



An Untargeted Metabolomics Strategy to Identify Substrates of Known and Orphan E. coli Transporters


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Cultures
2.2. Serum Incubation Experiments
2.3. Preparation for Metabolomics Analysis
2.4. LC-MS(MS) Analysis
2.5. Data Preprocessing and Analysis
3. Results and Discussion
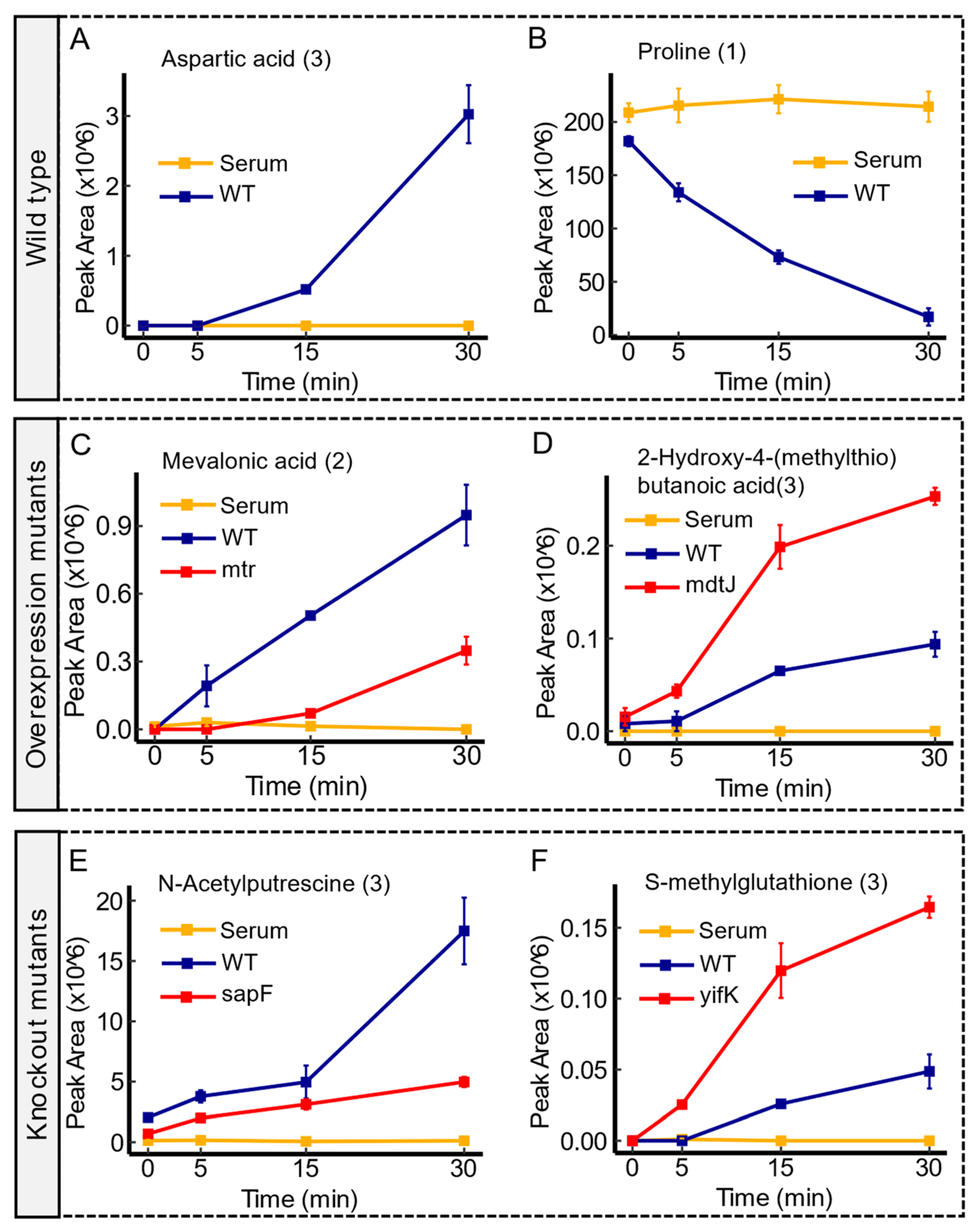
3.1. Metabolite Profiling of Strains and Compound Identification
3.2. The Differential Transport of Metabolites
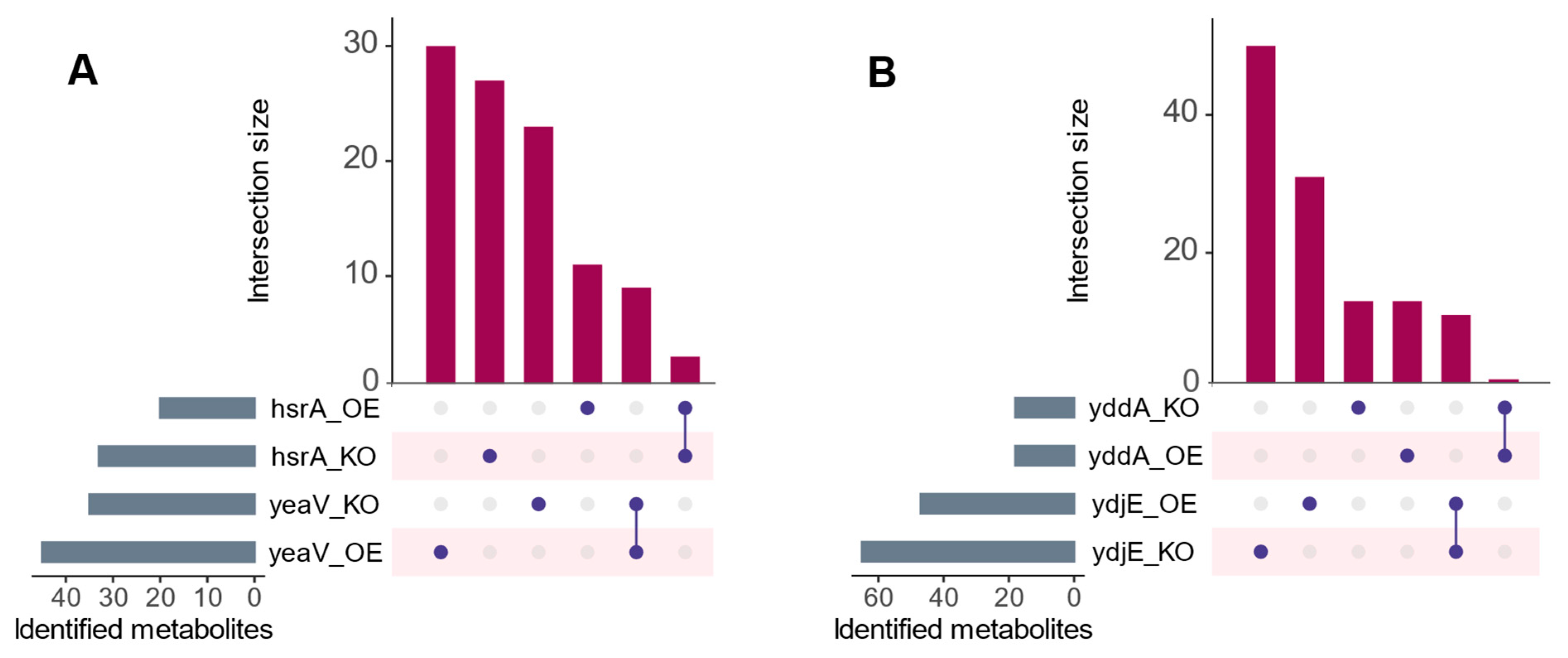
3.3. Probing Candidate Substrates of Putative Transporters
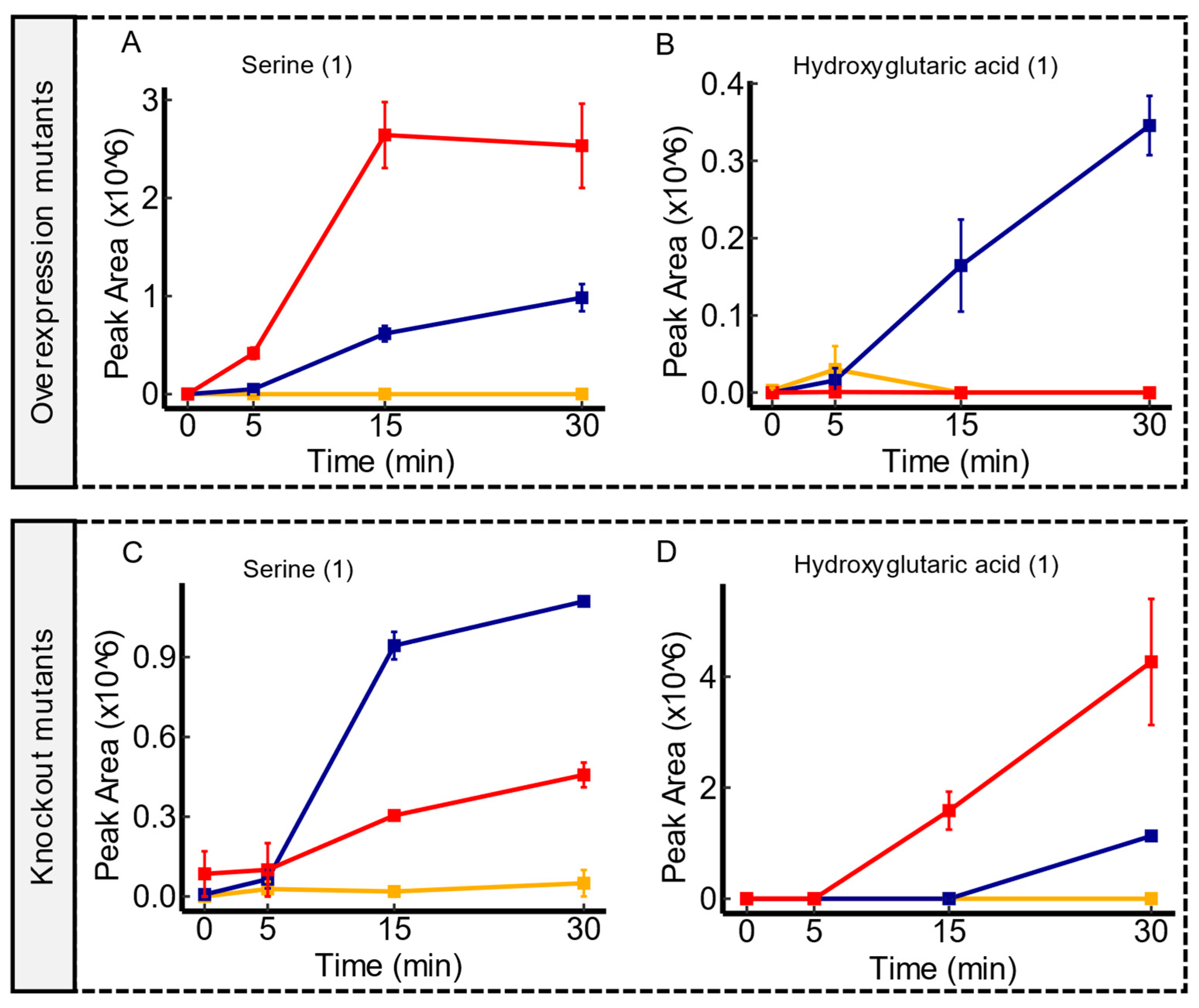
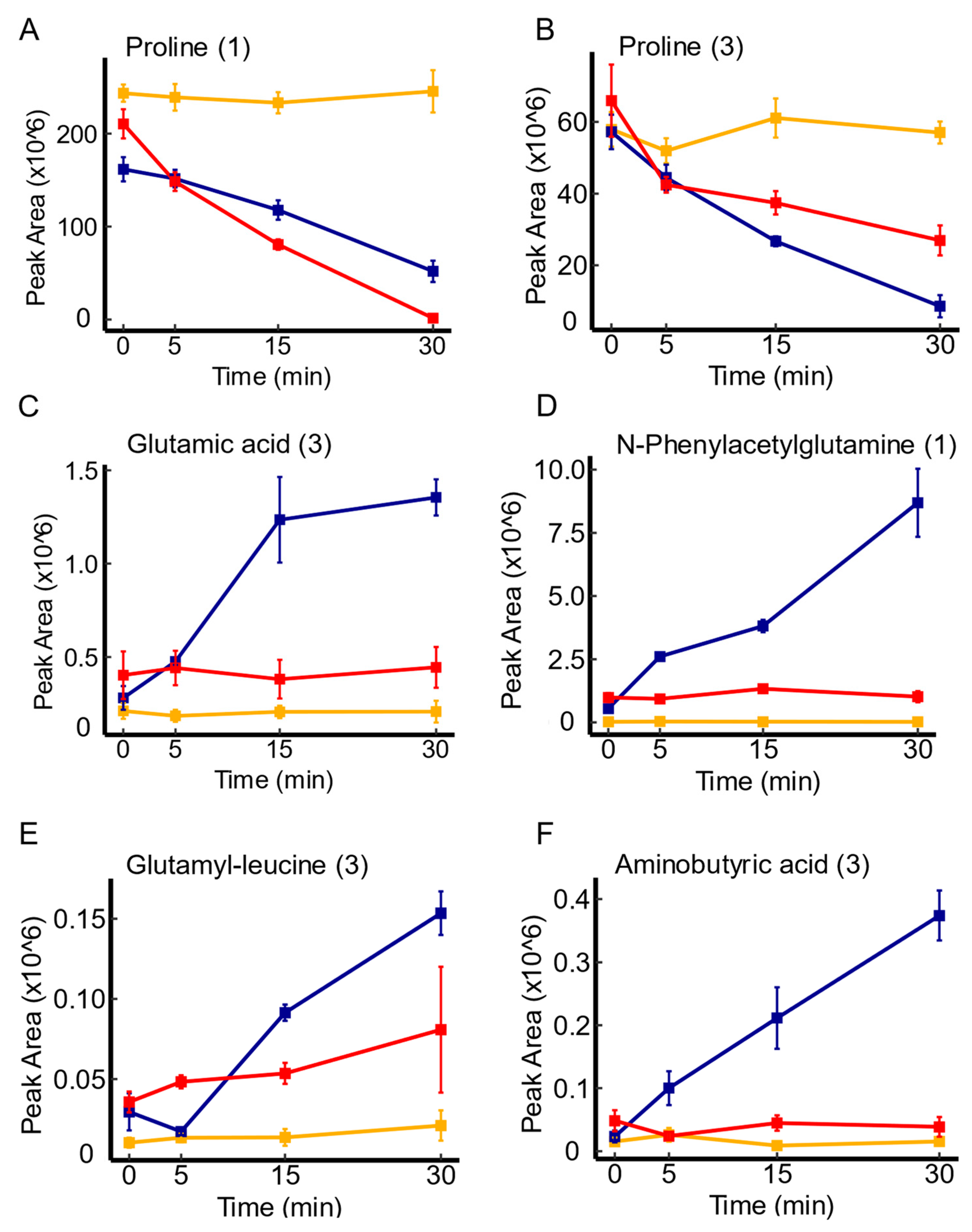
3.4. Candidate Substrates of yeaV
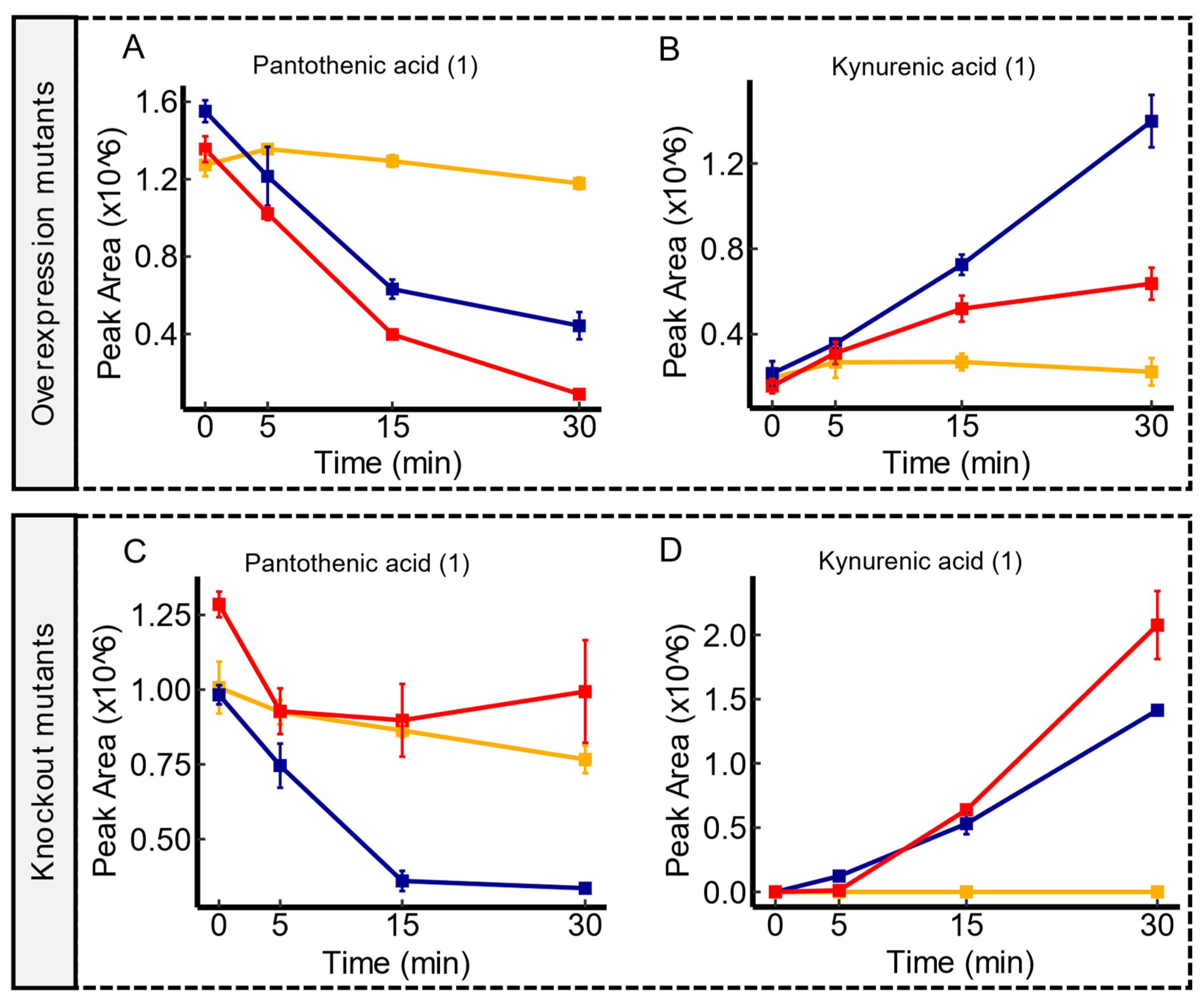
3.5. Candidate Substrates of hsrA
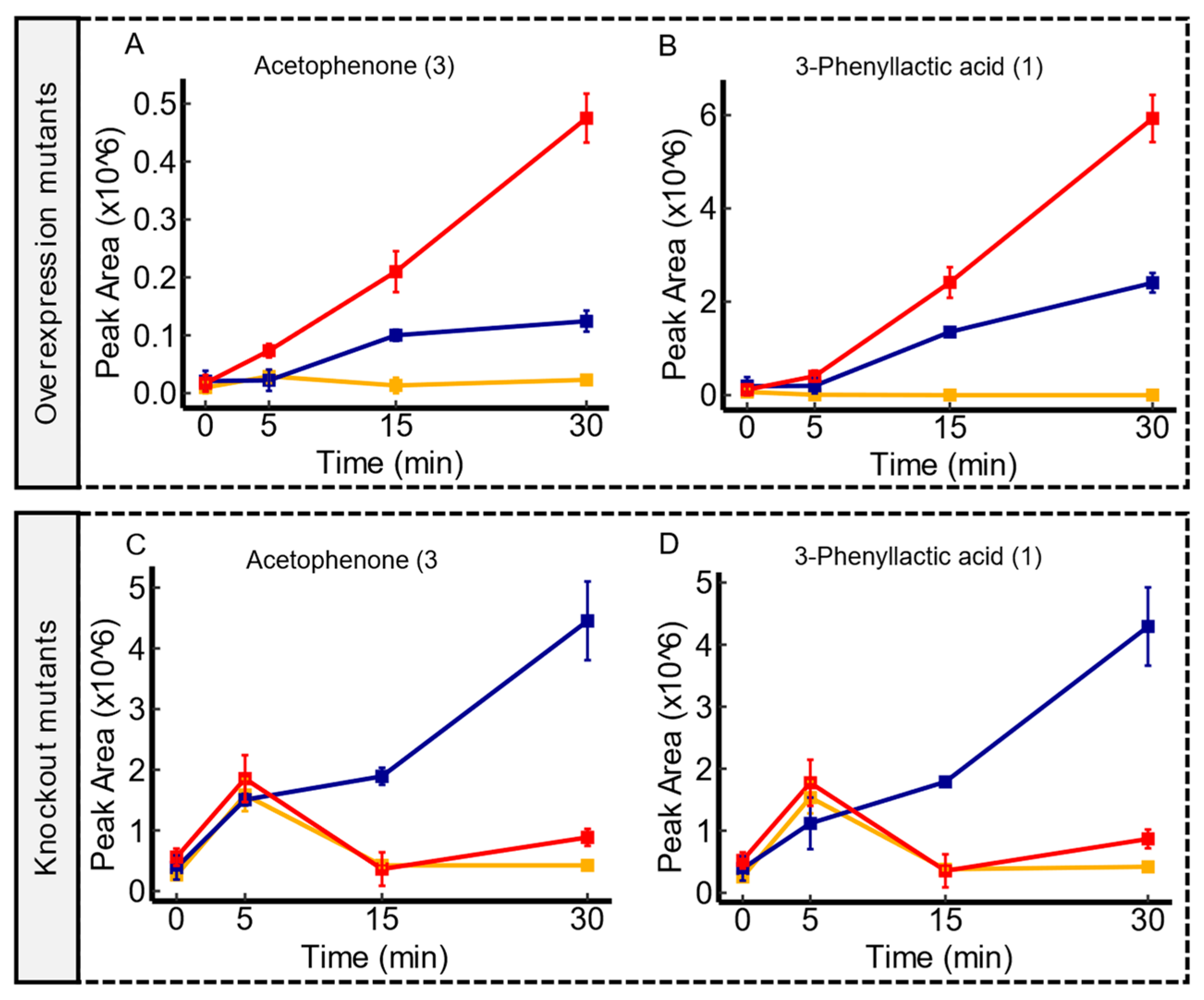
3.6. Candidate Substrates of ydjE
3.7. Candidate Substrates of yddA
3.8. Remaining Transporters
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thakur, V.; Uniyal, A.; Tiwari, V. A Comprehensive Review on Pharmacology of Efflux Pumps and Their Inhibitors in Antibiotic Resistance. Eur. J. Pharmacol. 2021, 903, 174151. [Google Scholar] [CrossRef]
- Nishino, K.; Yamasaki, S.; Nakashima, R.; Zwama, M.; Hayashi-Nishino, M. Function and Inhibitory Mechanisms of Multidrug Efflux Pumps. Front. Microbiol. 2021, 12, 737288. [Google Scholar] [CrossRef]
- Alam, S.; Doherty, E.; Ortega-Prieto, P.; Arizanova, J.; Fets, L. Membrane Transporters in Cell Physiology, Cancer Metabolism and Drug Response. Dis. Model. Mech. 2023, 16, dmm050404. [Google Scholar] [CrossRef]
- Giacomini, K.M.; Huang, S.M.; Tweedie, D.J.; Benet, L.Z.; Brouwer, K.L.R.; Chu, X.; Dahlin, A.; Evers, R.; Fischer, V.; Hillgren, K.M.; et al. Membrane Transporters in Drug Development. Nat. Rev. Drug Discov. 2010, 9, 215–236. [Google Scholar] [CrossRef]
- Kell, D.B.; Swainston, N.; Pir, P.; Oliver, S.G. Membrane Transporter Engineering in Industrial Biotechnology and Whole Cell Biocatalysis. Trends Biotechnol. 2015, 33, 237–246. [Google Scholar] [CrossRef]
- Jezierska, S.; Van Bogaert, I.N.A. Crossing Boundaries: The Importance of Cellular Membranes in Industrial Biotechnology. J. Ind. Microbiol. Biotechnol. 2017, 44, 721–733. [Google Scholar] [CrossRef]
