
Genomic Insights into Bacterial Resistance to Proline-Rich Antimicrobial Peptide Bac7

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antimicrobial Agents
2.2. Bacterial Strains
2.3. Antimicrobial Assay
2.4. Resistance Induction Experiments
2.5. Whole-Genome Sequencing
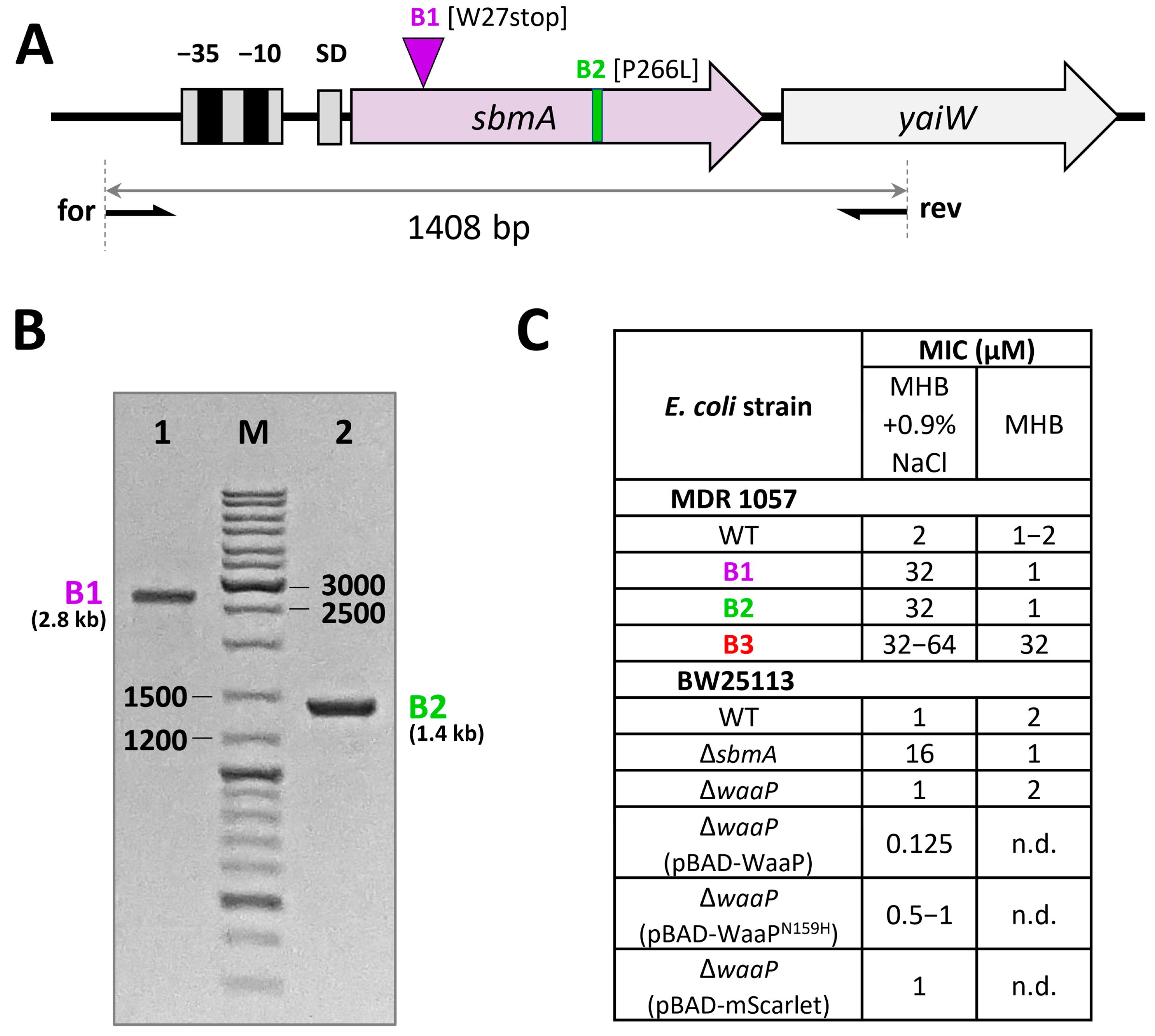
2.6. Analysis of the sbmA Gene
2.7. Construction of Complementation Plasmids
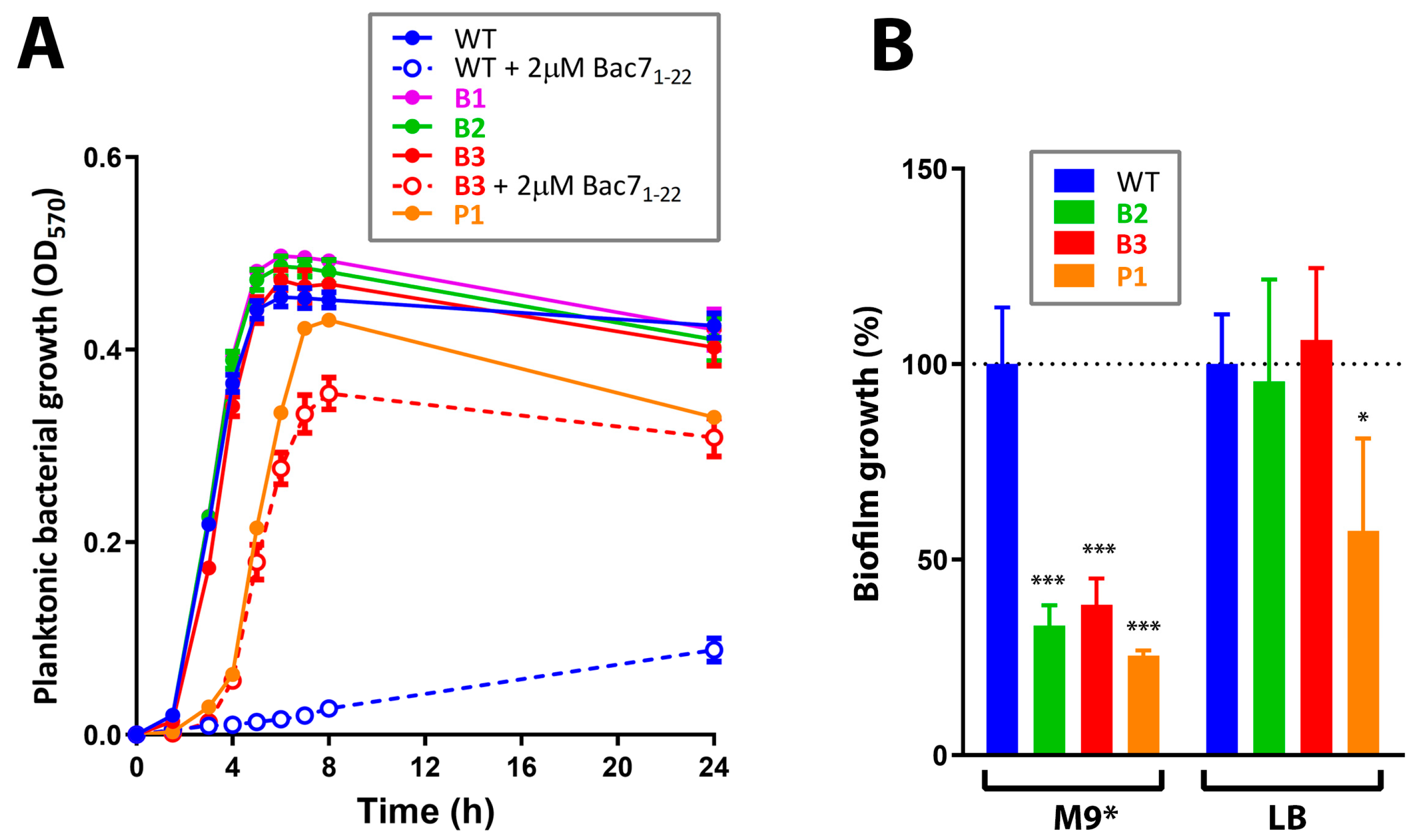
2.8. Growth Rate Determination
2.9. Biofilm Assay
3. Results and Discussion
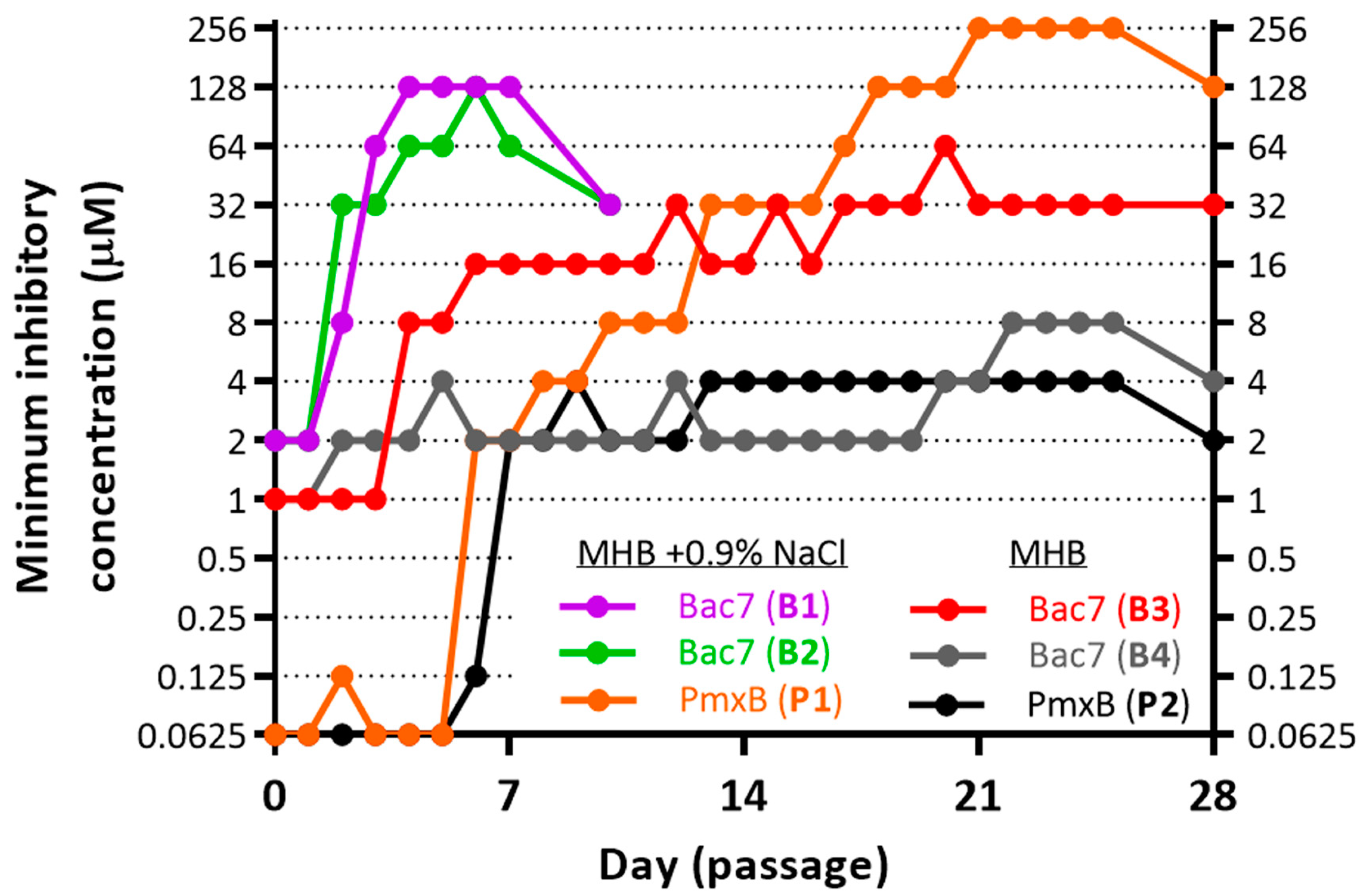
3.1. Induction of Bacterial Resistance
3.2. Resistance to Bac71-22 in the Salt-Containing Medium Is Mediated by Inactivation of the SbmA Transporter
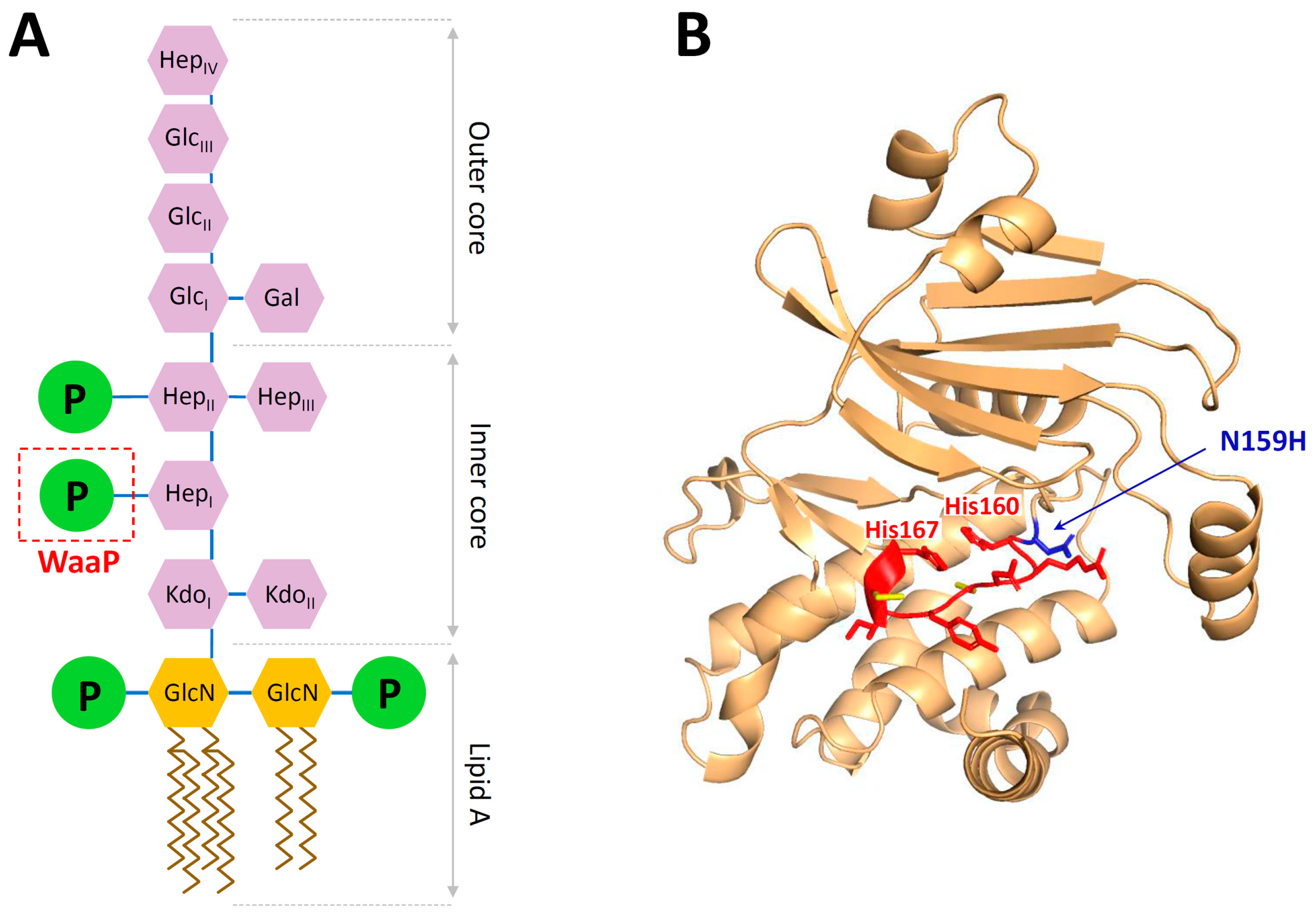
3.3. The Single Mutation in the WaaP Kinase Contributes to E. coli Resistance to Bac71-22
3.4. The Super-Resistance of E. coli to Polymyxin B Is Mediated by Seven Genome Mutations
3.5. Screening of Cross-Resistance Effects
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Graf, M.; Mardirossian, M.; Nguyen, F.; Seefeldt, A.C.; Guichard, G.; Scocchi, M.; Innis, C.A.; Wilson, D.N. Proline-Rich Antimicrobial Peptides Targeting Protein Synthesis. Nat. Prod. Rep. 2017, 34, 702–711. [Google Scholar] [CrossRef]
- Ghilarov, D.; Inaba-Inoue, S.; Stepien, P.; Qu, F.; Michalczyk, E.; Pakosz, Z.; Nomura, N.; Ogasawara, S.; Walker, G.C.; Rebuffat, S.; et al. Molecular Mechanism of SbmA, a Promiscuous Transporter Exploited by Antimicrobial Peptides. Sci. Adv. 2021, 7, eabj5363. [Google Scholar] [CrossRef] [PubMed]
- Panteleev, P.V.; Bolosov, I.A.; Kalashnikov, A.À.; Kokryakov, V.N.; Shamova, O.V.; Emelianova, A.A.; Balandin, S.V.; Ovchinnikova, T.V. Combined Antibacterial Effects of Goat Cathelicidins with Different Mechanisms of Action. Front. Microbiol. 2018, 9, 2983. [Google Scholar] [CrossRef] [PubMed]
- Gennaro, R.; Skerlavaj, B.; Romeo, D. Purification, Composition, and Activity of Two Bactenecins, Antibacterial Peptides of Bovine Neutrophils. Infect. Immun. 1989, 57, 3142–3146. [Google Scholar] [CrossRef] [PubMed]
- Seefeldt, A.C.; Graf, M.; Pérébaskine, N.; Nguyen, F.; Arenz, S.; Mardirossian, M.; Scocchi, M.; Wilson, D.N.; Innis, C.A. Structure of the Mammalian Antimicrobial Peptide Bac7(1–16) Bound within the Exit Tunnel of a Bacterial Ribosome. Nucleic Acids Res. 2016, 44, 2429–2438. [Google Scholar] [CrossRef]
