

Critical Role of Molecular Packing in Lo Phase Membrane Solubilization


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Large Unilamellar Vesicle Preparation
2.3. Light Scattering Measurements
2.4. Fluorescence Measurements
2.4.1. Lipid Aggregate Structure at the Molecular Level
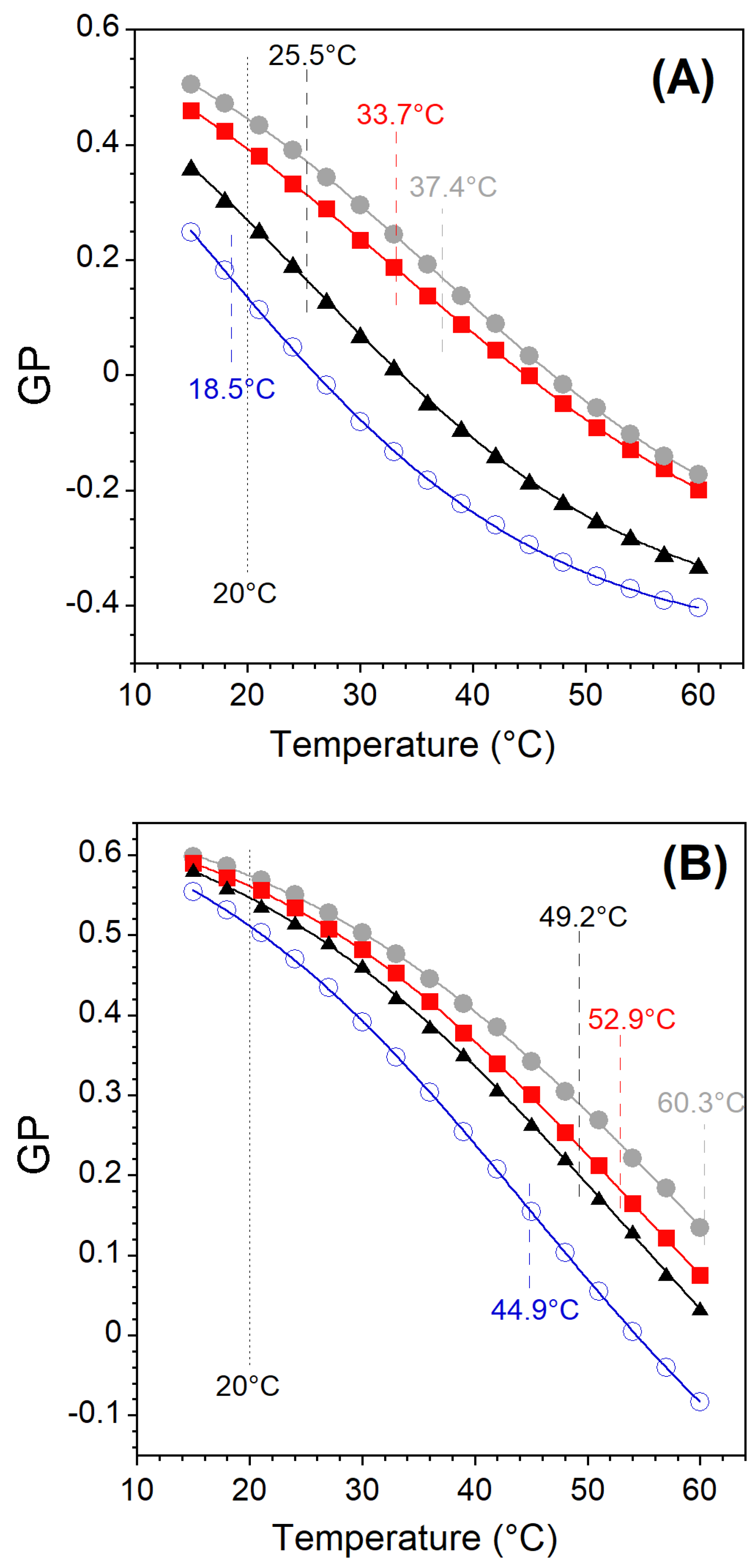
2.4.2. Membrane Phase State
3. Results
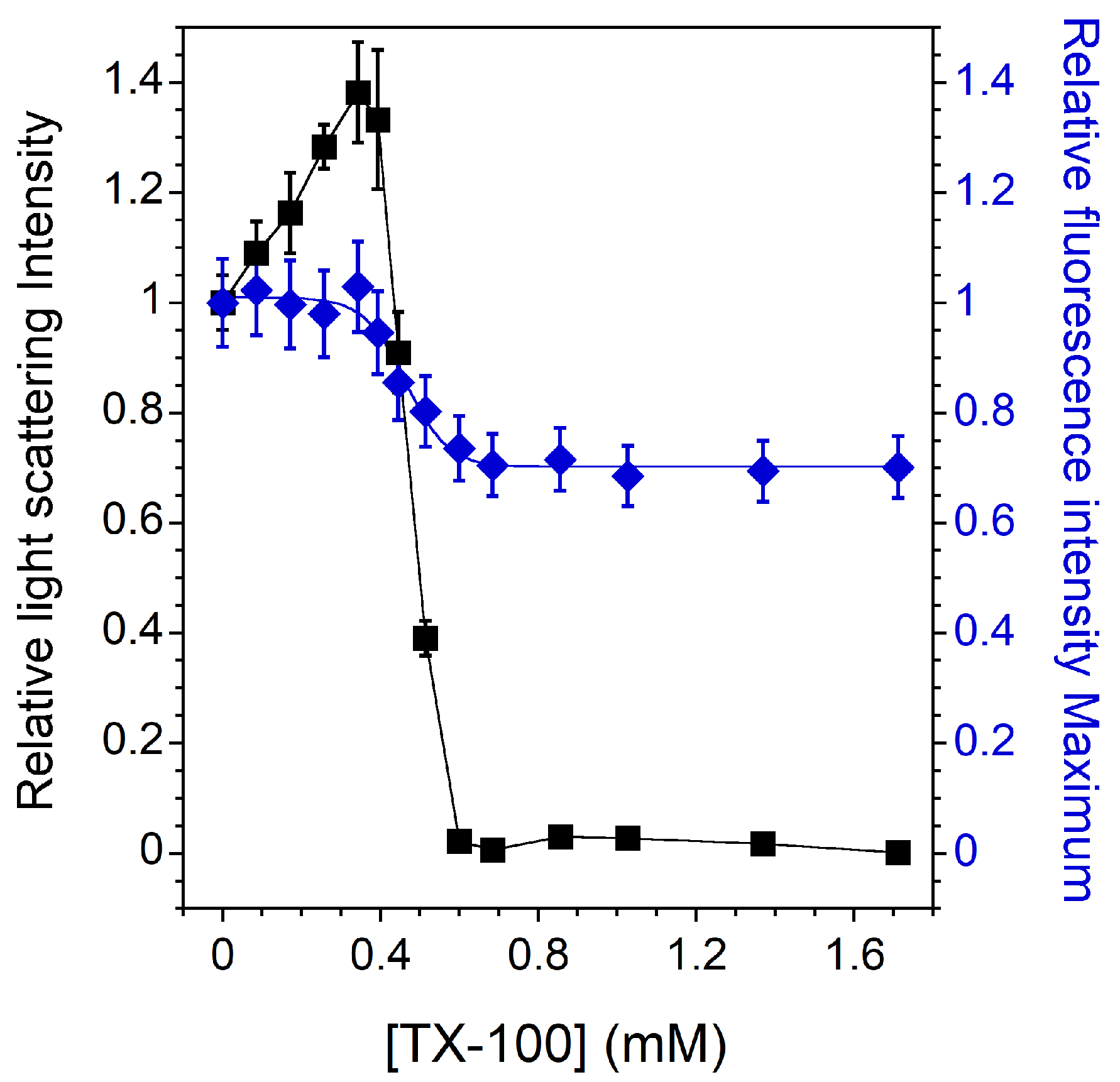
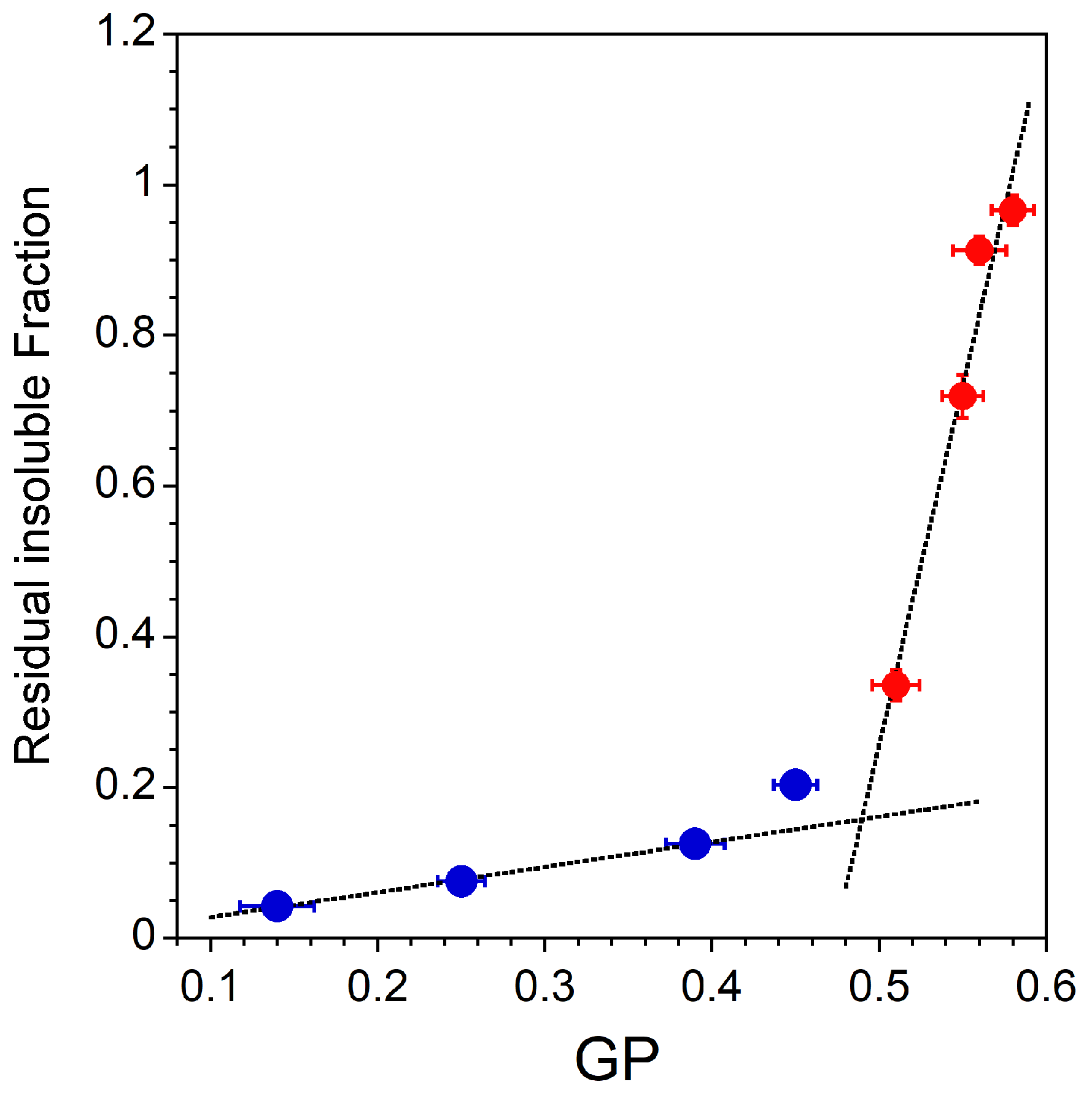
3.1. Vesicle-to-Micelle Transition
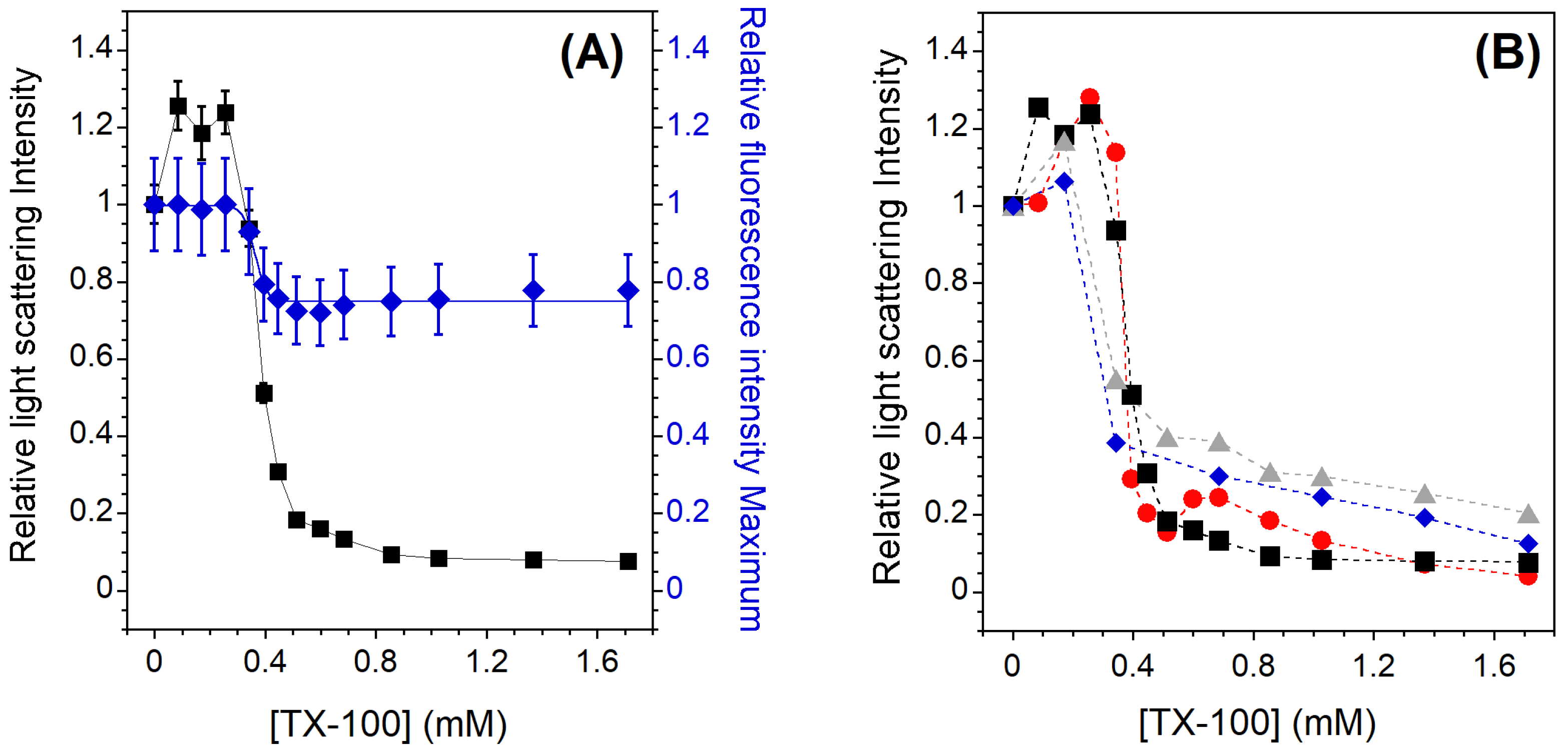
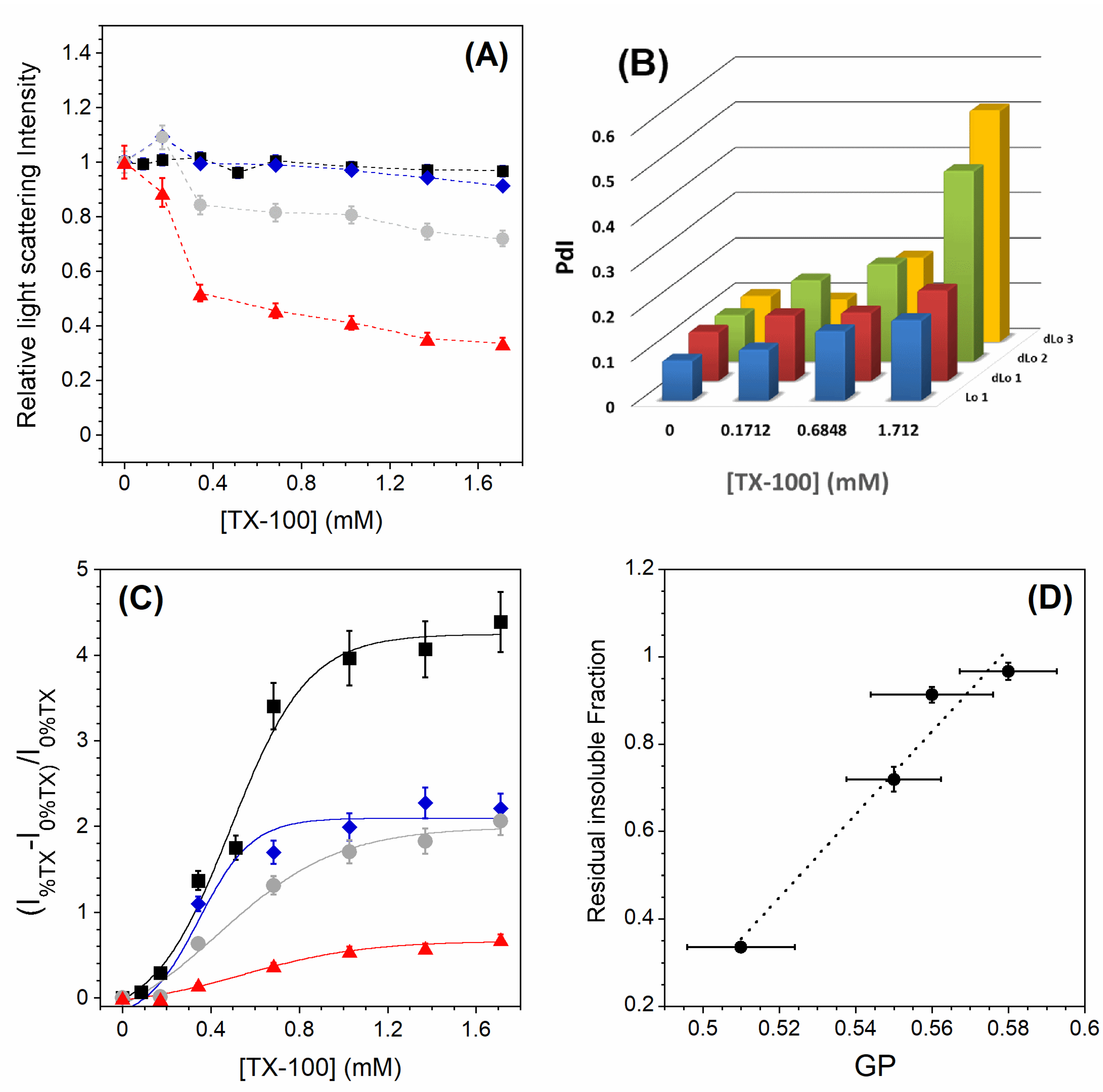
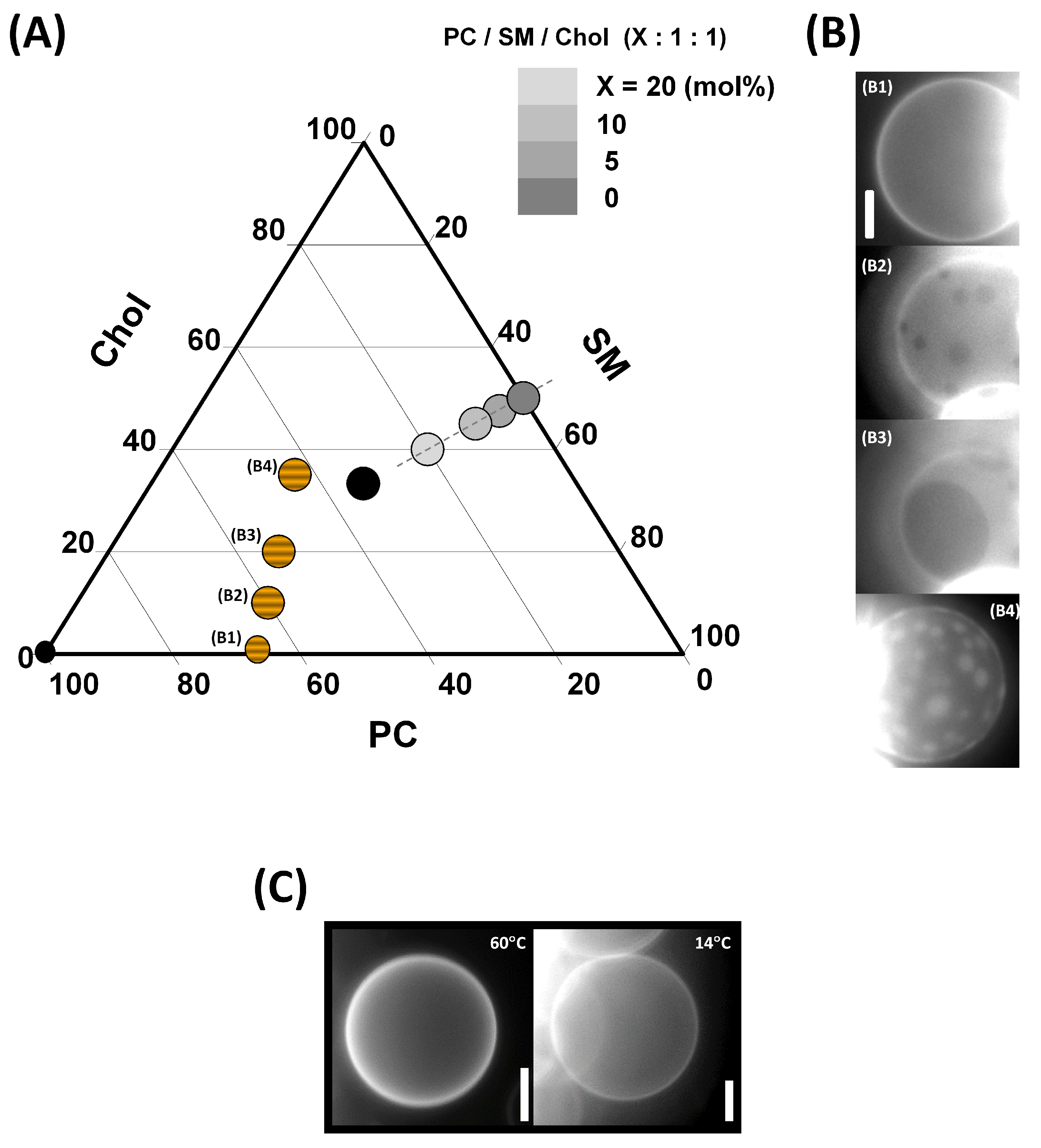
3.2. Solubilization of Membranes Exhibiting Lo/Ld Phase Coexistence
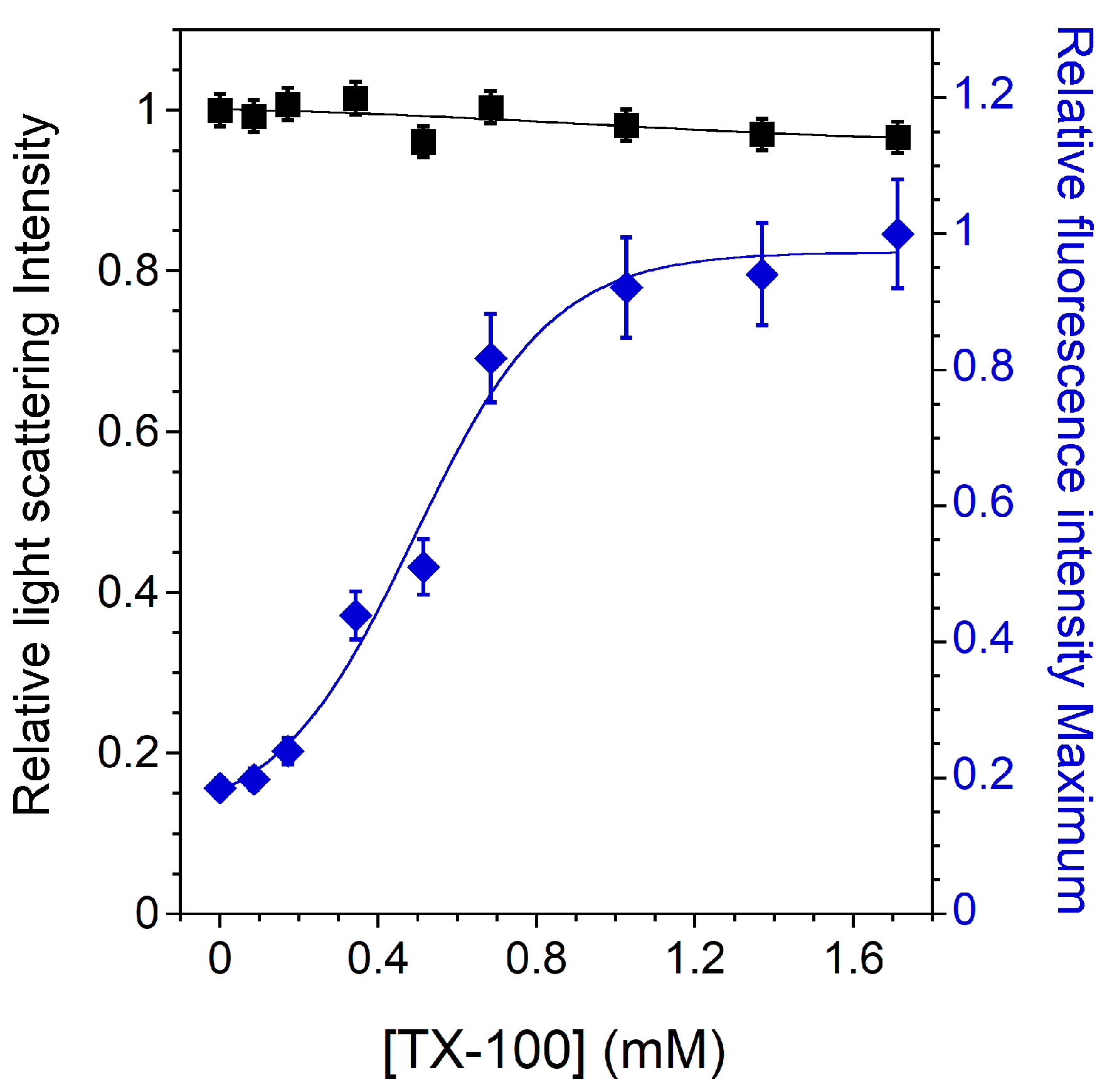
3.3. Lo and PC-Doped Lo Phase Membrane Solubilization
4. Discussion
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Chol | Cholesterol |
| DRM | Detergent-resistant membrane |
| TX-100 | Triton X-100 |
| Lo | Liquid-ordered |
| Ld | Liquid-disordered |
| GUV | Giant unilamellar vesicle |
| SM | Egg yolk sphingomyelin |
| LUV | Large unilamellar vesicle |
| GP | Generalized polarization |
| GPMV | Giant plasma membrane vesicle |
| PC | Egg yolk L-α-phosphatidylcholine |
| PdI | Polydispersity index |
| SLS | Static light scattering |
| RIF | Residual insoluble fraction |
| DLS | Dynamic light scattering |
Appendix A
Appendix A.1

Appendix A.2

Appendix A.3

References
- Lindner, R.; Naim, H.Y. Domains in biological membranes. Exp. Cell Res. 2009, 315, 2871–2878. [Google Scholar] [CrossRef]
- Kusumi, A.; Suzuki, K.G.; Kasai, R.S.; Ritchie, K.; Fujiwara, T.K. Hierarchical mesoscale domain organization of the plasma membrane. Trends Biochem. Sci. 2011, 36, 604–615. [Google Scholar] [CrossRef]
- Veatch, S.L.; Rogers, N.; Decker, A.; Shelby, S.A. The plasma membrane as an adaptable fluid mosaic. Biochim. Biophys. Acta Biomembr. 2023, 1865, 184114. [Google Scholar] [CrossRef]
