090863
Kageyama H, Ma T, Sato M, Komiya M, Tadaki D, Hirano-Iwata A. New Aspects of Bilayer Lipid Membranes for the Analysis of Ion Channel Functions. Membranes. 2022; 12(9):863. https://doi.org/10.3390/membranes12090863
Chicago/Turabian StyleKageyama, Hironori, Teng Ma, Madoka Sato, Maki Komiya, Daisuke Tadaki, and Ayumi Hirano-Iwata. 2022. "New Aspects of Bilayer Lipid Membranes for the Analysis of Ion Channel Functions" Membranes 12, no. 9: 863. https://doi.org/10.3390/membranes12090863
APA StyleKageyama, H., Ma, T., Sato, M., Komiya, M., Tadaki, D., & Hirano-Iwata, A. (2022). New Aspects of Bilayer Lipid Membranes for the Analysis of Ion Channel Functions. Membranes, 12(9), 863. https://doi.org/10.3390/membranes12090863