- Gagnon, M.G.; Roy, R.N.; Lomakin, I.B.; Florin, T.; Mankin, A.S.; Steitz, T.A. Structures of Proline-Rich Peptides Bound to the Ribosome Reveal a Common Mechanism of Protein Synthesis Inhibition. Nucleic Acids Res. 2016, 44, 2439–2450. [Google Scholar] [CrossRef]
- Mardirossian, M.; Sola, R.; Beckert, B.; Valencic, E.; Collis, D.W.P.; Borišek, J.; Armas, F.; Di Stasi, A.; Buchmann, J.; Syroegin, E.A.; et al. Peptide Inhibitors of Bacterial Protein Synthesis with Broad Spectrum and SbmA-Independent Bactericidal Activity against Clinical Pathogens. J. Med. Chem. 2020, 63, 9590–9602. [Google Scholar] [CrossRef]
- Benincasa, M.; Scocchi, M.; Podda, E.; Skerlavaj, B.; Dolzani, L.; Gennaro, R. Antimicrobial Activity of Bac7 Fragments against Drug-Resistant Clinical Isolates. Peptides 2004, 25, 2055–2061. [Google Scholar] [CrossRef]
- Veldhuizen, E.J.A.; Schneider, V.A.F.; Agustiandari, H.; van Dijk, A.; Tjeerdsma-van Bokhoven, J.L.M.; Bikker, F.J.; Haagsman, H.P. Antimicrobial and Immunomodulatory Activities of PR-39 Derived Peptides. PLoS ONE 2014, 9, e95939. [Google Scholar] [CrossRef]
- Shamova, O.V.; Orlov, D.S.; Zharkova, M.S.; Balandin, S.V.; Yamschikova, E.V.; Knappe, D.; Hoffmann, R.; Kokryakov, V.N.; Ovchinnikova, T.V. Minibactenecins ChBac7.Nα and ChBac7. Nβ—Antimicrobial Peptides from Leukocytes of the Goat Capra Hircus. Acta Nat. 2016, 8, 136–146. [Google Scholar] [CrossRef]
- Koch, P.; Schmitt, S.; Heynisch, A.; Gumpinger, A.; Wüthrich, I.; Gysin, M.; Shcherbakov, D.; Hobbie, S.N.; Panke, S.; Held, M. Optimization of the Antimicrobial Peptide Bac7 by Deep Mutational Scanning. BMC Biol. 2022, 20, 114. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.K.; Kumari, T.; Tandon, A.; Mohd, S.; Afshan, T.; Kathuria, M.; Shukla, P.K.; Mitra, K.; Ghosh, J.K. Selective Phenylalanine to Proline Substitution for Improved Antimicrobial and Anticancer Activities of Peptides Designed on Phenylalanine Heptad Repeat. Acta Biomater. 2017, 57, 170–186. [Google Scholar] [CrossRef] [PubMed]
- Holfeld, L.; Herth, N.; Singer, D.; Hoffmann, R.; Knappe, D. Immunogenicity and Pharmacokinetics of Short, Proline-Rich Antimicrobial Peptides. Future Med. Chem. 2015, 7, 1581–1596. [Google Scholar] [CrossRef] [PubMed]