- Nicolson, G.L. The Fluid—Mosaic Model of Membrane Structure: Still relevant to understanding the structure, function and dynamics of biological membranes after more than 40 years. Biochim. Biophys. Acta Biomembr. 2014, 1838, 1451–1466. [Google Scholar] [CrossRef] [Green Version]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef]
- Brown, D.A.; London, E. Structure and Function of Sphingolipid- and Cholesterol-rich Membrane Rafts. J. Biol. Chem. 2000, 275, 17221–17224. [Google Scholar] [CrossRef] [Green Version]
- Pralle, A.; Keller, P.; Florin, E.L.; Simons, K.; Hörber, J. Sphingolipid–Cholesterol Rafts Diffuse as Small Entities in the Plasma Membrane of Mammalian Cells. J. Cell Biol. 2000, 2000. 148, 997–1008. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, K.; Mouritsen, O.G.; Anderson, R.G.W. Lipid rafts: At a crossroad between cell biology and physics. Nat. Cell Biol. 2007, 9, 7–14. [Google Scholar] [CrossRef]
- Cambi, A.; Lidke, D.S. Nanoscale Membrane Organization: Where Biochemistry Meets Advanced Microscopy. ACS Chem. Biol. 2012, 7, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Owen, D.M.; Magenau, A.; Williamson, D.; Gaus, K. The lipid raft hypothesis revisited—New insights on raft composition and function from super-resolution fluorescence microscopy. BioEssays 2012, 34, 739–747. [Google Scholar] [CrossRef]
- Levental, I.; Levental, K.R.; Heberle, F.A. Lipid Rafts: Controversies Resolved, Mysteries Remain. Trends Cell Biol. 2020, 30, 341–353. [Google Scholar] [CrossRef]
- Sezgin, E.; Levental, I.; Mayor, S.; Eggeling, C. The mystery of membrane organization: Composition, regulation and roles of lipid rafts. Nat. Rev. Mol. Cell Biol. 2017, 18, 361–374. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.A.; Rose, J.K. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell 1992, 68, 533–544. [Google Scholar] [CrossRef]
- Schroeder, R.; London, E.; Brown, D. Interactions between saturated acyl chains confer detergent resistance on lipids and glycosylphosphatidylinositol (GPI)-anchored proteins: GPI-anchored proteins in liposomes and cells show similar behavior. Proc. Natl. Acad. Sci. USA 1994, 91, 12130–12134. [Google Scholar] [CrossRef]