- Borodina, I. Understanding Metabolite Transport Gives an Upper Hand in Strain Development. Microb. Biotechnol. 2019, 12, 69–70. [Google Scholar] [CrossRef]
- Salvador López, J.M.; Van Bogaert, I.N.A. Microbial Fatty Acid Transport Proteins and Their Biotechnological Potential. Biotechnol. Bioeng. 2021, 118, 2184–2201. [Google Scholar] [CrossRef]
- Mukhopadhyay, A. Tolerance Engineering in Bacteria for the Production of Advanced Biofuels and Chemicals. Trends Microbiol. 2015, 23, 498–508. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhou, C.; Wang, Y.; Li, C. Transporter Engineering for Microbial Manufacturing. Biotechnol. J. 2020, 15, 1900494. [Google Scholar] [CrossRef]
- Kell, D.B. Control of Metabolite Efflux in Microbial Cell Factories: Current Advances and Future Prospects. In Fermentation Microbiology and Biotechnology, 4th ed.; CRC Press: Boca Raton, FL, USA, 2018; pp. 117–138. [Google Scholar] [CrossRef]
- Kell, D.B. The Transporter-Mediated Cellular Uptake and Efflux of Pharmaceutical Drugs and Biotechnology Products: How and Why Phospholipid Bilayer Transport Is Negligible in Real Biomembranes. Molecules 2021, 26, 5629. [Google Scholar] [CrossRef]
- Khunnonkwao, P.; Jantama, S.S.; Kanchanatawee, S.; Jantama, K. Re-Engineering Escherichia Coli KJ122 to Enhance the Utilization of Xylose and Xylose/Glucose Mixture for Efficient Succinate Production in Mineral Salt Medium. Appl. Microbiol. Biotechnol. 2018, 102, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Malla, S.; Özdemir, E.; Kim, S.H.; Lennen, R.; Christensen, H.B.; Christensen, U.; Munro, L.J.; Herrgård, M.J.; Kell, D.B.; et al. Identification and Engineering of Transporters for Efficient Melatonin Production in Escherichia Coli. Front. Microbiol. 2022, 13, 880847. [Google Scholar] [CrossRef] [PubMed]
- Carreón-Rodríguez, O.E.; Gosset, G.; Escalante, A.; Bolívar, F. Glucose Transport in Escherichia Coli: From Basics to Transport Engineering. Microorganisms 2023, 11, 1588. [Google Scholar] [CrossRef] [PubMed]