- Lamb, H.M.; Wiseman, L.R. Pexiganan Acetate. Drugs 1998, 56, 1047–1052; discussion 1053–1054. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Safronova, V.N.; Kruglikov, R.N.; Bolosov, I.A.; Bogdanov, I.V.; Ovchinnikova, T.V. A Novel Proline-Rich Cathelicidin from the Alpaca Vicugna pacos with Potency to Combat Antibiotic-Resistant Bacteria: Mechanism of Action and the Functional Role of the C-Terminal Region. Membranes 2022, 12, 515. [Google Scholar] [CrossRef]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 In-frame, Single-gene Knockout Mutants: The Keio Collection. Mol. Syst. Biol. 2006, 2. [Google Scholar] [CrossRef]
- de Sena Brandine, G.; Smith, A.D. Falco: High-Speed FastQC Emulation for Quality Control of Sequencing Data. F1000Research 2021, 8, 1874. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Koboldt, D.C.; Zhang, Q.; Larson, D.E.; Shen, D.; McLellan, M.D.; Lin, L.; Miller, C.A.; Mardis, E.R.; Ding, L.; Wilson, R.K. VarScan 2: Somatic Mutation and Copy Number Alteration Discovery in Cancer by Exome Sequencing. Genome Res. 2012, 22, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Komp Lindgren, P.; Karlsson, A.; Hughes, D. Mutation Rate and Evolution of Fluoroquinolone Resistance in Escherichia coli Isolates from Patients with Urinary Tract Infections. Antimicrob. Agents Chemother. 2003, 47, 3222–3232. [Google Scholar] [CrossRef]
- Pitt, M.E.; Cao, M.D.; Butler, M.S.; Ramu, S.; Ganesamoorthy, D.; Blaskovich, M.A.T.; Coin, L.J.M.; Cooper, M.A. Octapeptin C4 and Polymyxin Resistance Occur via Distinct Pathways in an Epidemic XDR Klebsiella pneumoniae ST258 Isolate. J. Antimicrob. Chemother. 2019, 74, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Bolosov, I.A.; Panteleev, P.V.; Balandin, S.V.; Shamova, O.V.; Ovchinnikova, T.V. Structural and Functional Characteristics of the Proline-Rich Antimicrobial Peptide Minibactenecin from Leukocytes of Domestic Goat Capra Hircus. Bull. Exp. Biol. Med. 2023, 174, 440–445. [Google Scholar] [CrossRef]
- Slotboom, D.J.; Ettema, T.W.; Nijland, M.; Thangaratnarajah, C. Bacterial Multi-solute Transporters. FEBS Lett. 2020, 594, 3898–3907. [Google Scholar] [CrossRef]
- Krizsan, A.; Knappe, D.; Hoffmann, R. Influence of the YjiL-MdtM Gene Cluster on the Antibacterial Activity of Proline-Rich Antimicrobial Peptides Overcoming Escherichia coli Resistance Induced by the Missing SbmA Transporter System. Antimicrob. Agents Chemother. 2015, 59, 5992–5998. [Google Scholar] [CrossRef]