- Sot, J.; Collado, M.I.; Arrondo, J.L.R.; Alonso, A.; Goñi, F.M. Triton X-100-Resistant Bilayers: Effect of Lipid Composition and Relevance to the Raft Phenomenon. Langmuir 2002, 18, 2828–2835. [Google Scholar] [CrossRef]
- Lichtenberg, D.; Goñi, F.M.; Heerklotz, H. Detergent-resistant membranes should not be identified with membrane rafts. Trends Biochem. Sci. 2005, 30, 430–436. [Google Scholar] [CrossRef]
- Aureli, M.; Grassi, S.; Sonnino, S.; Prinetti, A. Isolation and Analysis of Detergent-Resistant Membrane Fractions. In Lipid Signaling Protocols; Waugh, M.G., Ed.; Springer: New York, NY, USA, 2016; pp. 107–131. [Google Scholar] [CrossRef]
- London, E.; Brown, D.A. Insolubility of lipids in Triton X-100: Physical origin and relationship to sphingolipid/cholesterol membrane domains (rafts). Biochim. Biophys. Acta Biomembr. 2000, 1508, 182–195. [Google Scholar] [CrossRef] [Green Version]
- McMullen, T.P.; Lewis, R.N.; McElhaney, R.N. Cholesterol–phospholipid interactions, the liquid-ordered phase and lipid rafts in model and biological membranes. Curr. Opin. Colloid Interface Sci. 2004, 8, 459–468. [Google Scholar] [CrossRef]
- Coste, V.; Breton, M.; Angelova, M.I.; Puff, N. How to extract selectively the lo-phase domains from large unilamellar vesicles with Triton X-100? Colloids Surf. A Physicochem. Eng. Asp. 2006, 282-283, 402–409. [Google Scholar] [CrossRef] [Green Version]
- El Kirat, K.; Morandat, S. Cholesterol modulation of membrane resistance to Triton X-100 explored by atomic force microscopy. Biochim. Biophys. Acta Biomembr. 2007, 1768, 2300–2309. [Google Scholar] [CrossRef] [Green Version]
- Caritá, A.C.; Mattei, B.; Domingues, C.C.; de Paula, E.; Riske, K.A. Effect of Triton X-100 on Raft-Like Lipid Mixtures: Phase Separation and Selective Solubilization. Langmuir 2017, 33, 7312–7321. [Google Scholar] [CrossRef]