- Gosset, G. Improvement of Escherichia Coli Production Strains by Modification of the Phosphoenolpyruvate: Sugar Phosphotransferase System. Microb. Cell Fact. 2005, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Blattner, F.R.; Plunkett, G.; Bloch, C.A.; Perna, N.T.; Burland, V.; Riley, M.; Collado-Vides, J.; Glasner, J.D.; Rode, C.K.; Mayhew, G.F.; et al. The Complete Genome Sequence of Escherichia Coli K-12. Science 1997, 277, 1453–1462. [Google Scholar] [CrossRef] [PubMed]
- Elbourne, L.D.H.; Tetu, S.G.; Hassan, K.A.; Paulsen, I.T. TransportDB 2.0: A Database for Exploring Membrane Transporters in Sequenced Genomes from All Domains of Life. Nucleic Acids Res. 2017, 45, D320. [Google Scholar] [CrossRef]
- Ghatak, S.; King, Z.A.; Sastry, A.; Palsson, B.O. The Y-Ome Defines the 35% of Escherichia Coli Genes That Lack Experimental Evidence of Function. Nucleic Acids Res. 2019, 47, 2446–2454. [Google Scholar] [CrossRef] [PubMed]
- Gründemann, D.; Harlfinger, S.; Golz, S.; Geerts, A.; Lazar, A.; Berkels, R.; Jung, N.; Rubbert, A.; Schömig, E. Discovery of the Ergothioneine Transporter. Proc. Natl. Acad. Sci. USA 2005, 102, 5256–5261. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Xu, J.Z.; Zhang, W.G. Advances and Prospects in Metabolic Engineering of Escherichia Coli for L-Tryptophan Production. World J. Microbiol. Biotechnol. 2022, 38, 22. [Google Scholar] [CrossRef]
- Andrews, A.E.; Lawley, B.; Pittard, A.J. Mutational Analysis of Repression and Activation of the TyrP Gene in Escherichia Coli. J. Bacteriol. 1991, 173, 5068–5078. [Google Scholar] [CrossRef]
- Brown, K.D. Maintenance and Exchange of the Aromatic Amino Acid Pool in Escherichia Coli. J. Bacteriol. 1971, 106, 70–81. [Google Scholar] [CrossRef]
- Hiraga, S.; Ito, K.; Matsuyama, T.; Ozaki, H.; Yura, T. 5-Methyltryptophan-Resistant Mutations Lniked with the Arginine G Marker in Escherichia Coli. J. Bacteriol. 1968, 96, 1880–1881. [Google Scholar] [CrossRef] [PubMed]
- Young, G.B.; Jack, D.L.; Smith, D.W.; Saier, M.H. The Amino Acid/Auxin: Proton Symport Permease Family. Biochim. Biophys. Acta 1999, 1415, 306–322. [Google Scholar] [CrossRef] [PubMed]