- Imai, Y.; Hauk, G.; Quigley, J.; Liang, L.; Son, S.; Ghiglieri, M.; Gates, M.F.; Morrissette, M.; Shahsavari, N.; Niles, S.; et al. Evybactin Is a DNA Gyrase Inhibitor That Selectively Kills Mycobacterium tuberculosis. Nat. Chem. Biol. 2022, 18, 1236–1244. [Google Scholar] [CrossRef]
- Metelev, M.; Osterman, I.A.; Ghilarov, D.; Khabibullina, N.F.; Yakimov, A.; Shabalin, K.; Utkina, I.; Travin, D.Y.; Komarova, E.S.; Serebryakova, M.; et al. Klebsazolicin Inhibits 70S Ribosome by Obstructing the Peptide Exit Tunnel. Nat. Chem. Biol. 2017, 13, 1129–1136. [Google Scholar] [CrossRef]
- Glazebrook, J.; Ichige, A.; Walker, G.C. A Rhizobium Meliloti Homolog of the Escherichia coli Peptide-Antibiotic Transport Protein SbmA Is Essential for Bacteroid Development. Genes Dev. 1993, 7, 1485–1497. [Google Scholar] [CrossRef] [PubMed]
- Pränting, M.; Negrea, A.; Rhen, M.; Andersson, D.I. Mechanism and Fitness Costs of PR-39 Resistance in Salmonella enterica Serovar Typhimurium LT2. Antimicrob. Agents Chemother. 2008, 52, 2734–2741. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Krizsan, A.; Volke, D.; Knappe, D.; Hoffmann, R. Identification of New Resistance Mechanisms in Escherichia coli against Apidaecin 1b Using Quantitative Gel- and LC–MS-Based Proteomics. J. Proteome Res. 2016, 15, 2607–2617. [Google Scholar] [CrossRef] [PubMed]
- Knappe, D.; Kabankov, N.; Herth, N.; Hoffmann, R. Insect-Derived Short Proline-Rich and Murine Cathelicidin-Related Antimicrobial Peptides Act Synergistically on Gram-Negative Bacteria in Vitro. Future Med. Chem. 2016, 8, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, K.; Venclovas, C.; Ioerger, T.R.; Sacchettini, J.C.; McKinney, J.D.; Mizrahi, V.; Warner, D.F. A Vitamin B₁₂ Transporter in Mycobacterium tuberculosis. Open Biol. 2013, 3, 120175. [Google Scholar] [CrossRef]
- Rempel, S.; Gati, C.; Nijland, M.; Thangaratnarajah, C.; Karyolaimos, A.; de Gier, J.W.; Guskov, A.; Slotboom, D.J. A Mycobacterial ABC Transporter Mediates the Uptake of Hydrophilic Compounds. Nature 2020, 580, 409–412. [Google Scholar] [CrossRef]
- Domenech, P.; Kobayashi, H.; LeVier, K.; Walker, G.C.; Barry, C.E. BacA, an ABC Transporter Involved in Maintenance of Chronic Murine Infections with Mycobacterium tuberculosis. J. Bacteriol. 2009, 191, 477–485. [Google Scholar] [CrossRef]