- Helenius, A.; Simons, K. Solubilization of membranes by detergents. Biochim. Biophys. Acta Rev. Biomembr. 1975, 415, 29–79. [Google Scholar] [CrossRef] [PubMed]
- De la Maza, A.; Parra, J.L. Vesicle-micelle structural transition of phosphatidylcholine bilayers and Triton X-100. Biochem. J. 1994, 303, 907–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patra, S.K.; Alonso, A.; Goñi, F.M. Detergent solubilisation of phospholipid bilayers in the gel state: The role of polar and hydrophobic forces. Biochim. Biophys. Acta Biomembr. 1998, 1373, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Tamba, Y.; Tanaka, T.; Yahagi, T.; Yamashita, Y.; Yamazaki, M. Stability of giant unilamellar vesicles and large unilamellar vesicles of liquid-ordered phase membranes in the presence of Triton X-100. Biochim. Biophys. Acta Biomembr. 2004, 1667, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattei, B.; França, A.D.C.; Riske, K.A. Solubilization of Binary Lipid Mixtures by the Detergent Triton X-100: The Role of Cholesterol. Langmuir 2015, 31, 378–386. [Google Scholar] [CrossRef]
- Staneva, G.; Seigneuret, M.; Koumanov, K.; Trugnan, G.; Angelova, M.I. Detergents induce raft-like domains budding and fission from giant unilamellar heterogeneous vesicles: A direct microscopy observation. Chem. Phys. Lipids 2005, 136, 55–66. [Google Scholar] [CrossRef]
- Casadei, B.; Domingues, C.; de Paula, E.; Riske, K. Direct Visualization of the Action of Triton X-100 on Giant Vesicles of Erythrocyte Membrane Lipids. Biophys. J. 2014, 106, 2417–2425. [Google Scholar] [CrossRef] [Green Version]
- Heerklotz, H. Triton Promotes Domain Formation in Lipid Raft Mixtures. Biophys. J. 2002, 83, 2693–2701. [Google Scholar] [CrossRef] [Green Version]
- Pathak, P.; London, E. Measurement of Lipid Nanodomain (Raft) Formation and Size in Sphingomyelin/POPC/Cholesterol Vesicles Shows TX-100 and Transmembrane Helices Increase Domain Size by Coalescing Preexisting Nanodomains However, Do Not Induce Domain Formation. Biophys. J. 2011, 101, 2417–2425. [Google Scholar] [CrossRef] [Green Version]