- Sarsero, J.P.; Wookey, P.J.; Gollnick, P.; Yanofsky, C.; Pittard, A.J. A New Family of Integral Membrane Proteins Involved in Transport of Aromatic Amino Acids in Escherichia Coli. J. Bacteriol. 1991, 173, 3231–3234. [Google Scholar] [CrossRef]
- Featherstone, D.E.; Broadie, K. Wrestling with Pleiotropy: Genomic and Topological Analysis of the Yeast Gene Expression Network. Bioessays 2002, 24, 267–274. [Google Scholar] [CrossRef]
- Jindal, S.; Yang, L.; Day, P.J.; Kell, D.B. Involvement of Multiple Influx and Efflux Transporters in the Accumulation of Cationic Fluorescent Dyes by Escherichia Coli. BMC Microbiol. 2019, 19, 195. [Google Scholar] [CrossRef] [PubMed]
- Nishino, K.; Yamaguchi, A. Analysis of a Complete Library of Putative Drug Transporter Genes in Escherichia Coli. J. Bacteriol. 2001, 183, 5803–5812. [Google Scholar] [CrossRef] [PubMed]
- Mirandela, G.D.; Tamburrino, G.; Hoskisson, P.A.; Zachariae, U.; Javelle, A. The Lipid Environment Determines the Activity of the Escherichia Coli Ammonium Transporter AmtB. FASEB J. 2019, 33, 1989–1999. [Google Scholar] [CrossRef]
- Malla, S.; van der Helm, E.; Darbani, B.; Wieschalka, S.; Förster, J.; Borodina, I.; Sommer, M.O.A. A Novel Efficient L-Lysine Exporter Identified by Functional Metagenomics. Front. Microbiol. 2022, 13, 855736. [Google Scholar] [CrossRef]
- Sá-Correia, I.; Godinho, C.P. Exploring the Biological Function of Efflux Pumps for the Development of Superior Industrial Yeasts. Curr. Opin. Biotechnol. 2022, 74, 32–41. [Google Scholar] [CrossRef]
- Erian, A.M.; Egermeier, M.; Marx, H.; Sauer, M. Insights into the Glycerol Transport of Yarrowia Lipolytica. Yeast 2022, 39, 323–336. [Google Scholar] [CrossRef]
- Zelena, E.; Dunn, W.B.; Broadhurst, D.; Francis-McIntyre, S.; Carroll, K.M.; Begley, P.; O’Hagan, S.; Knowles, J.D.; Halsall, A.; Wilson, I.D.; et al. Development of a Robust and Repeatable UPLC-MS Method for the Long-Term Metabolomic Study of Human Serum. Anal. Chem. 2009, 81, 1357–1364. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-Mcintyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for Large-Scale Metabolic Profiling of Serum and Plasma Using Gas Chromatography and Liquid Chromatography Coupled to Mass Spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, S.; Kell, D.B. Consensus Rank Orderings of Molecular Fingerprints Illustrate the Most Genuine Similarities between Marketed Drugs and Small Endogenous Human Metabolites, but Highlight Exogenous Natural Products as the Most Important ‘Natural’ Drug Transporter Substrates. ADMET DMPK 2017, 5, 85–125. [Google Scholar] [CrossRef]
- Wright Muelas, M.; Roberts, I.; Mughal, F.; O’Hagan, S.; Day, P.J.; Kell, D.B. An Untargeted Metabolomics Strategy to Measure Differences in Metabolite Uptake and Excretion by Mammalian Cell Lines. Metabolomics 2020, 16, 107. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Venkat, S.; Wilson, J.; McGuire, P.; Chang, A.L.; Gan, Q.; Fan, C. Genome-Wide Quantification of the Effect of Gene Overexpression on Escherichia Coli Growth. Genes 2018, 9, 414. [Google Scholar] [CrossRef]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia Coli K-12 in-Frame, Single-Gene Knockout Mutants: The Keio Collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef]
- Kell, D.B.; Brown, M.; Davey, H.M.; Dunn, W.B.; Spasic, I.; Oliver, S.G. Metabolic Footprinting and Systems Biology: The Medium Is the Message. Nat. Rev. Microbiol. 2005, 3, 557–565. [Google Scholar] [CrossRef]
- Allen, J.; Davey, H.M.; Broadhurst, D.; Heald, J.K.; Rowland, J.J.; Oliver, S.G.; Kell, D.B. High-Throughput Classification of Yeast Mutants for Functional Genomics Using Metabolic Footprinting. Nat. Biotechnol. 2003, 21, 692–696. [Google Scholar] [CrossRef]
- Saier, M.H.; Reddy, V.S.; Tsu, B.V.; Ahmed, M.S.; Li, C.; Moreno-Hagelsieb, G. The Transporter Classification Database (TCDB): Recent Advances. Nucleic Acids Res. 2016, 44, D372–D379. [Google Scholar] [CrossRef]
- Schymanski, E.L.; Jeon, J.; Gulde, R.; Fenner, K.; Ruff, M.; Singer, H.P.; Hollender, J. Identifying Small Molecules via High Resolution Mass Spectrometry: Communicating Confidence. Environ. Sci. Technol. 2014, 48, 2097–2098. [Google Scholar] [CrossRef] [PubMed]
- Conway, J.R.; Lex, A.; Gehlenborg, N. UpSetR: An R Package for the Visualization of Intersecting Sets and Their Properties. Bioinformatics 2017, 33, 2938–2940. [Google Scholar] [CrossRef] [PubMed]