- Arnold, M.F.F.; Haag, A.F.; Capewell, S.; Boshoff, H.I.; James, E.K.; McDonald, R.; Mair, I.; Mitchell, A.M.; Kerscher, B.; Mitchell, T.J.; et al. Partial Complementation of Sinorhizobium meliloti BacA Mutant Phenotypes by the Mycobacterium tuberculosis BacA Protein. J. Bacteriol. 2013, 195, 389–398. [Google Scholar] [CrossRef]
- Linde, C.M.; Hoffner, S.E.; Refai, E.; Andersson, M. In Vitro Activity of PR-39, a Proline-Arginine-Rich Peptide, against Susceptible and Multi-Drug-Resistant Mycobacterium tuberculosis. J. Antimicrob. Chemother. 2001, 47, 575–580. [Google Scholar] [CrossRef]
- Yethon, J.A.; Whitfield, C. Purification and Characterization of WaaP from Escherichia coli, a Lipopolysaccharide Kinase Essential for Outer Membrane Stability. J. Biol. Chem. 2001, 276, 5498–5504. [Google Scholar] [CrossRef]
- Spohn, R.; Daruka, L.; Lázár, V.; Martins, A.; Vidovics, F.; Grézal, G.; Méhi, O.; Kintses, B.; Számel, M.; Jangir, P.K.; et al. Integrated Evolutionary Analysis Reveals Antimicrobial Peptides with Limited Resistance. Nat. Commun. 2019, 10, 4538. [Google Scholar] [CrossRef] [PubMed]
- Kreamer, N.N.K.; Chopra, R.; Caughlan, R.E.; Fabbro, D.; Fang, E.; Gee, P.; Hunt, I.; Li, M.; Leon, B.C.; Muller, L.; et al. Acylated-Acyl Carrier Protein Stabilizes the Pseudomonas aeruginosa WaaP Lipopolysaccharide Heptose Kinase. Sci. Rep. 2018, 8, 14124. [Google Scholar] [CrossRef] [PubMed]
- Yethon, J.A.; Heinrichs, D.E.; Monteiro, M.A.; Perry, M.B.; Whitfield, C. Involvement of WaaY, WaaQ, and WaaP in the Modification of Escherichia coliLipopolysaccharide and Their Role in the Formation of a Stable Outer Membrane. J. Biol. Chem. 1998, 273, 26310–26316. [Google Scholar] [CrossRef] [PubMed]
- Yethon, J.A.; Gunn, J.S.; Ernst, R.K.; Miller, S.I.; Laroche, L.; Malo, D.; Whitfield, C. Salmonella enterica Serovar Typhimurium WaaP Mutants Show Increased Susceptibility to Polymyxin and Loss of Virulence In Vivo. Infect. Immun. 2000, 68, 4485–4491. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, J.; Ren, G.; Li, Y.; Wang, X. Influence of Core Oligosaccharide of Lipopolysaccharide to Outer Membrane Behavior of Escherichia coli. Mar. Drugs 2015, 13, 3325–3339. [Google Scholar] [CrossRef] [PubMed]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making Protein Folding Accessible to All. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]