- Muddana, H.; Chiang, H.; Butler, P. Tuning Membrane Phase Separation Using Nonlipid Amphiphiles. Biophys. J. 2012, 102, 489–497. [Google Scholar] [CrossRef] [Green Version]
- Sezgin, E.; Kaiser, H.J.; Baumgart, T.; Schwille, P.; Simons, K.; Levental, I. Elucidating membrane structure and protein behavior using giant plasma membrane vesicles. Nat. Protoc. 2012, 7, 1042–1051. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P.; Hammond, A.; Holowka, D.; Baird, B. Structural determinants for partitioning of lipids and proteins between coexisting fluid phases in giant plasma membrane vesicles. Biochim. Biophys. Acta Biomembr. 2008, 1778, 20–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levental, I.; Byfield, F.; Chowdhury, P.; Gai, F.; Baumgart, T.; Janmey, P. Cholesterol-dependent phase separation in cell-derived giant plasma-membrane vesicles. Biochem. J. 2009, 424, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, H.J.; Lingwood, D.; Levental, I.; Sampaio, J.L.; Kalvodova, L.; Rajendran, L.; Simons, K. Order of lipid phases in model and plasma membranes. Proc. Natl. Acad. Sci. USA 2009, 106, 16645–16650. [Google Scholar] [CrossRef]
- Sezgin, E.; Gutmann, T.; Buhl, T.; Dirkx, R.; Grzybek, M.; Coskun, U.; Solimena, M.; Simons, K.; Levental, I.; Schwille, P. Adaptive Lipid Packing and Bioactivity in Membrane Domains. PLoS ONE 2015, 10, e0123930. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, R.C.; MacDonald, R.I.; Menco, B.P.; Takeshita, K.; Subbarao, N.K.; Rong Hu, L. Small-volume extrusion apparatus for preparation of large, unilamellar vesicles. Biochim. Biophys. Acta Biomembr. 1991, 1061, 297–303. [Google Scholar] [CrossRef]
- Grit, M.; de Smidt, J.H.; Struijke, A.; Crommelin, D.J. Hydrolysis of phosphatidylcholine in aqueous liposome dispersions. Int. J. Pharm. 1989, 50, 1–6. [Google Scholar] [CrossRef]
- Grit, M.; Crommelin, D.J. The effect of aging on the physical stability of liposome dispersions. Chem. Phys. Lipids 1992, 62, 113–122. [Google Scholar] [CrossRef]