- Lewinson, O.; Adler, J.; Sigal, N.; Bibi, E. Promiscuity in Multidrug Recognition and Transport: The Bacterial MFS Mdr Transporters. Mol. Microbiol. 2006, 61, 277–284. [Google Scholar] [CrossRef]
- Radi, M.S.; Munro, L.J.; Salcedo-Sora, J.E.; Kim, S.H.; Feist, A.M.; Kell, D.B. Understanding Functional Redundancy and Promiscuity of Multidrug Transporters in E. Coli under Lipophilic Cation Stress. Membranes 2022, 12, 1264. [Google Scholar] [CrossRef] [PubMed]
- McHugh, J.P.; Rodríguez-Quiñones, F.; Abdul-Tehrani, H.; Svistunenko, D.A.; Poole, R.K.; Cooper, C.E.; Andrews, S.C. Global Iron-Dependent Gene Regulation in Escherichia Coli. A New Mechanism for Iron Homeostasis. J. Biol. Chem. 2003, 278, 29478–29486. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, N.; Nishino, K.; Yamaguchi, A. Novel Macrolide-Specific ABC-Type Efflux Transporter in Escherichia Coli. J. Bacteriol. 2001, 183, 5639–5644. [Google Scholar] [CrossRef]
- Smith, H.E.; Blair, J.M.A. Redundancy in the Periplasmic Adaptor Proteins AcrA and AcrE Provides Resilience and an Ability to Export Substrates of Multidrug Efflux. J. Antimicrob. Chemother. 2014, 69, 982–987. [Google Scholar] [CrossRef]
- Wong, K.; Ma, J.; Rothnie, A.; Biggin, P.C.; Kerr, I.D. Towards Understanding Promiscuity in Multidrug Efflux Pumps. Trends Biochem. Sci. 2014, 39, 8–16. [Google Scholar] [CrossRef]
- Radi, M.S.; SalcedoSora, J.E.; Kim, S.H.; Sudarsan, S.; Sastry, A.V.; Kell, D.B.; Herrgård, M.J.; Feist, A.M. Membrane Transporter Identification and Modulation via Adaptive Laboratory Evolution. Metab. Eng. 2022, 72, 376–390. [Google Scholar] [CrossRef]
- Stanchev, L.D.; Møller-Hansen, I.; Lojko, P.; Rocha, C.; Borodina, I. Screening of Saccharomyces Cerevisiae Metabolite Transporters by 13C Isotope Substrate Labeling. Front. Microbiol. 2023, 14, 1286597. [Google Scholar] [CrossRef] [PubMed]