- Rubin, E.J.; Herrera, C.M.; Crofts, A.A.; Trent, M.S. PmrD Is Required for Modifications to Escherichia coli Endotoxin That Promote Antimicrobial Resistance. Antimicrob. Agents Chemother. 2015, 59, 2051–2061. [Google Scholar] [CrossRef]
- Ghiselli, R.; Giacometti, A.; Cirioni, O.; Circo, R.; Mocchegiani, F.; Skerlavaj, B.; D’Amato, G.; Scalise, G.; Zanetti, M.; Saba, V. Neutralization of Endotoxin In Vitro and In Vivo by BAC7(1-35), a Proline-Rich Antibacterial Peptide. Shock 2003, 19, 577–581. [Google Scholar] [CrossRef]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Chin, C.-Y.; Gregg, K.A.; Napier, B.A.; Ernst, R.K.; Weiss, D.S. A PmrB-Regulated Deacetylase Required for Lipid A Modification and Polymyxin Resistance in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2015, 59, 7911–7914. [Google Scholar] [CrossRef]
- Knopp, M.; Babina, A.M.; Gudmundsdóttir, J.S.; Douglass, M.V.; Trent, M.S.; Andersson, D.I. A Novel Type of Colistin Resistance Genes Selected from Random Sequence Space. PLoS Genet. 2021, 17, e1009227. [Google Scholar] [CrossRef] [PubMed]
- Quesada, A.; Porrero, M.C.; Téllez, S.; Palomo, G.; García, M.; Domínguez, L. Polymorphism of Genes Encoding PmrAB in Colistin-Resistant Strains of Escherichia coli and Salmonella enterica Isolated from Poultry and Swine. J. Antimicrob. Chemother. 2015, 70, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Negrea, A.; Rhen, M.; Andersson, D.I. Genetic Analysis of Colistin Resistance in Salmonella enterica Serovar Typhimurium. Antimicrob. Agents Chemother. 2009, 53, 2298–2305. [Google Scholar] [CrossRef] [PubMed]
- Luther, A.; Urfer, M.; Zahn, M.; Müller, M.; Wang, S.-Y.; Mondal, M.; Vitale, A.; Hartmann, J.-B.; Sharpe, T.; Monte, F.L.; et al. Chimeric Peptidomimetic Antibiotics against Gram-Negative Bacteria. Nature 2019, 576, 452–458. [Google Scholar] [CrossRef]
- Vetterli, S.U.; Zerbe, K.; Müller, M.; Urfer, M.; Mondal, M.; Wang, S.-Y.; Moehle, K.; Zerbe, O.; Vitale, A.; Pessi, G.; et al. Thanatin Targets the Intermembrane Protein Complex Required for Lipopolysaccharide Transport in Escherichia coli. Sci. Adv. 2018, 4, eaau2634. [Google Scholar] [CrossRef]
- Elliott, A.G.; Huang, J.X.; Neve, S.; Zuegg, J.; Edwards, I.A.; Cain, A.K.; Boinett, C.J.; Barquist, L.; Lundberg, C.V.; Steen, J.; et al. An Amphipathic Peptide with Antibiotic Activity against Multidrug-Resistant Gram-Negative Bacteria. Nat. Commun. 2020, 11, 3184. [Google Scholar] [CrossRef]