- Raghuraman, H.; Shrivastava, S.; Chattopadhyay, A. Monitoring the looping up of acyl chain labeled NBD lipids in membranes as a function of membrane phase state. Biochim. Biophys. Acta Biomembr. 2007, 1768, 1258–1267. [Google Scholar] [CrossRef] [Green Version]
- Coste, V.; Puff, N.; Lockau, D.; Quinn, P.J.; Angelova, M.I. Raft-like domain formation in large unilamellar vesicles probed by the fluorescent phospholipid analogue, C12NBD-PC. Biochim. Biophys. Acta Biomembr. 2006, 1758, 460–467. [Google Scholar] [CrossRef] [Green Version]
- Brown, R.S.; Brennan, J.D.; Krull, U.J. Self-quenching of nitrobenzoxadiazole labeled phospholipids in lipid membranes. J. Chem. Phys. 1994, 100, 6019–6027. [Google Scholar] [CrossRef]
- Viard, M.; Gallay, J.; Vincent, M.; Paternostre, M. Origin of Laurdan Sensitivity to the Vesicle-to-Micelle Transition of Phospholipid-Octylglucoside System: A Time-Resolved Fluorescence Study. Biophys. J. 2001, 80, 347–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puff, N.; Staneva, G.; Angelova, M.I.; Seigneuret, M. Improved Characterization of Raft-Mimicking Phase-Separation Phenomena in Lipid Bilayers Using Laurdan Fluorescence with Log-Normal Multipeak Analysis. Langmuir 2020, 36, 4347–4356. [Google Scholar] [CrossRef] [PubMed]
- Umbsaar, J.; Kerek, E.; Prenner, E.J. Cobalt and nickel affect the fluidity of negatively-charged biomimetic membranes. Chem. Phys. Lipids 2018, 210, 28–37. [Google Scholar] [CrossRef]
- Lichtenberg, D.; Ahyayauch, H.; Goñi, F. The Mechanism of Detergent Solubilization of Lipid Bilayers. Biophys. J. 2013, 105, 289–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lete, M.G.; Monasterio, B.G.; Collado, M.I.; Medina, M.; Sot, J.; Alonso, A.; Goñi, F.M. Fast and slow biomembrane solubilizing detergents: Insights into their mechanism of action. Colloids Surf. Biointerfaces 2019, 183, 110430. [Google Scholar] [CrossRef]