- Darbani, B.; Stovicek, V.; van der Hoek, S.A.; Borodina, I.; Darbani, B.; Stovicek, V.; van der Hoek, S.A.; Borodina, I. Engineering Energetically Efficient Transport of Dicarboxylic Acids in Yeast Saccharomyces Cerevisiae. Proc. Natl. Acad. Sci. USA 2019, 116, 19415–19420. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, A.D.; Swainston, N.; Samanta, S.; Roberts, I.; Muelas, M.W.; Kell, D.B. MassGenie: A Transformer-Based Deep Learning Method for Identifying Small Molecules from Their Mass Spectra. Biomolecules 2021, 11, 1793. [Google Scholar] [CrossRef] [PubMed]
- Sastry, A.V.; Gao, Y.; Szubin, R.; Hefner, Y.; Xu, S.; Kim, D.; Choudhary, K.S.; Yang, L.; King, Z.A.; Palsson, B.O. The Escherichia Coli Transcriptome Mostly Consists of Independently Regulated Modules. Nat. Commun. 2019, 10, 5536. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transporter/TC# a | Family | Previously Identified Substrates |
|---|---|---|
| yddG/2.A.7.17.2 | Aromatic Amino Acid/Paraquat Exporter (ArAA/P-E) | Aromatic amino acids |
| Mtr/2.A.42.1.2 | The Hydroxy/Aromatic Amino Acid Permease (HAAAP) | Tryptophan |
| ybbW/2.A.39.3.8 | Nucleobase:Cation Symporter-1 (NCS1) | Allantoin |
| ynfM/2.A.1.36.1 | Acriflavin-sensitivity (YnfM) Family | Arabinose |
| uraA/2.A.40.1.1 | Nucleobase/Ascorbate Transporter (NAT) or Nucleobase:Cation Symporter-2 (NCS2) | Uracil |
| sapF/3.A.1.5.42 | Peptide/Opine/Nickel Uptake Transporter (PepT) | Putrescine |
| yeeE(tsuA)/9.B.102.1.2 | The YedE/YeeE (YeeE) | Thiosulfate |
| yifK/2.A.3.1.15 | Amino Acid Transporter (AAT) | Serine and threonine |
| mdtJ/2.A.7.1.9 | Small multidrug resistance (SMR) | Spermidine, SDS b, deoxycholate |
| yojI/3.A.1.113.3 | The Peptide-3 Exporter (Pep3E) | Microcin |
| ydfJ/2.A.1.6.9 | Metabolite:H+ Symporter (MHS) | K+ * |
| ydhK/2.A.85.1.6 | The Aromatic Acid Exporter (ArAE) | Uncharacterized |
| yeaV/2.A.15.2.3 | The Betaine/Carnitine/Choline Transporter (BCCT) Family | Uncharacterized |
| yfdV/2.A.69.3.5 | Auxin Efflux Carrier (AEC) | Uncharacterized |
| yfdC/1.A.16.4.1 | Formate-Nitrite Transporter (FNT) | Uncharacterized |
| rarD/2.A.7.7.2 | Chloramphenicol-Sensitivity Protein | Uncharacterized |
| hsrA/2.A.1.3.51 | Drug:H+ Antiporter-2 (14 Spanner) (DHA2) | Uncharacterized |
| yfcJ/2.A.1.46.6 | Uncharacterized Major Facilitator-5 (UMF5) | Uncharacterized |
| ydjX/9.B.27.1.1 | Death Effector Domain A (DedA) | Uncharacterized |