- Krenev, I.A.; Panteleev, P.V.; Umnyakova, E.S.; Gorbunov, N.P.; Kostevich, V.A.; Balandin, S.V.; Ovchinnikova, T.V.; Aleshina, G.M.; Berlov, M.N. In Vitro Modulation of Complement Activation by Therapeutically Prospective Analogues of the Marine Polychaeta Arenicin Peptides. Mar. Drugs 2022, 20, 612. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Ovchinnikova, T.V. Improved Strategy for Recombinant Production and Purification of Antimicrobial Peptide Tachyplesin I and Its Analogs with High Cell Selectivity. Biotechnol. Appl. Biochem. 2017, 64, 35–42. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Bolosov, I.A.; Khokhlova, V.A.; Dhanda, G.; Balandin, S.V.; Haldar, J.; Ovchinnikova, T.V. Analysis of Antibacterial Action of Mammalian Host-Defense Cathelicidins and Induction of Resistance to Them in MβL-Producing Pseudomonas aeruginosa. Bull. Exp. Biol. Med. 2022, 172, 447–452. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Balandin, S.V.; Ovchinnikova, T.V. Effect of Arenicins and Other β-Hairpin Antimicrobial Peptides on Pseudomonas aeruginosa PAO1 Biofilms. Pharm. Chem. J. 2017, 50, 715–720. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Selection Conditions | Mutation | Type | Gene | Gene Product |
|---|---|---|---|---|---|
| B1 | MHB + 0.9%NaCl Bac71-22 (7 passages) | W27stop (TGG→TGA) | insertion (IC3-like element, 1.4 kb) | sbmA | cytoplasmic membrane transporter |
| B2 | MHB + 0.9%NaCl Bac71-22 (7 passages) | P266L(CCG→CTG) | single nucleotide polymorphism | sbmA | cytoplasmic membrane transporter |
| B3 | MHB Bac71-22 (25 passages) | N159H (AAC→CAC) | single nucleotide polymorphism | waaP | heptose specific lipopolysaccharide (LPS) core kinase |
| 21st position (G→T) | single nucleotide polymorphism | menE/ pmrD | intergenic 109 bp region | ||
| P1 | MHB + 0.9%NaCl PmxB (25 passages) | I130S (ATT→AGT) | single nucleotide polymorphism | bamA | outer membrane protein assembly complex |
| R371H (CGT→CAT) | single nucleotide polymorphism | sppA | cytoplasmic membrane signal peptide peptidase | ||
| N118H (AAC→CAC) | single nucleotide polymorphism | spoT | bifunctional (p)ppGpp synthase/hydrolase | ||
| Q304stop (CAG→TAG) | in-frame stop codon | rpoS | RNA polymerase sigma factor σS | ||
| Q830stop (CAG→TAG) | in-frame stop codon | secA | protein translocation ATPase | ||