- Koynova, R.; Caffrey, M. Phases and phase transitions of the phosphatidylcholines. Biochim. Biophys. Acta Rev. Biomembr. 1998, 1376, 91–145. [Google Scholar] [CrossRef]
- Lin, S.; Struve, W.S. Time-resolved fluorescence of nitrobenzoxadiazole-aminohexanoic acid: Effect of intermolecular hydrogen-bonding on non-radiative decay. Photochem. Photobiol. 1991, 54, 361–365. [Google Scholar] [CrossRef]
- Mainali, L.; Raguz, M.; Subczynski, W. Phase-Separation and Domain-Formation in Cholesterol-Sphingomyelin Mixture: Pulse-EPR Oxygen Probing. Biophys. J. 2011, 101, 837–846. [Google Scholar] [CrossRef] [Green Version]
- Keyvanloo, A.; Shaghaghi, M.; Zuckermann, M.J.; Thewalt, J.L. The Phase Behavior and Organization of Sphingomyelin/Cholesterol Membranes: A Deuterium NMR Study. Biophys. J. 2018, 114, 1344–1356. [Google Scholar] [CrossRef] [Green Version]
- Mattei, B.; Lira, R.B.; Perez, K.R.; Riske, K.A. Membrane permeabilization induced by Triton X-100: The role of membrane phase state and edge tension. Chem. Phys. Lipids 2017, 202, 28–37. [Google Scholar] [CrossRef]
- Loura, L.M.; Ramalho, J.P. Location and dynamics of acyl chain NBD-labeled phosphatidylcholine (NBD-PC) in DPPC bilayers. A molecular dynamics and time-resolved fluorescence anisotropy study. Biochim. Biophys. Acta Biomembr. 2007, 1768, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Filipe, H.A.L.; Moreno, M.J.; Loura, L.M.S. The Secret Lives of Fluorescent Membrane Probes as Revealed by Molecular Dynamics Simulations. Molecules 2020, 25, 3424. [Google Scholar] [CrossRef]
- Toro, C.; Sanchez, S.; Zanocco, A.; Lemp, E.; Gratton, E.; Gunther, G. Solubilization of lipid bilayers by myristyl sucrose ester: Effect of cholesterol and phospholipid head group size. Chem. Phys. Lipids 2009, 157, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Levental, I.; Grzybek, M.; Simons, K. Raft domains of variable properties and compositions in plasma membrane vesicles. Proc. Natl. Acad. Sci. USA 2011, 108, 11411–11416. [Google Scholar] [CrossRef]