| dcuD/2.A.61.1.2 | The C4-dicarboxylate Uptake C (DcuC) Family | Uncharacterized |
| mdlA/3.A.1.106.13 | Lipid Exporter (LipidE) Family | Uncharacterized |
| ydjE/2.A.1.1.92 | Sugar Porter (SP) | Uncharacterized |
| yddA/3.A.1.203.11 | The Peroxysomal Fatty Acyl CoA Transporter (P-FAT) | Uncharacterized |
| ygiS/3.A.1.5.41 | The ATP-binding cassette superfamily | Uncharacterized |
| yiaV/8.A.1.1.4 | The Membrane Fusion Protein (MFP) Family | Uncharacterized |
| yidE/2.A.81.1.5 | Aspartate:Alanine Exchanger (AAEx) Family | Uncharacterized |
| ESI+ | % All Compounds | ESI− | % All Compounds | |
|---|---|---|---|---|
| Features Compounds Unique Compounds | 531,376 | 135,776 | ||
| 9406 | 4092 | |||
| 3671 | 39% | 1632 | 40% | |
| MS2 No MS2 | 5607 | 60% | 3287 | 80% |
| 3779 | 40% | 805 | 20% | |
| MS2 Preferred Ion | 5305 | 56% | 3213 | 79% |
| MS2 Other Ion | 302 | 3% | 74 | 2% |
| Level 1 (Match to reference standard) * | 16 | 0.2% | 67 | 2% |
| Level 2 (Probable structure, mzCloud > 70%) * Level 3 (Tentative Candidate, match to predicted composition) * | 461 | 5% | 70 | 2% |
| 7062 | 75% | 2520 | 62% | |
| Level 4 (Molecular formula) * | 504 | 5% | 251 | 6% |
| Level 5 (Mass) * | 1363 | 14% | 1184 | 29% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radi, M.S.; Munro, L.J.; Rago, D.; Kell, D.B. An Untargeted Metabolomics Strategy to Identify Substrates of Known and Orphan E. coli Transporters. Membranes 2024, 14, 70. https://doi.org/10.3390/membranes14030070
Radi MS, Munro LJ, Rago D, Kell DB. An Untargeted Metabolomics Strategy to Identify Substrates of Known and Orphan E. coli Transporters. Membranes. 2024; 14(3):70. https://doi.org/10.3390/membranes14030070
Chicago/Turabian StyleRadi, Mohammad S., Lachlan J. Munro, Daniela Rago, and Douglas B. Kell. 2024. "An Untargeted Metabolomics Strategy to Identify Substrates of Known and Orphan E. coli Transporters" Membranes 14, no. 3: 70. https://doi.org/10.3390/membranes14030070
APA StyleRadi, M. S., Munro, L. J., Rago, D., & Kell, D. B. (2024). An Untargeted Metabolomics Strategy to Identify Substrates of Known and Orphan E. coli Transporters. Membranes, 14(3), 70. https://doi.org/10.3390/membranes14030070