| V161G (GTG→GGG) | single nucleotide polymorphism | pmrB (basS) | cytoplasmic membrane sensor histidine kinase PmrB | ||
| S305R (AGT→CGT) | single nucleotide polymorphism |
| Antibacterial Agent | Minimum Inhibitory Concentration, µM | |||||
|---|---|---|---|---|---|---|
| WT | B2 | B3 | P1 | |||
| Antimicrobial peptides | proline-rich | Bac71-22 | 2 | >32 | >32 | 16 * |
| mini-ChBac7.5Nα | 8 | >32 | >32 | >32 | ||
| PR-391-22 | 2 | >32 | >32 | 16 | ||
| VicBac | 1 | 2 | 2 | 4 | ||
| β-hairpin | AA139 ** | 0.25 | 0.25 | 1 | 0.5 | |
| Tachyplesin-1 | 0.063 | 0.063 | 0.063 | 0.063 | ||
| Protegrin-1 | 0.25 | 0.25 | 0.25 | 0.25 | ||
| Thanatin | 1 | 1 | >32 | >32 | ||
| α-helical | Pexiganan ** | 1 | 1 | 2 | 8 | |
| LL-37 | 4 | 2 | 2 | 1 | ||
| ChMAP-28 | 0.063 | 0.063 | 0.063 | 0.063 | ||
| Melittin | 8 | 8 | 4 | 4 | ||
| Polymyxin B | 0.125 | 0.125 | 8 | >128 | ||
| Conventional antibiotics | Ampicillin | 256 | 256 | 256 | 256 | |
| Ceftriaxone | 128 | 128 | 128 | 128 | ||
| Ciprofloxacin | 256 | 128 | 256 | 128 | ||
| Rifampicin | 64 | 128 | 64 | 64 | ||
| Tetracycline | 8 | 16 | 8 | 8 | ||
| Gentamicin | 2 | 2 | 2 | 4 | ||
| Chloramphenicol | 8 | 16 | 8 | 16 | ||
| Spectinomycin | 32 | 32 | 32 | 64 | ||
| Clindamycin | 128 | 256 | 128 | 256 | ||
| Roxithromycin | 256 | 256 | 256 | >256 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panteleev, P.V.; Safronova, V.N.; Kruglikov, R.N.; Bolosov, I.A.; Ovchinnikova, T.V. Genomic Insights into Bacterial Resistance to Proline-Rich Antimicrobial Peptide Bac7. Membranes 2023, 13, 438. https://doi.org/10.3390/membranes13040438
Panteleev PV, Safronova VN, Kruglikov RN, Bolosov IA, Ovchinnikova TV. Genomic Insights into Bacterial Resistance to Proline-Rich Antimicrobial Peptide Bac7. Membranes. 2023; 13(4):438. https://doi.org/10.3390/membranes13040438
Chicago/Turabian StylePanteleev, Pavel V., Victoria N. Safronova, Roman N. Kruglikov, Ilia A. Bolosov, and Tatiana V. Ovchinnikova. 2023. "Genomic Insights into Bacterial Resistance to Proline-Rich Antimicrobial Peptide Bac7" Membranes 13, no. 4: 438. https://doi.org/10.3390/membranes13040438
APA StylePanteleev, P. V., Safronova, V. N., Kruglikov, R. N., Bolosov, I. A., & Ovchinnikova, T. V. (2023). Genomic Insights into Bacterial Resistance to Proline-Rich Antimicrobial Peptide Bac7. Membranes, 13(4), 438. https://doi.org/10.3390/membranes13040438