- Manni, M.M.; Cano, A.; Alonso, C.; Goñi, F.M. Lipids that determine detergent resistance of MDCK cell membrane fractions. Chem. Phys. Lipids 2015, 191, 68–74. [Google Scholar] [CrossRef]
- Puff, N.; Watanabe, C.; Seigneuret, M.; Angelova, M.I.; Staneva, G. Lo/Ld phase coexistence modulation induced by GM1. Biochim. Biophys. Acta Biomembr. 2014, 1838, 2105–2114. [Google Scholar] [CrossRef] [Green Version]
- Becerra, N.; de la Nuez, L.; Zanocco, A.; Lemp, E.; Günther, G. Solubilization of dodac small unilamellar vesicles by sucrose esters: A fluorescence study. Colloids Surf. Physicochem. Eng. Asp. 2006, 272, 2–7. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
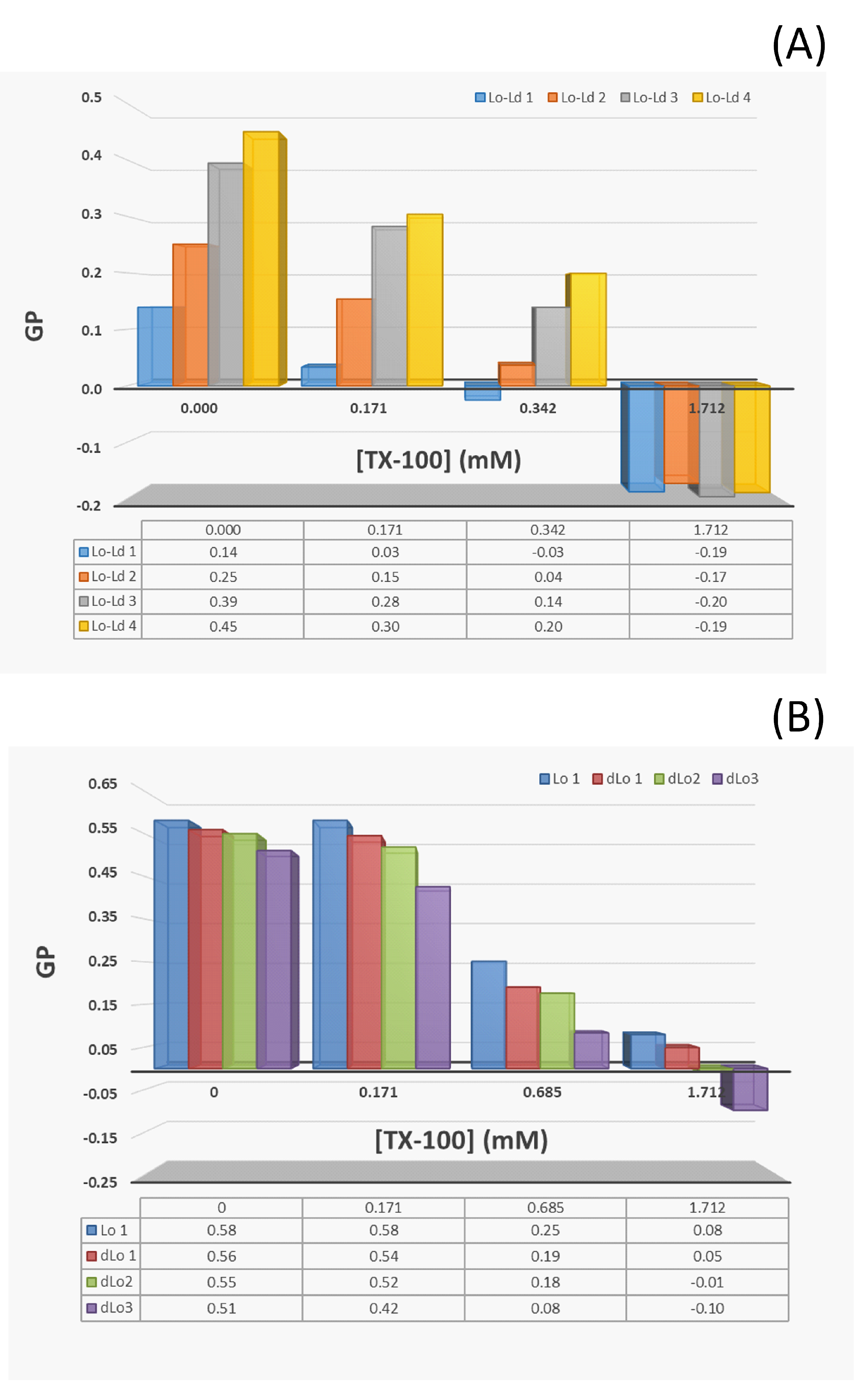
| Sample Name | PC/SM/Chol (mol%) (a) | (b) | (c) | ( °C) (d) | RIF (e) | (c) |
|---|---|---|---|---|---|---|
| Lo-Ld 1 | 2:1:10 | + | 0.14 | 18.5 (f) | + | −0.19 |
| Lo-Ld 2 | 2:1:20 | ++ | 0.25 | 25.5 | + | −0.17 |
| Lo-Ld 3 | 2:1:35 | +++ | 0.39 | 33.7 | ++ | −0.20 |
| Lo-Ld 4 | 1:1:1 | No data | 0.45 | 37.4 | ++ | −0.19 |
| Sample Name | PC/SM/Chol (mol%) (a) | (b) | (a.u.) (c) | ( °C) (d) | RIF (e) | (f) |
|---|---|---|---|---|---|---|
| Lo 1 | 0:1:1 | 0.57 | 76.2 | 60.3 | ++++ | 0.50 |
| dLo 1 | 5:1:1 | 0.56 | 103.3 | 52.9 | ++++ | 0.51 |
| dLo 2 | 10:1:1 | 0.55 | 122.1 | 49.2 | +++ | 0.56 |
| dLo 3 | 20:1:1 | 0.51 | 243.8 | 44.9 | ++ | 0.61 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puff, N. Critical Role of Molecular Packing in Lo Phase Membrane Solubilization. Membranes 2023, 13, 652. https://doi.org/10.3390/membranes13070652
Puff N. Critical Role of Molecular Packing in Lo Phase Membrane Solubilization. Membranes. 2023; 13(7):652. https://doi.org/10.3390/membranes13070652
Chicago/Turabian StylePuff, Nicolas. 2023. "Critical Role of Molecular Packing in Lo Phase Membrane Solubilization" Membranes 13, no. 7: 652. https://doi.org/10.3390/membranes13070652
APA StylePuff, N. (2023). Critical Role of Molecular Packing in Lo Phase Membrane Solubilization. Membranes, 13(7), 652. https://doi.org/10.3390/membranes13070652